
Study on monitoring technology of fish motion trajectories based on Acoustic Tag System
-
摘要:
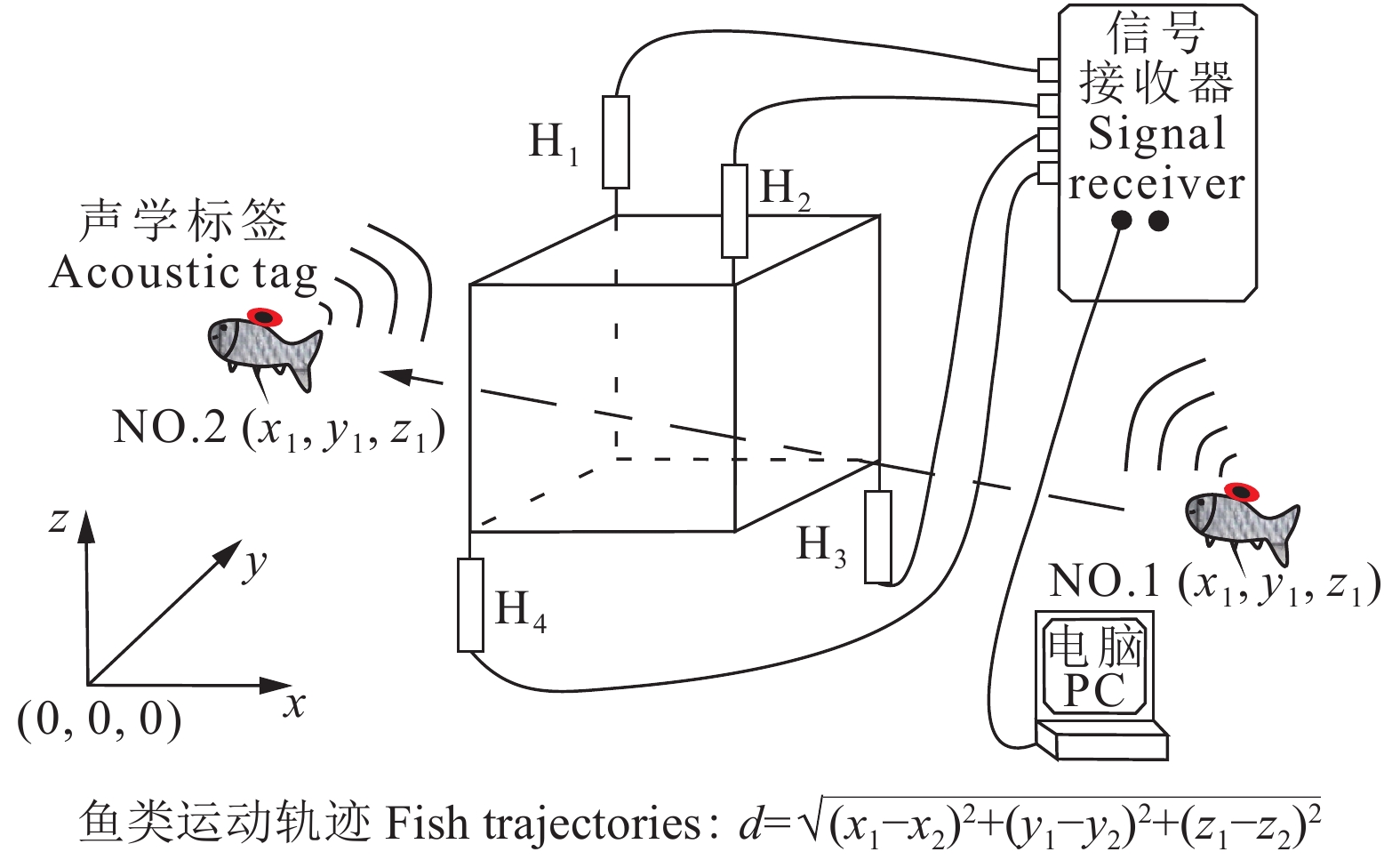
声学标签监测技术是声波监测技术的一种主动识别方法,其通过接收、处理移植或捆绑于鱼类身上的声学标签发射的声波信号来获得鱼类二维、三维运动轨迹,具有原位观察、定位准确、数据处理简单和数据连续性好等优点。文章介绍了声学标签监测技术的组成及工作原理,并根据异常数据的不同特点总结了一套完整的数据去噪、清洗的处理方法,该法可快速得到鱼类的二维、三维运动轨迹。通过实例分析了鱼类运动轨迹的分布情况以及变化规律。根据鱼类在不同水生态环境中的运动行为响应情况,可以直接或间接判断水中污染物或有毒物质对鱼类的潜在影响以及实际毒性,为渔业养殖水质环境评价、水质监测及预警、水生态健康评价以及水生态修复效果评价的研究提供参考依据。
Abstract:Acoustic tag monitoring technology, an active identification method of acoustic detection technology, can obtain 2D and 3D motion trajectories of fish by receiving and processing the acoustic signals emitting from acoustic tags which are transplanted or tied to fish. It has the advantages of in-situ observation, accurate positioning, simple data processing and good data continuity. The paper introduces the composition and working principle of acoustic tag monitoring technology. A complete data processing method of denoising and cleaning is proposed according to the different characteristics of abnormal data. With this method, the real-time 2D and 3D motion trajectories can be quickly obtained. The distribution of fish motion trajectories and changing rules are analyzed by an example. According to the behavioral response of fish in different water ecological environments, the potential effects and actual toxicity of pollutants or toxic substances in water can be judged directly or indirectly. It provides the references for water quality assessment in aquaculture, monitoring and early warning of water environment, health evaluation of aquatic ecological, as well as evaluation of aquatic ecological restoration.
-
长体圆鲹(Decapterus macrosoma),又名长身圆鲹,隶属于鲈形目、鲹科、圆鲹属,主要分布于中国南海、印度尼西亚、澳洲和日本南部沿海等地[1]。长体圆鲹在中国南海分布较广,是南海灯光围网渔业的主要捕捞对象,具有较高的经济价值[2-5]。目前,国内外学者关于长体圆鲹的研究主要集中在生长繁殖[6]和资源评估[7-8]方面,与种群遗传和分子标记相关的研究报道较少。微卫星标记仅见翟云等[9]开发蓝圆鲹微卫星标记中获得5个跨物种通用标记可于用长体圆鲹,并无专门针对长体圆鲹开发的微卫星标记。种群遗传信息的匮乏,将大大影响对其资源的评估和长期有效的管理。
微卫星分子标记因是共显性标记,具有多态性高、变异性强、数据易统计等突出优点[10],广泛应用于海洋生物遗传结构及遗传多样性分析[11-12]。但由于微卫星标记通用性较差,常常具有极强的种属特异性。鱼类微卫星标记开发中多以二核苷酸重复为主[13-15],普遍认为它们具有较高的遗传变异[16],但是也有部分学者研究认为三、四核苷酸重复位点较二核苷酸重复具有更高的筛选效率和多态性[17-19]。
本研究通过RAD-Seq高通量测序方法开发长体圆鲹二、三核苷酸微卫星分子标记,并对测试群体进行多样性分析,同时比较二、三核苷酸的筛选效率和多态性差异,旨在为长体圆鲹种群遗传结构及遗传多样性分析提供技术基础,并为该资源的评估和管理提供帮助。
1. 材料与方法
1.1 样品采集与基因组DNA提取
长体圆鲹样品采集于中国南海中沙群岛东部海域,共35尾。剪取部分肌肉样品加入无水乙醇保存。每个样品剪取少量肌肉组织,使用海洋动物组织基因组DNA提取试剂盒(天根,北京) 提取基因组DNA,0.8%的琼脂糖凝胶电泳检测DNA提取质量,−20 ℃保存备用。
1.2 高通量测序与引物合成
使用HiSeq2000高通量测序仪(Illumina,USA) 对长体圆鲹基因组DNA进行RAD-seq (测序服务由广州基迪奥生物科技有限公司提供),经生物信息学搜索出微卫星位点[20]。使用Premier 5.0软件在重复单元侧翼序列上选择性设计出112条引物,主要参数为:G-C含量为40%~60%,引物长度为18~25 bp,退火温度为45~60 ℃,预期产物长度180~320 bp。送上海英潍捷基贸易有限公司合成引物。
1.3 引物筛选与分型检测
选取3个样本混合成的基因组DNA为模板,优化PCR反应条件,对引物进行首轮筛选,琼脂糖电泳检测是否能扩增出稳定且均一的目的片段。之后选取8尾个体的基因组DNA作为模板,使用三引物法[21],利用M13荧光接头引物进行PCR扩增,扩增产物送华大基因公司经毛细管电泳进行等位基因分型,检测引物是否具有多态性。PCR反应体系为15 μL,其中包括10×PCR Buffer 1.5 μL,2.5 mmol·L–1 MgCL2 1.2 μL,2 mmol·L–1 dNTPs 2 μL,M13正向引物(10 μmol·L–1) 0.2 μL,M13反向引物(10 μmol·L–1) 0.6 μL,M13通用荧光引物(10 μmol·L–1) 0.5 μL,Taq酶(5 U·μL–1) 0.15 μL,DNA模版1 μL,加双蒸水至15 μL。PCR扩增程序为:94 ℃预变性5 min;94 ℃变性45 s,55~60 ℃退火45 s,72 ℃延伸45 s,35个循环;94 ℃变性45 s,53 ℃退火45 s,72 ℃延伸45 s,8个循环;72 ℃延伸30 min。
1.4 长体圆鲹群体遗传学评价
使用35尾长体圆鲹个体的基因组DNA为模板,对通过筛选的微卫星标记的种群遗传学特征进行评价。PCR反应体系和条件、等位基因分型方法如上。使用软件Genepop 4.0[22]对每个标记的种群遗传学特征值进行计算,包括等位基因数(Na)、表观杂合度(Ho)和期望杂合度(He),进行“哈迪-温伯格”平衡(HWE)检验和连锁不平衡检测,并对P值进行Bonferroni校正。使用Cervus 3.0.7[23]软件计算多态信息含量(PIC)。
2. 结果
2.1 高通量测序结果与微卫星位点分析
RAD-seq高通量测序共获得长体圆鲹基因组原始数据2.06 G,GC含量为41.43%,Q30达93.05%。说明测序结果质量较好,可用于后续分析。搜索后共获得微卫星序列58 180条,一至六核苷酸重复微卫星位点70 508个,其中二核苷酸重复微卫星位点最多(37 646个),占总数的53.39% (表1),说明二核苷酸重复为主要的微卫星类型。二核苷酸重复微卫星位点共有4种重复类型,4种类型重复微卫星数量相差较大,AC/GT类有29 754个,占二核苷酸重复的68.4%;AG/CT类有6 487个,占17.2%;AT/TA有1 340个,占3.6%;GC/CG仅有65个,占0.17%。
表 1 长体圆鲹基因组中不同类型SSR统计Table 1. Different types of SSR statistics in D. macrosoma genome重复单元
repeat unit微卫星数量/个
number of microsatellite占比/%
ratio一核苷酸 mono-nucleotide 8 184 11.61 二核苷酸 di-nucleotide 37 646 53.39 三核苷酸 tri-nucleotide 13 960 19.80 四核苷酸 tetra-nucleotide 7 741 10.98 五核苷酸 penta-nucleotide 2 255 3.20 六核苷酸 hexa-nucleotide 722 1.02 合计 total 70 508 100.00 2.2 PCR引物设计和筛选
选取112条二、三核苷酸重复序列设计引物,其中二核苷酸重复为81对,三核苷酸重复为31对。经过筛选后,共有27对引物通过筛选(表2),27对引物扩增的序列中18个位点为二核苷酸重复,重复次数为9~14次;8个位点为三核苷酸重复,重复次数为6~10次。二核苷酸重复位点检出效率为22.2%,三核苷酸重复位点检出效率为25.8%。
表 2 27对长体圆鲹微卫星引物信息Table 2. Information of 27 pairs of primers in D. macrosoma位点
locus引物序列 (5'−3')
primer sequence重复单元
repeat motif退火温度/℃
annealing temperature期望长度/bp
allele sizeDma03 F:CCACGCCTATTGAGTTACAGA (CA)9 60 186 R:GAGCCAGTGGATGAACAGAGT Dma07 F:GCCCCTGTGGGTGTGTGA (CA)9 60 225 R:GGGTGGTGGGTTCGGTTT Dma12 F:GAACCAGTGCCTACAATAGA (AC)9 60 243 R:CTGCTCACGGTAAGTCCA Dma15 F:ACAGGAAGGAACAGGACAG (TG)10 55 254 R:TATTGAAGTGAAAAAGCCG Dma22 F:CGCTGTTGAAATGAAGAAGA (GT)10 60 317 R:AGTGATGTCGCCTCATAAAT Dma23 F:AAACTGAGGGCGAGATAGAGG (AC)10 55 190 R:CCACAGGCTGAGTAAACCAAC Dma26 F:ATCCCATTCACCGACATAG (TG)10 58 258 R:CTGTGGTATCGTTCCCTGT Dma28 F:TGATTGGCTTCTACTCTGC (AC)10 55 281 R:AGTGGCTTGTTTGACTCTTAT Dma36 F:GGATGTAGTGAAGAGGGGAG (GT)11 55 239 R:CACAATCAGTGTTATGGCAG Dma38 F:GCCAATAAAGGCAAACAGT (CA)11 60 227 R:ATCCGAGACAAAGACATACAA Dma39 F:AGTGTGCTGACTTTTCTCTG (CA)11 55 241 R:TTATTGTTTGTTGTCTGGGT Dma45 F:CTCCTTTTTCTTCTTCCTCT (CA)11 60 281 R:CTACCTGCTCTTCAACTCAT Dma51 F:TGACAGCCTCCACTACTCC (GA)12 55 225 R:GCTAACCAGACACGCAAA Dma54 F:AAAGCCCATCTGTCTCGT (GT)12 60 202 R:TGTTTCAGTCCGTTCCTG Dma58 F:TCAAGAGGGAGTGGGAGC (AC)12 58 279 R:TCAAATGGGTGTTTAGCG Dma64 F:GCTCAGACTGCGTGGACA (TG)13 55 314 R:GCTGGTGAACAACAGGACA Dma72 F:TTCCGCAGGCATAAAAAC (CT)13 58 301 R:CCAAGGTCCGCTACACTA Dma76 F:TTCTCGCTGACCTGCTTG (TG)14 55 253 R:GCGTCCTCGTCGTCTTTC Dma81 F:GAGACACGGTCAGAAAACA (TGC)6 60 216 R:GGAAGTAGGACTCTAGGGG Dma82 F:CTGTCACTCCATTCCTATTCC (GTT)6 58 244 R:CCTACATTTGTGCTTTTGTTC Dma83 F:CTCTAAAGCCGACCTAACC (CTT)6 58 239 R:TGTCTCAACACAGCGAAAC Dma84 F:AAACTAACTCATCACCAG (TGT)6 55 283 R:AAACGACAGGAACTCAAT Dma85 F:CTCACTTTGACCCAACCAG (AGG)6 55 256 R:CCTTTCACCGAGACACCAG Dma131 F:TGCGGATGGGTGGTAGTGT (GGT)8 55 208 R:ATTGCTGGTAGTCGGTGGC Dma132 F:CCCAGTGAGACCAGAACCA (GCT)8 55 268 R:GACCCGTAGACAGGAGAGT Dma135 F:GTTGTTGTTTTTTTCCTT (GCA)9 55 301 R:CATCAGTCTGGCTTTATA Dma145 F:ACGATACAGCAGCCGAAG (TCA)10 60 197 R:AGTGATGTCGCCTCATAAAT 2.3 微卫星标记的种群遗传学评价
使用1个采集自南海东南部海域的长体圆鲹群体对筛选合格的微卫星标记进行种群遗传学评价。所有27个标记在测试群体中共检测到285个等位基因,等位基因数为5~17,Ho为0.342 9~0.857 1,平均为0.631 7;He为0.538 3~0.911 8,平均为0.7968。PIC为0.497~0.886,平均为0.780 9 (表3),表明开发的微卫星位点具有较高的多态性。共有19个标记等位基因频率符合“哈迪-温伯格”平衡。连锁不平衡检测表明各位点间无连锁不平衡现象。
表 3 长体圆鲹微卫星标记的种群遗传学特征Table 3. Characteristics of microsatellite loci in D. macrosoma位点
locusN Na Ho He PHWE PIC Dma03 35 8 0.857 1 0.790 9 0.042 0 0.746 0 Dma07 35 9 0.771 4 0.855 9 0.553 4 0.825 0 Dma12 34 11 0.685 7 0.816 6 0.106 4 0.809 0 Dma15* 35 14 0.542 9 0.911 8 0.000 0 0.890 0 Dma22* 32 14 0.485 7 0.813 2 0.000 0 0.864 0 Dma23 34 17 0.714 3 0.864 8 0.003 3 0.868 0 Dma26* 34 14 0.428 6 0.869 1 0.000 0 0.870 0 Dma28 35 11 0.742 9 0.837 7 0.035 3 0.803 0 Dma36 35 9 0.685 7 0.786 3 0.054 5 0.743 0 Dma38* 34 12 0.628 6 0.851 2 0.000 0 0.849 0 Dma39* 33 12 0.514 3 0.782 0 0.000 0 0.793 0 Dma45 34 12 0.771 4 0.847 8 0.139 2 0.844 0 Dma51* 31 12 0.485 7 0.784 5 0.000 0 0.859 0 Dma54 35 15 0.771 4 0.900 6 0.139 0 0.878 0 Dma58 34 12 0.771 4 0.845 6 0.449 7 0.843 0 Dma64* 31 13 0.428 6 0.788 3 0.000 0 0.865 0 Dma72 35 8 0.628 6 0.713 9 0.155 7 0.659 0 Dma76 35 12 0.685 7 0.908 1 0.003 6 0.886 0 Dma81 35 8 0.628 6 0.795 0 0.045 7 0.752 0 Dma82 35 5 0.485 7 0.538 3 0.023 6 0.497 0 Dma83* 34 8 0.342 9 0.705 8 0.000 0 0.674 0 Dma84 35 7 0.542 9 0.704 8 0.005 9 0.649 0 Dma85 35 8 0.828 6 0.747 8 0.876 9 0.704 0 Dma131 35 9 0.628 6 0.717 6 0.646 6 0.661 0 Dma132 35 8 0.657 1 0.746 6 0.365 5 0.692 0 Dma135 34 10 0.742 9 0.840 5 0.241 8 0.835 0 Dma145 34 7 0.600 0 0.749 3 0.077 4 0.725 0 注:N. 有效样品数;Na. 等位基因数;Ho. 表观杂合度;He. 期望杂合度;PHWE. “哈迪-温伯格”平衡显著性检验P值;PIC. 多态信息含量;*. 经Bonferroni校正后显著背离“哈迪-温伯格”平衡 (校正P<0.001 85) Note: N. effective number of samples; Na. number of alleles; Ho. observed heterozygosity; He. expected heterozygosity; PHWE. Hardy–Weinberg probability test; PIC. polymorphism information content; *. significant deviation from HWE after Bonferroni's correction (adjusted P-value<0.001 85) 3. 讨论
3.1 高通量测序发掘微卫星序列的技术优势
传统微卫星标记开发方法耗时长、花费高、技术难度大。以磁珠富集法为例,标记开发过程中基因组DNA浓度、接头连接效率、富集过程中的杂交温度以及洗脱条件的控制等因素都会影响微卫星筛选的效率[24-25],且最终获得的有效微卫星序列仅几百条[26-27]。相比较而言,高通量测序技术开发微卫星标记,省略了建库、克隆、筛选等,只需提取基因组DNA测序,利用生物信息学手段可直接获取微卫星序列,通常是传统方法获得微卫星序列数目的几百倍[15,28-29],具有高效、便捷、准确的特点,能够满足短时间内大批量微卫星位点的开发需求,比如连锁图谱构建、QTL定位等[30-31]。
本次RAD-seq高通量测序共获得长体圆鲹基因组原始数据2.06 G,GC含量为41.43%,测序质量Q30达93.05%;共获得微卫星序列58 180条,一至六核苷酸重复微卫星位点70 508个。说明测序质量稳定高效,并获得了数量庞大、类型丰富的长体圆鲹微卫星序列,可用于后续长体圆鲹微卫星标记的大规模开发和相关遗传学研究。
3.2 不同核苷酸重复微卫星位点比较
本次高通量测序结果表明在长体圆鲹微卫星位点中二核苷酸重复为主要重复类型,AC/GT类重复数量最为丰富,GC/CG重复较为少见。此结果与大量水产动物微卫星位点研究结果相一致[32-34],差异仅在于比例多少,以及除二核苷酸重复占主要地位外其他核苷酸重复的含量差异。熊良伟等[33]对中华鳑鲏(Rhodeus sinensis)微卫星的分析中,二核苷酸占总微卫星位点的53.59%,其次为单核苷酸重复,二核苷酸重复中AC/GT类占60.63%,GC/CG仅占0.32%。在裸体异鳔鳅鮀 (Xenophysogobio nudicorpa)中[32],二核苷酸重复占总微卫星位点比例高达83.15%,AC/GT类重复占49.36%,GC/CG重复仅有4个。
多数鱼类开发的微卫星标记以二核苷酸重复为主,但研究表明,在人类基因组中三核苷酸重复序列与遗传疾病的发生有关,并且具有较高的多态性和遗传稳定性[35]。部分学者对三、四核苷酸重复微卫星标记的研究结果存在差异。房祖业等[28]对大刺鳅 (Mastacembelus armatus) 二、三、四核苷酸重复微卫星标记的筛选发现二核苷酸重复较三、四核苷酸重复具有更高的筛选效率和多态性;鲁翠云等[17]、谭照君等[18]、李文升等[19]的研究认为三、四核苷酸具有更高的多态性和分型效果。长体圆鲹二、三核苷酸的筛选效率分别为22.2%和29.0%,PIC分别为0.827 4和0.687 7 (表4)。就筛选效率而言,三核苷酸重复略高于二核苷酸重复,但二者相差不大。PIC为衡量种群遗传变异程度的重要指标[36],二核苷酸重复多态性明显高于三核苷酸重复。本文中长体圆鲹二核苷酸重复筛选效率低于三核苷酸重复,但多态性二核苷酸重复明显高于三核苷酸重复。因此,筛选效率和多态性的差异可能由种属差异或其他多种因素导致。
表 4 长体圆鲹二、三核苷酸重复微卫星标记的比较Table 4. Comparison on di- and trinucleotide-repeated microsatellite loci in D. macrosoma序列
sequence引物数
primer number重复次数
repeat number筛选效率
efficiencyPIC 二核苷酸重复
di-nucleotide-repeated18 9~14 22.2% 0.827 4 三核苷酸重复
tri-nucleotide-repeated9 6~10 29.0% 0.687 7 通过筛选的27对引物中18个位点为二核苷酸重复,重复次数为9~14次不等;9个位点为三核苷酸重复,重复次数为6~10次不等,符合Ellegren[37]提出的真核生物微卫星位点重复大部分在30次重复以下。但Ellegren[37]认为二核苷酸重复以15~19次为主,本文中高通量测序获得的二核苷酸重复主要在6~15次。基于Weber[38]的研究结果,重复次数高的微卫星在种群中表现出的多态性较高,龚小玲等[39]对澳洲鳗鲡 (Anguilla australis) 进行标记开发时发现,微卫星重复序列的重复次数过高会影响PCR效果,应选择居中的重复次数为宜。长体圆鲹二核苷酸重复PIC为0.827 4,具有较高多态性,表明选择6~15次的二核苷酸重复是合适的。
3.3 微卫星标记的种群遗传学特征
群体杂合度的高低反映了群体在多个基因座上的遗传变异及群体遗传多样性丰富度[19]。本研究中长体圆鲹中沙群体的平均Ho为0.631 7,平均He为0.796 8,说明长体圆鲹该群体的遗传多样性较高。平均Ho和He存在差异,说明存在杂合子缺失或者纯合子过剩的情况。PIC也是衡量群体遗传多样性的重要指数,Botstein等[36]认为基因标记PIC>0.5为高度多态位点,0.25<PIC<0.5为中度多态位点,PIC<0.25为低度多态性位点,通常不作为遗传多样性分析。本文中长体圆鲹位点除1个为中度多态外,其他位点均为高度多态位点。表明开发所得的长体圆鲹微卫星标记在中沙群体中具有较好的遗传稳定性和丰富的遗传多样性。
在所有27个位点中有8个位点偏离了“哈迪-温伯格”平衡,这些位点不适合进一步的遗传分析。近亲杂交、无效等位基因、种群退化和自然选择等因素皆可能导致微卫星位点偏离HWE[15]。
-
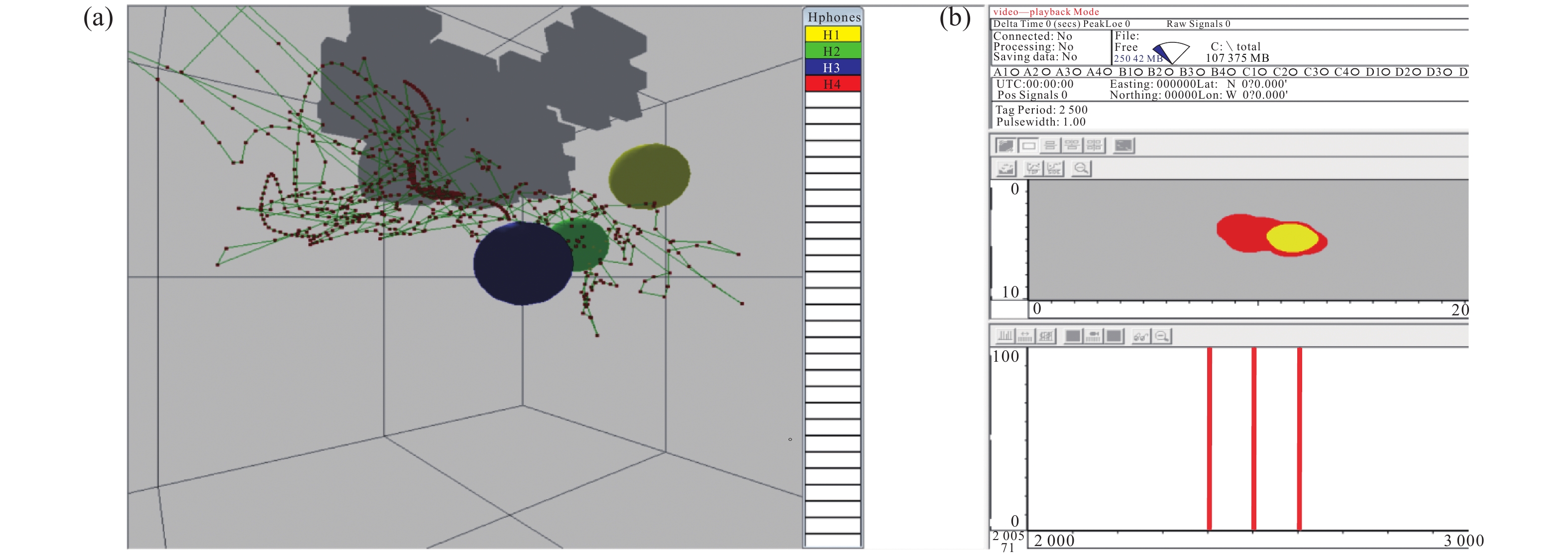
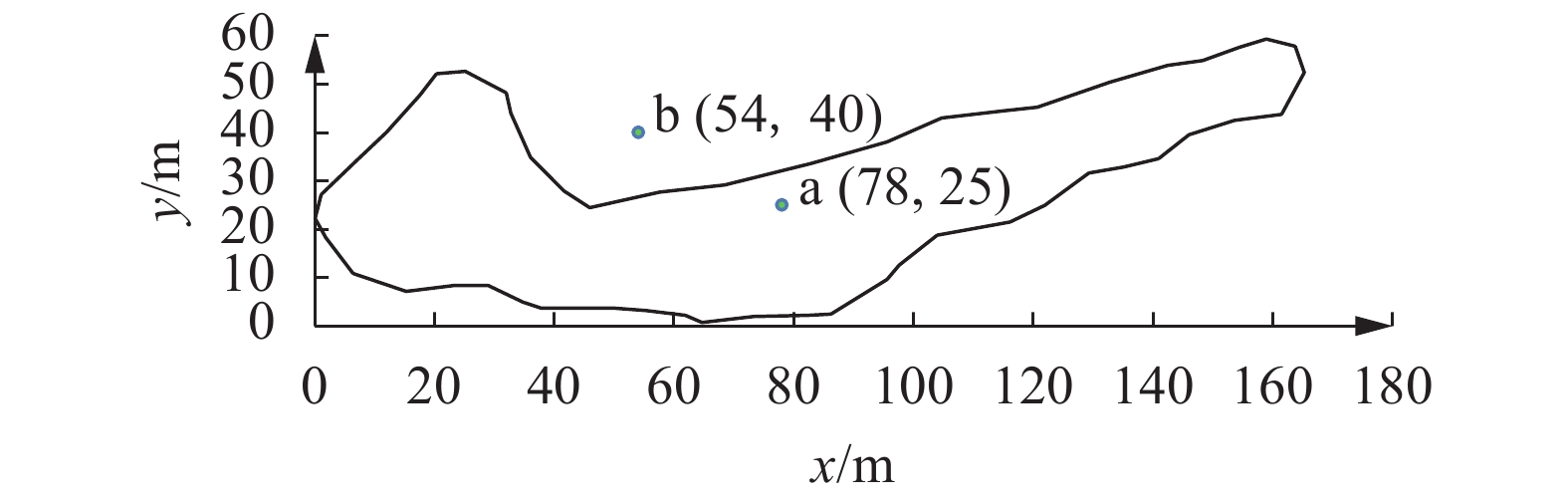
![]()
图 2 实时监测的鱼类三维 (a)、二维 (b) 运动轨迹
Figure 2. 3D (a) and 2D (b) motion trajectories of fish in real-time
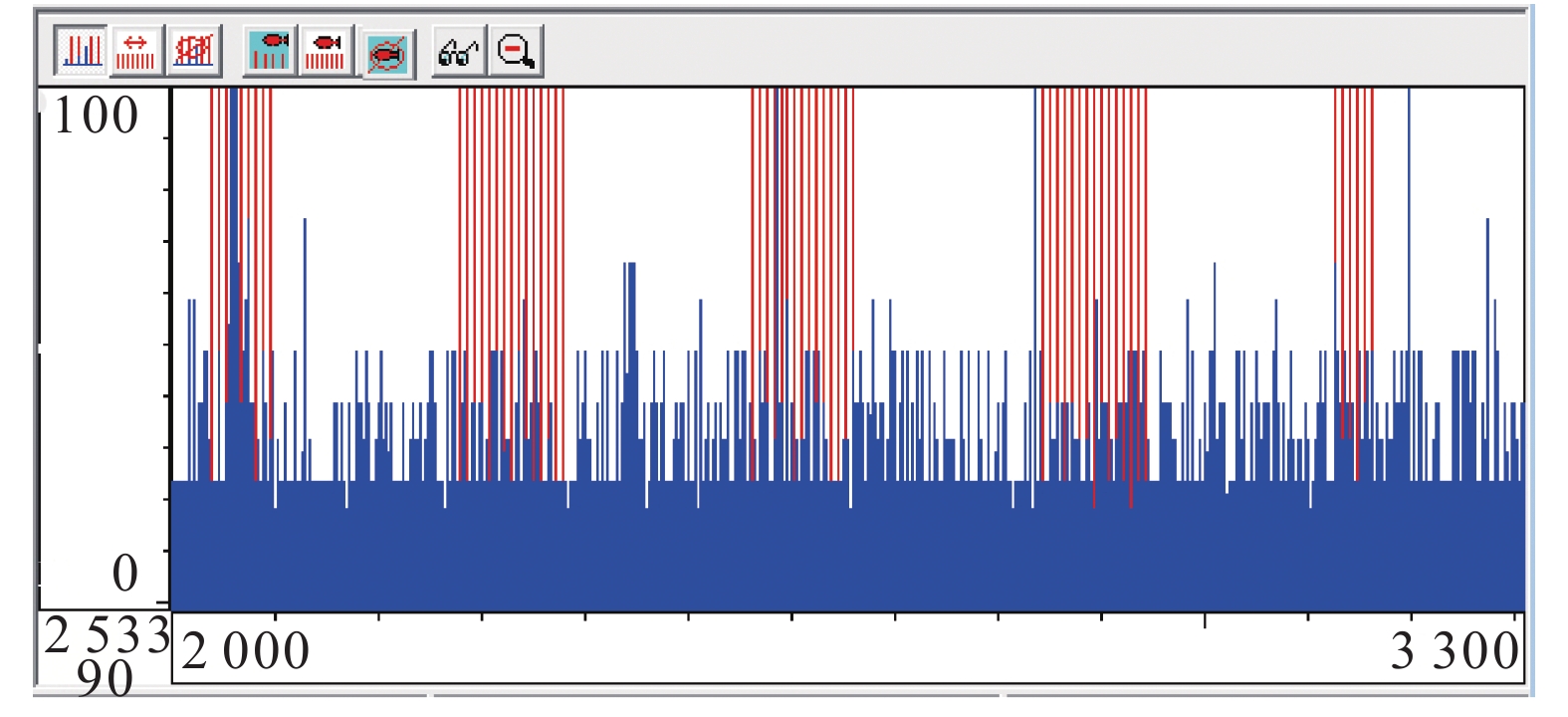
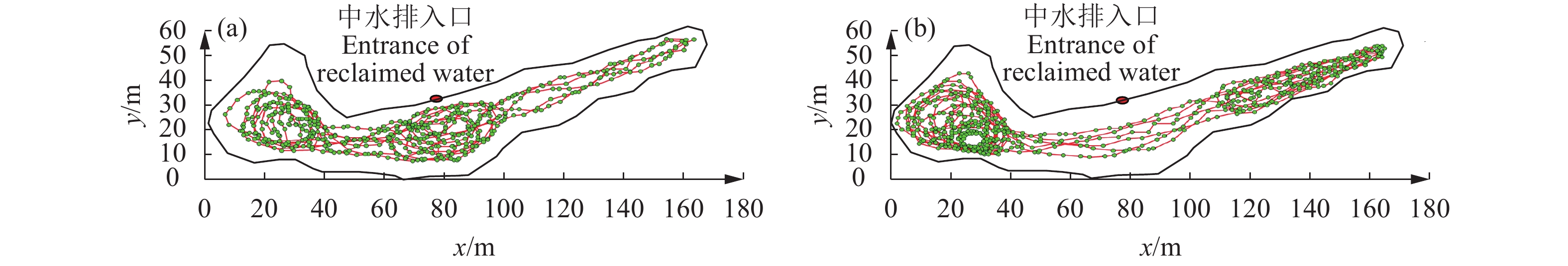
![]()
图 4 去噪处理后鱼类三维 (a)、二维 (b) 运动轨迹
Figure 4. 3D (a) and 2D (b) motion trajectories of fish after denoising
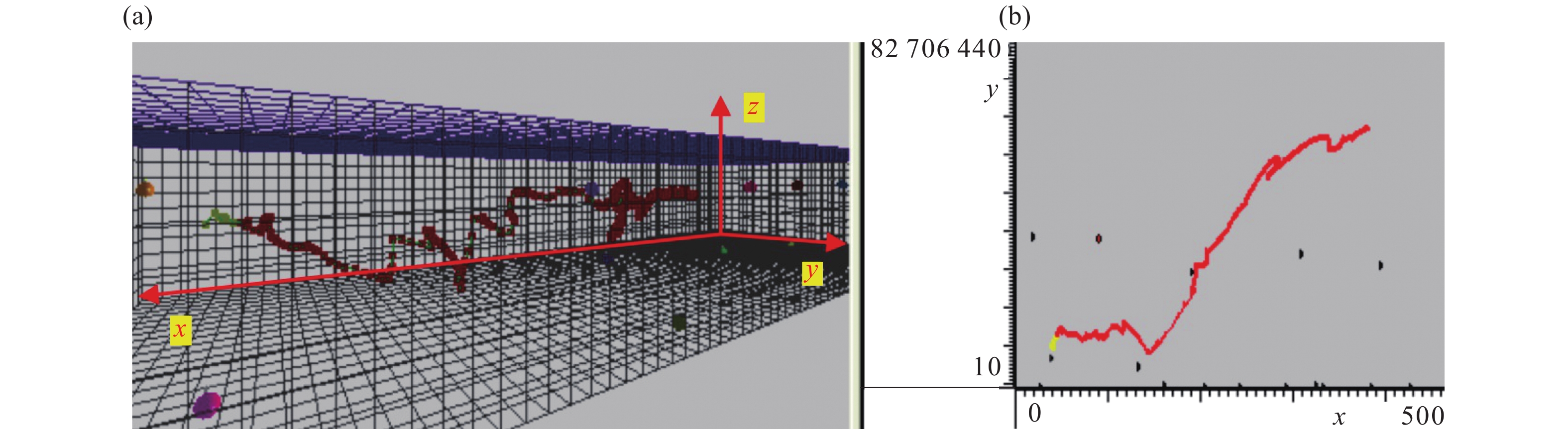
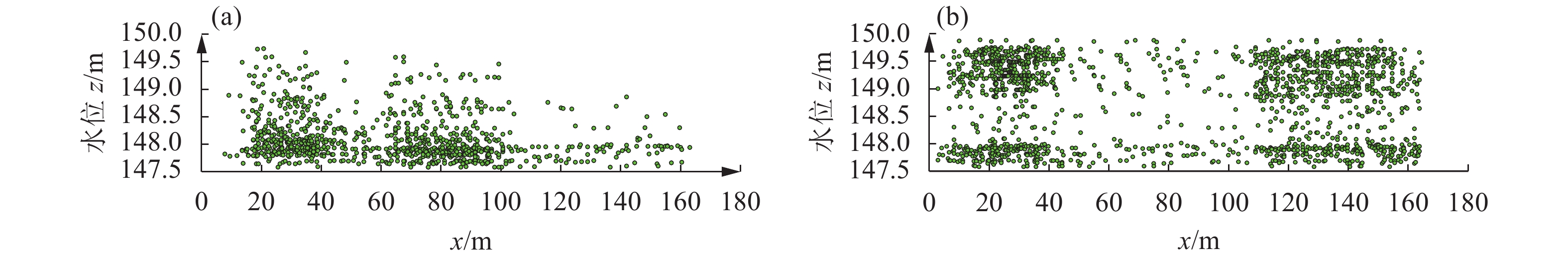
![]()
图 7 对缺失值进行均值插补后鱼类运动轨迹
Figure 7. Fish trajectories after mean interpolation of missing values
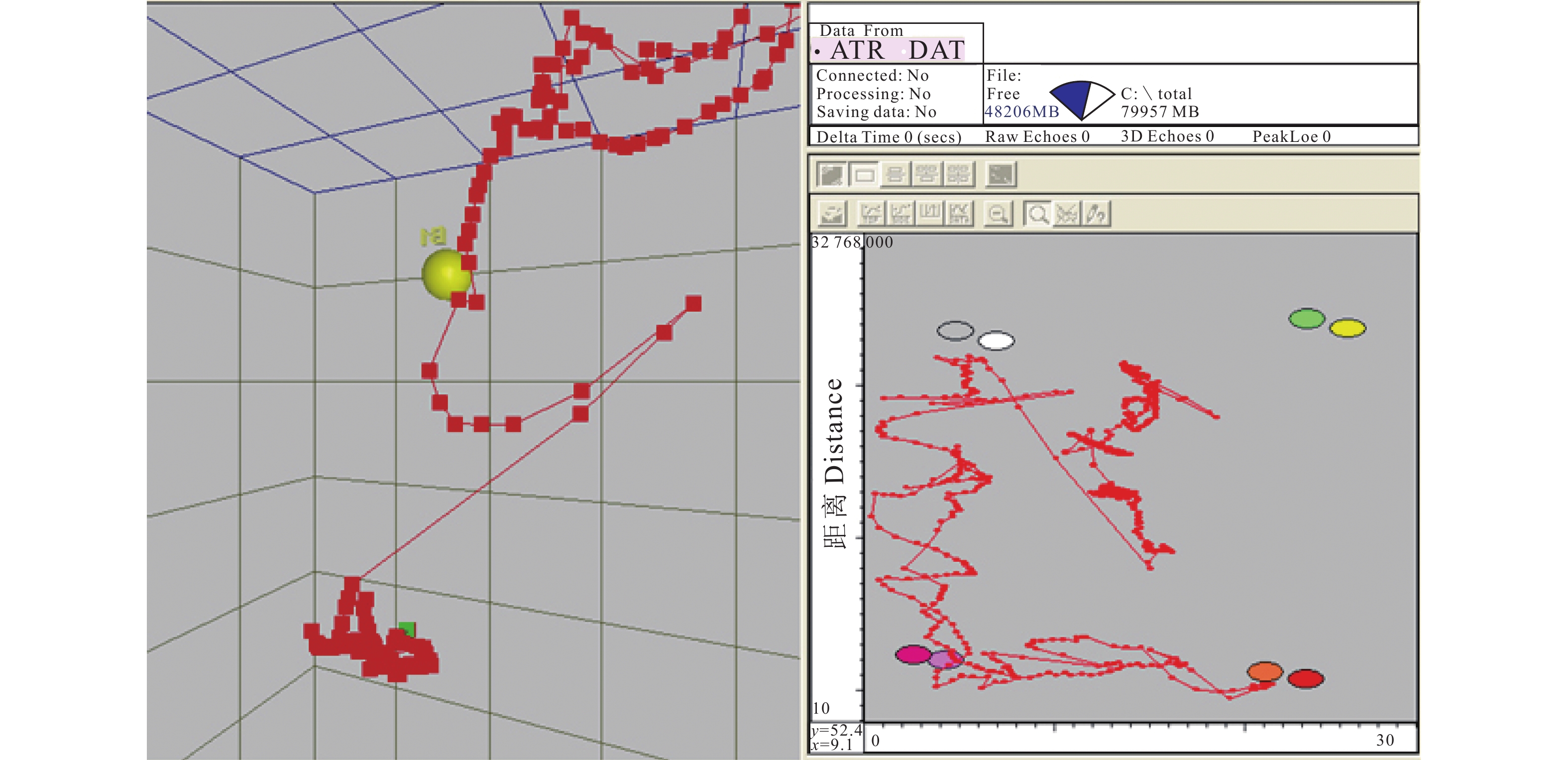
![]()
图 11 春季 (a) 和夏季 (b) 鱼类运动轨迹在水平面的分布情况
Figure 11. Distribution of fish motion trajectories in horizontal direction in spring (a) and summer (b)
-
[1] KANG I J, MOROISHI J, NAKAMURA A, et al. Biological monitoring for detection of toxic chemicals in water by the swimming behavior of small freshwater fish[J]. J Fac Agr Kyushu U, 2009, 54(1): 209-214.
[2] 王丽娜, 方景龙. 基于视频理解的活体鱼水质监测实验[J]. 杭州电子科技大学学报, 2010, 30(6): 36-38. doi: 10.3969/j.issn.1001-9146.2010.06.010 [3] 徐盼麟, 韩军. 基于视频的水箱内鱼类运动行为观测系统[J]. 湖南农业科学, 2011(19): 140-142. doi: 10.3969/j.issn.1006-060X.2011.19.043 [4] 江丹丹, 桂福坤. 基于视频图像的鱼类运动行为轨迹追踪[J]. 浙江海洋学院学报(自然科学版), 2015, 34(2): 112-118. [5] 范伟康. 基于鱼群行为分析的水质监测系统研究[D]. 杭州: 浙江工业大学, 2015: 13-14. [6] 余祥瑞. 基于双摄像机视频的生物水质预警系统的研究[D]. 宁波: 宁波大学, 2017: 5-6. [7] 颜鹏东, 谭均军, 高柱, 等. 基于视频跟踪的竖缝式鱼道内鱼类运动行为分析[J]. 水生生物学报, 2018, 42(2): 250-254. doi: 10.7541/2018.031 [8] 于铭, 杨士莪. 被动式渔探仪开发设计与试验研究[J]. 应用声学, 2007, 26(4): 231-238. doi: 10.3969/j.issn.1000-310X.2007.04.008 [9] 于铭, 杨士莪, 牟冬英. 被动式渔探仪的信号检测与方位估计[J]. 哈尔滨工业大学学报, 2008, 40(1): 147-151. doi: 10.3321/j.issn:0367-6234.2008.01.037 [10] STANTON, TIMOTHY K. 30 years of advances in active bioacoustics: a personal perspective[J]. Meth Oceanogr, 2012, s1/2: 49-77.
[11] GODLEWSKA M, SWIERZOWSKI A. Hydroacoustical parameters of fish in reservoirs with contrasting levels of eutrophication[J]. Aquat Living Resour, 2003, 16(3): 167-173. doi: 10.1016/S0990-7440(03)00014-7
[12] EVERSON I, TAABU M A, KAYANDA R. Acoustic estimates of commercial fish species in Lake Victoria: moving towards ecosystem-based fisheries management[J]. Fish Res, 2013, 139: 65-75. doi: 10.1016/j.fishres.2012.09.019
[13] 武智, 谭细畅, 李新辉, 等. 珠江首次禁渔西江段鱼类资源声学跟踪监测分析[J]. 南方水产科学, 2014, 10(3): 24-28. doi: 10.3969/j.issn.2095-0780.2014.03.004 [14] 武智, 李新辉, 李捷, 等. 红水河岩滩水库鱼类资源声学评估[J]. 南方水产科学, 2017, 13(3): 20-25. doi: 10.3969/j.issn.2095-0780.2017.03.003 [15] SZCZUCKA J, HOPPE T, SCHMIDT B, et al. Acoustical estimation of fish distribution and abundance in two Spitsbergen fjords[J]. Oceanologia, 2017, 59(4): 585-591.
[16] 张翔, 沈蔚, 童剑锋, 等. 基于DIDSON双频识别声纳技术的青草沙水库鱼类资源量评估[J]. 上海海洋大学学报, 2017, 26(4): 561-569. doi: 10.12024/jsou.20161201919 [17] CONTI S G, MAURER B D, ROUX P, et al. Acoustical monitoring of fish behavior in a tank[J]. J Acoust Soc Am, 2004, 116(4): 2489-2489.
[18] 张赞, 蔺丹清, 汤勇, 等. 基于声学测量方法的大伙房水库鱼类资源季节变动特征[J]. 南方水产科学, 2014, 10(6): 12-19. doi: 10.3969/j.issn.2095-0780.2014.06.002 [19] 孙明波, 谷孝鸿, 曾庆飞, 等. 基于水声学方法的天目湖鱼类季节和昼夜空间分布研究[J]. 生态学报, 2015, 23(9): 5597-5605. [20] 连玉喜, 叶少文, 黄耿, 等. 三峡库区香溪河鱼类资源水声学探测效果的昼夜差异研究[J]. 水生生物学报, 2015, 39(5): 1041-1045. doi: 10.7541/2015.136 [21] 李斌, 陈国宝, 于杰, 等. 海南陵水湾口海域不同季节鱼类资源声学探查[J]. 水产学报, 2018, 42(4): 544-555. [22] 郭杰. 航道整治透水框架群对鱼类集群影响的水声学探测[J]. 水生态学杂志, 2015, 36(5): 29-35. [23] 马燕芹, 司纪锋. 基于水声技术的黄海近海鱼类活动定点监测研究[J]. 渔业现代化, 2016, 43(4): 70-75. doi: 10.3969/j.issn.1007-9580.2016.04.013 [24] EHRENBERG J E, STEIG T W. A study of the relationship between tag-signal characteristics and achievable performances in acoustic fish-tag studies[J]. ICES J Mar Sci, 2009, 66(6): 1278-1283. doi: 10.1093/icesjms/fsp051
[25] 王志超, 陈国宝, 曾雷. 基于声学标志和无线跟踪方法的鱼类行为研究[J]. 南方水产科学, 2018, 14(2): 51-59. doi: 10.3969/j.issn.2095-0780.2018.02.007 [26] STEIG T W, TIMKO M A. Innovative techniques for analyzing the three-dimensional behavioral results from acoustically tagged fish[J]. J Acoust Soc Am, 2005, 117(4): 2367-2367.
[27] SEMMENS B X. Acoustically derived fine-scale behaviors of juvenile Chinook salmon (Oncorhynchus tshawytscha) associated with intertidal benthic habitats in an estuary[J]. Can J Fish Aquat Sci, 2008, 65(9): 2053-2062. doi: 10.1139/F08-107
[28] SKALSKI J R, STEIG T W, HEMSTROM S L. Assessing compliance with fish survival standards: a case study at Rock Island dam, Washington[J]. Environ Sci Policy, 2012, 18: 45-51. doi: 10.1016/j.envsci.2012.01.001
[29] 李荣辉.鱼类栖息迁徒习性及其监测技术研究[D]. 南宁: 广西大学, 2013: 75-94 [30] ABAID N, PORFIRI M. Collective behavior of fish shoals in one-dimensional annular domains[C]. Riverside: Institute of Electrical and Electronics Engineers, 2010 American Control Conference, Riverside: Institute of Electrical and Electronics Engineers,2010: 63-68.
[31] SCHULTZ A A, KUMAGAI K K, BRIDGES B B. Methods to evaluate gut evacuation rates and predation using acoustic telemetry in the tracy fish collection facility primary channel[J]. Anim Biotel, 2015, 3(1): 1-9. doi: 10.1186/s40317-014-0021-8
[32] HOLLO T, WATSON B M, JOHNSTON S V, et al. Behaviour of growth hormone transgenic coho salmon Oncorhynchus kisutch in marine mesocosms assessed by acoustic tag telemetry[J]. J Fish Biol, 2017, 90(4): 1660-1667. doi: 10.1111/jfb.13233
[33] ROMINE J G, PERRY R W, JOHNSTON S V, et al. Identifying when tagged fishes have been consumed by piscivorous predators: application of multivariate mixture models to movement parameters of telemetered fishes[J]. Anim Biotel, 2014, 2(1): 3. doi: 10.1186/2050-3385-2-3
[34] 黄月群, 蔡德所, 李明泉, 等. 大坝泄水对鱼类洄游能力的影响研究[J]. 人民长江, 2019, 50(8): 74-80. [35] CAI D S, LI R H. Research on fish habitat based on acoustic fish tracking system[J]. JCIT, 2012, 7(23): 195-201. doi: 10.4156/jcit.vol7.issue23.23
[36] HUANG Y Q, CAI D S, LI M Q, et al. Influence of changes in dissolved oxygen content on fish behavioral trajectories during water eutrophication[J]. Appl Ecol Env Res, 2018, 17(1): 653-666.
[37] 赵方旭. 不同水力条件下鱼类运动行为轨迹特性研究[D]. 南宁: 广西大学, 2016: 48-55 [38] KRISTIANSEN T S, FEMO A, HOLM J C, et al. Swimming behavior as an indicator of low growth rate and impaired welfare in Atlantic halibut (Hippoglossus hippoglossus L.) reared at three stocking densities[J]. Aquaculture, 2004, 230(1/2/3/4): 137-151.
[39] MCFARLANE W J, CUBITT K F, WILLIAMS H, et al. Can feeding status and stress level be assessed by analyzing patterns of muscle activity in free swimming rainbow trout (Oncorhynchus mykiss Walbaum)[J]. Aquaculture, 2004, 239(4): 467-484.
-
期刊类型引用(4)
1. 袁太平,廖宇琦,黄小华,胡昱,陶启友,李根,庞国良. 深水网箱网衣清洗装备关键部件设计及试验. 南方水产科学. 2024(01): 25-33 .  本站查看
本站查看
2. 魏盛军,蔡文鸿,丁兰,张哲,张艺,潘柏霖,陈思源,杨章武,魏观渊,林元俊,郑国富. 福建海上养殖设施污损生物处理方式现状调查. 渔业研究. 2023(02): 147-154 . 百度学术
3. 黄小华,庞国良,袁太平,胡昱,王绍敏,郭根喜,陶启友. 我国深远海网箱养殖工程与装备技术研究综述. 渔业科学进展. 2022(06): 121-131 . 百度学术
4. 廖宇琦,袁太平,胡昱,王绍敏,陶启友,黄小华. 转盘流道结构对网箱清洗装备喷嘴流场特性的影响. 渔业现代化. 2021(06): 9-18 . 百度学术
其他类型引用(3)
 下载:
下载:



计量
- 文章访问数: 3705
- HTML全文浏览量: 1623
- PDF下载量: 82
- 被引次数: 7
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
