
Seasonal variation of fish community structure in Zhelin Bay, the South China Sea
-
摘要:
为阐明南海柘林湾鱼类群落结构的动态变化,基于2011年4月(春季)、8月(夏季)、11月(秋季)和2012年2月(冬季)对南海柘林湾拖网调查的数据,对柘林湾鱼类群落结构季节动态、鱼类营养级指数季节变化进行了初步分析。结果显示,各季节调查共捕获鱼类99种,底层鱼类占49.49%,暖水性鱼类占78.78%,肉食性鱼类占79.79%。各季节质量百分比和数量百分比优势种以拟矛尾
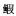 虎鱼(Parachaeturichthys polynema)、龙头鱼(Harpodon nehereus)、皮氏叫姑鱼(Johnius belengeri)等为主。柘林湾鱼类资源密度和资源尾数密度最高季节为夏季,各季节鱼类个体体质量大多小于15 g。春、秋季鱼类组成相对丰富,冬季生物量优势度最高。柘林湾鱼类以中级肉食性和高级肉食性为主,年度平均营养级指数为3.478,属于中级水平,夏季高营养级鱼类平均体质量最高。Abstract:
虎鱼(Parachaeturichthys polynema)、龙头鱼(Harpodon nehereus)、皮氏叫姑鱼(Johnius belengeri)等为主。柘林湾鱼类资源密度和资源尾数密度最高季节为夏季,各季节鱼类个体体质量大多小于15 g。春、秋季鱼类组成相对丰富,冬季生物量优势度最高。柘林湾鱼类以中级肉食性和高级肉食性为主,年度平均营养级指数为3.478,属于中级水平,夏季高营养级鱼类平均体质量最高。Abstract:Based on the date collected by quarterly otter trawl surveys in Zhelin Bay from 2011 to 2012, we analyzed the fish species composition, resources structure and trophic level. A total of 99 fishes were collected during the surveys. The proportions of demersal fish, tropical fish, and carnivorous fish were 49.49%, 78.78% and 79.79%, respectively. The composition of the dominant species of biomass and abundance percentages was similar, including Parachaeturichthys polynema, Harpodon nehereus and Johnius belengeri. The maximal biomass and abundance of fish were in summer in Zhelin Bay. The average body weight of fish in four seasons was less than 15 g. The composition of fish was more abundant in spring and autumn. The dominance of biomass of fish was the highest in winter. The feeding habit of fish in Zhelin Bay was mainly intermediate carnivorous and advanced carnivorous. The change of trophic level of four seasons was inconspicuously. The annual average trophic level index was 3.478 of intermediate level. It was in summer that the average body weight of high trophic level fish peaked.
-
Keywords:
- fish /
- community structure /
- seasonal variation /
- trophic level /
- Zhelin Bay of the South China Sea
-
凡纳滨对虾 (Litopenaeus vannamei) 因具有耐高温、耐低盐、生长快、抗病力强等优点,近年在我国内陆地区的养殖规模迅速扩大。根据养殖现状,很多从业者为了维持高生产力,投喂大量饵料,已远超对虾的需要量,并且长期使用抗生素预防和治疗病害[1]。过量投饵和滥用药物等不当措施会引起水质逐渐恶化并滋生大量病原体,在养殖中后期尤为严重[2],会引发病害并造成经济损失。水产养殖池塘水环境主要受人为调控,如何通过环境控制策略改善水质环境、降低病害发生,是目前生产上亟待解决的问题。
浮游细菌作为有机物的分解者和高营养级的食物源,可促进碳 (C)、氮 (N)、磷 (P)和硫 (S) 的生物化学循环[3]以及其他一些生态过程[4],是淡水生态系统物质循环和能量流动的主要驱动因子[5-6]。作为水环境不可或缺的重要组成部分,浮游细菌会直接影响对虾的健康状况[7]。因此,生产中浮游细菌的调控是人工养殖水体环境控制策略的核心。浮游细菌群落多样性是重要研究方向之一,生物多样性的变动会对水生态系统各项生化反应产生重要影响,并且群落多样性会对水环境状态、污染等情况迅速响应[8],可用于水环境变化的监控。
已有研究表明,不同水体浮游细菌群落结构及多样性特征存在明显差异[9-11],并受到水体不同理化因子的影响[12-13],其中有关对虾养殖池塘的报道主要集中在我国沿海地区,且多为海水养殖池塘[14-17]。因此,对我国内陆干旱、半干旱地区开展对虾养殖池塘浮游细菌群落和多样性研究,具有显著的地域特色和生态学意义。本文通过对新疆凡纳滨对虾淡化池塘水体采样调查,阐述了浮游细菌群落结构特征和多样性变化,并分析其对不同水体理化因子的响应程度及差异,以期为进一步开展凡纳滨对虾养殖池塘水质调控技术研究提供科学依据,推动西北内陆地区对虾产业的健康可持续发展。
1. 材料与方法
1.1 池塘条件及样品采集
试验区位于新疆维吾尔自治区昌吉市森淼渔业专业合作社,随机选择3口池塘 (编号分别为A、B和C) 进行监测,池塘面积均约为6 667 m2,凡纳滨对虾苗种首先在池塘内设的温棚中进行淡化养殖,温棚养殖密度约为 2 000~3 000 尾·m−3,入塘初始盐度为18.2‰,淡化期每天降低2‰~3‰左右,至盐度0.8‰左右。淡化后拆除池塘内的温棚逐渐升高水位,至平均水深约1.6 m,池塘养殖阶段3口池塘的放养密度均约为45 尾·m−3。
池塘A从温棚淡化养殖投放苗种当天开始取样,池塘B和C从虾苗淡化后的池塘养殖当天开始第一次采样,养殖期间每周采样1 次,固定采样时间为上午12:00左右。池塘A水深大于1.5 m时进行分层采样,每层水样形成一个样方,表层样品在水下20 cm处采集,底层样品距池底约20 cm,中层为水深的1/2处取样。池塘B和C每次仅采集表层菌样。采水器置于所需水层,弃去前端水样后将样品收集到500 mL无菌采样瓶中,4 ℃车载冰箱保存带回实验室进行样品的处理。池塘A各样品编号记作S1—S25,池塘B各样品编号记作S26、S28、S30、S32、S34、S36、S38、S40、S42、S44、S46、S48,池塘C各样品编号记作S27、S29、S33、S35、S37、S39、S41、S43、S45、S47、S49。
现场采集浮游细菌样品的同时,同位测定水体理化参数。理化因子包括水温、pH、溶解氧 (DO)、铵态氮 (NH4-N)、亚硝酸态氮 (NO2-N)、硝酸态氮 (NO3-N)、活性磷 (PO4-P)、可溶性硅酸盐 (SiO3-Si)、高锰酸盐指数 (CODMn)、叶绿素a (Chl-a)、硫化物 (Sul)、矿化度、碱度 (ALK) 和水总硬度等14项指标。
采用哈希HQ30d便携式溶解氧测试仪现场测定水温和溶解氧,哈希HQ11d型pH检测仪测定pH值,其他水化学参数取5 L水样低温保存带回实验室检测。测定方法按照《水和废水检测分析方法 (第四版)》[18]进行测定。理化因子实验结果以“平均值±标准差 (
$ \overline X \pm {\rm{SD}} $ )”表示。1.2 DNA提取与测序
每个样品采用孔径为0.22 μm的无菌滤膜 (WondaDisc,MCE水系,50 mm,中国) 进行过滤,滤液约200 mL,为避免微藻等其他微生物的影响,每个样品使用2~3个滤膜进行浮游细菌收集,以保证滤膜的通透性。然后样品迅速送至生工生物工程 (上海) 股份有限公司,以OMEGA试剂盒E.Z.N.A™ Mag-Bind Soil DNA Kit试剂盒提取浮游细菌DNA。第一轮采用引物314F (CCTACGGGNGGCWGCAG) 和805R (GACTACHVGGGTATCTAATCC) 对浮游细菌V3—V4区进行扩增,反应体系为30 μL,包含2×Hieff® Robust PCR Master Mix 15 μL、正反向引物各1 μL、模板DNA 15 ng,补充ddH2O至30 μL,PCR反应条件为 95 ℃ 3 min;5×(94 ℃, 30 s; 45 ℃, 20 s; 65 ℃, 30 s);20×(94 ℃, 20 s; 55 ℃, 20 s; 72 ℃, 30 s);72 ℃10 min;10 ℃直到停止。第二轮扩增引入Illumina桥式PCR兼容引物,反应体系和条件与第一轮相同。后使用MiSeq测序仪 (Illumina Inc. San Diego,美国) 进行高通量测序。
1.3 数据分析
使用cutadapt软件去除Read1 3'端测序引物接头和Read2 3'端测序引物接头;根据PE reads之间的重叠关系使用PEAR将成对reads拼接成一条序列;根据各样本barcode序列和引物序列从拼接后数据中分割出各样本数据,并校正序列方向;使用PRINSEQ软件切除reads尾部质量值20以下的碱基,设置10 bp的窗口,过滤质控后的含N序列和短序列,最终过滤掉低复杂度的序列,得到各样本有效数据。基于有效数据,利用Usearch软件[19],对各样本优化序列提取非重复序列,便于降低分析中间过程冗余计算量,所有样本去冗余序列合并后去除没有重复的单序列,将具有不小于97%相似性的序列进行聚类,形成不同的操作分类单元(Operational taxonomic units, OTU)。使用RDP classifier比对RDP数据库,获得每个OTU代表序列的分类学信息。基于OTU,利用R语言vegan包和picante包计算优势度、系统发育多样性指数和群落多样性指数等。利用pheatmap包绘制优势菌群组成热图,用ggrepel包、ggplot2包和vegan包等进行非度量多维标度 (Non-metric multidimensional scaling, NMDS) 和冗余分析 (Redundancy analysis, RDA),用anosim函数进行群落差异显著性比较,利用aov函数进行组间数据方差分析。通过Galaxy在线分析平台 (http://huttenhower.sph.harvard.edu/galaxy/) 对属水平的相对丰度矩阵进行线性判别分析 (Linear discriminant analysis, LDA),并识别不同组间统计学上潜在的标志性功能菌群 (biomarkers)。非参数因子克鲁斯卡尔-沃利斯和秩验检 [Kruskal-Wallis (KW) sum-rank test] α为0.05,LDA得分阈值为4.0。因池塘A温棚养殖阶段与池塘养殖阶段水环境和养殖密度等差异较大,在进行群落多样性和RDA分析比较中,去除了池塘A温棚养殖阶段的S1、S2和S3样方。
优势度 (Y) 用来评价浮游细菌的优势类群[20],计算公式为:
$$ {\text{Y}} = \frac{{{n_i}}}{N} \cdot {f_i} $$ (1) 式中:ni为第i属调查期间的相对丰度;N为浮游细菌总相对丰度;fi为该属出现的频率。
系统发育多样性的计算选用广泛使用的Faith系统发育多样性指数 (PD)[21],按下式计算:
$$ {\rm{PD}} = \sum\limits_{\{ {\text{c}} \in {\text{C}}\} } {{{\text{L}}_{\text{C}}}} $$ (2) 式中:C为连接系统发育树上所有物种的最短路径上的所有分支之和;c为连接节点的一段分支;LC为C的分支长度。
物种多样性指数的计算采用 Shannon指数 (H')[22] ,计算公式为:
$$ {\text{H}{{{\text{'}}}}} = - \sum\limits_{i = 1}^S {{P_i}{{\log }_2}{P_i}} $$ (3) 式中:S为群落中的总物种数;Pi为第i种的个体数与样品中总个数的比值。
2. 结果
2.1 主要理化因子状况
各池塘14项理化因子状况见表1。各池塘DO质量浓度基本保持在较高水平,变幅介于6.15~10.48 mg·L−1。各池塘NH4-N、NO3-N、NO2-N和PO4-P浓度养殖前、中期变动不大,后期显著升高,其中池塘A中NO2-N质量浓度显著高于池塘C (P<0.05),池塘B中PO4-P质量浓度显著高于池塘A (P<0.01)。各池塘SiO3-Si质量浓度均值分别为11.2、12.38和4.03 mg·L−1,池塘C中SiO3-Si质量浓度显著低于池塘A和B (P<0.01),各池塘SiO3-Si质量浓度在养殖前、中期较高 (最高达17.2 mg·L−1),后期显著降低 (最低值为0.88 mg·L−1)。各池塘ALK质量浓度均值分别为138.18、115.32和84.46 mg·L−1,各池塘ALK质量浓度差异极显著 (P<0.01)。其余各变量在各池塘间差异均不显著 (P>0.05)。
表 1 凡纳滨对虾养殖池塘水体主要理化因子Table 1. Environmental factors in L. vannamei ponds环境因子
Environmental factor池塘A
Pond A池塘B
Pond B池塘C
Pond C水温 Temperature/℃ 26.19±2.01 26.15±1.57 26.37±2.00 pH 8.10±0.50 8.29±0.37 8.30±0.38 溶解氧质量浓度 DO/(mg·L−1) 8.00±0.68 8.45±1.17 8.11±1.15 铵态氮质量浓度 NH4-N/(mg·L−1) 0.45±0.73 0.44±0.39 0.33±0.27 亚硝酸氮质量浓度 NO2-N/(mg·L−1) 0.01±0.01 0.01±0.01 0.01±0.01 硝酸态氮质量浓度 NO3-N/(mg·L−1) 0.34±0.44 0.41±0.42 0.48±0.53 活性磷质量浓度 PO4-P/(mg·L−1) 0.12±0.14 0.37±0.24 0.22±0.28 活性硅酸盐质量浓度 SiO3-Si/(mg·L−1) 11.2±3.55 12.38±7.38 4.03±2.98 高锰酸盐指数 CODMn/(mg·L−1) 8.34±7.19 6.12±3.18 5.34±2.91 叶绿素 a 质量浓度 Chl-a/(mg·L−1) 75.78±131.01 56.13±42.81 58.36±61.48 硫化物质量浓度 Sul/(mg·L−1) 0.03±0.04 0.02±0.01 0.02±0.01 矿化度质量浓度 Mineralization degree/(mg·L−1) 1209.44±2445.58 813.39±171.69 814.35±41.91 碱度 ALK/(mg·L−1) 138.18±12.06 115.32±33.82 84.46±14.73 总硬度 Total hardness/(mg·L−1) 362.76±395.28 476.5±227.34 349.36±60.18 2.2 浮游细菌群落结构及变动
2.2.1 各池塘浮游细菌门水平群落组成
48个浮游细菌样品共测得3 403 929条优化序列,平均69 467条,变幅介于41 169~100 945 条,序列长度介于350~476 bp (表2)。数据分析共获得2 854个OTU (97%相似性),序列比对发现古细菌2门1纲1科1属,细菌30门59纲98目199科433属。池塘A调查期间共鉴出浮游细菌29门,相对丰度占比前9位的门类共占99.5% (变幅为0.21%~38.8%),按相对丰度占比依次为变形菌门、放线菌门、蓝细菌门、拟杆菌门、疣微菌门、浮霉菌门、厚壁菌门、Parcubacteria、芽单胞菌门 (图1-a);池塘B调查期间共鉴出浮游细菌24门,前9位的门类相对丰度占比达99.5% (变幅介于0.3%~43.4%),排在前9位的门类中除没有Parcubacteria,而出现Candidatus Saccharibacteria (占比0.3%)外,其余门类同池塘A (图1-b);池塘C调查期间共鉴出浮游细菌25门,前9门相对丰度占比达99.5% (变幅介于0.2%~44.3%),前9位出现的浮游细菌门类在池塘A和B中均有发现 (图1-c)。
表 2 各样本有效序列数据统计Table 2. Valid sequences of each sample样品
Sample条形码
Barcode有效序列
Valid sequence/条碱基数
Base number平均长度
Mean length/bp最短序列长度
Min. length/bp最长序列长度
Max. length/bpS1 GTAACA 82 736 34 604 966 418.26 363 469 S2 CCAGAC 81 905 34 619 470 422.68 361 476 S3 GGTGAA 56 859 23 683 959 416.54 367 475 S4 TGCATC 85 212 35 667 198 418.57 353 469 S5 TCGACC 83 246 34 547 936 415.01 365 470 S6 GTCGCG 73 829 30 547 465 413.76 362 453 S7 CGGATG 83 662 34 668 763 414.39 356 465 S8 GTGAAA 84 384 34 811 366 412.54 350 466 S9 ATCTTG 100 945 41 590 813 412.01 350 452 S10 TATGCA 73 801 30 659 241 415.43 352 459 S11 GTAACA 86 471 35 924 249 415.45 354 471 S12 GCGAGG 90 432 37 722 526 417.14 351 465 S13 CACGAT 54 439 22 735 194 417.63 373 465 S14 GCGGTA 44 227 18 478 585 417.81 352 471 S15 TATCGA 61 431 25 638 627 417.36 359 473 S16 ATCACG 52 384 21 792 829 416.02 376 471 S17 CGGATG 99 211 41 215 038 415.43 360 473 S18 CGCATA 100 001 41 427 908 414.27 372 476 S19 TGCATC 65 712 27 246 907 414.64 359 476 S20 TCAGTA 76 029 31 690 841 416.83 362 471 S21 CGGCAC 75 202 31 405 455 417.61 364 469 S22 ATCACG 71 605 30 328 221 423.55 352 470 S23 CGGATG 63 358 26 635 233 420.39 367 470 S24 GTGAAA 56 429 23 698 925 419.98 350 471 S25 TCAGTA 92 400 38 487 742 416.53 355 435 S26 GAAGTG 87 305 37 043 958 424.31 368 448 S27 TCGACC 94 075 39 472 386 419.58 360 464 S28 CTTGTA 52 273 22 054 548 421.91 373 472 S29 GTTTCG 44 697 18 684 447 418.02 360 462 S30 ATCTTG 59 737 25 072 871 419.72 365 465 S32 GCCATC 78 462 32 993 239 420.5 357 474 S33 TGTGTT 69 613 29 167 734 419.00 351 474 S34 CTTGTA 56 083 23 551 548 419.94 356 471 S35 GTTTCG 45 701 18 993 334 415.60 359 464 S36 TTCGTA 46 337 19 212 573 414.63 356 468 S37 CCAGAC 51 978 21 598 326 415.53 372 436 S38 AGCAGT 77 233 31 835 991 412.21 350 470 S39 GAGGAA 75 226 31 060 291 412.89 370 468 S40 AAGGTA 46 920 19 633 227 418.44 352 450 S41 ATCACG 43 983 18 242 559 414.76 352 469 S42 TAGGAC 66 639 27 702 230 415.71 356 470 S43 TGGACG 49 206 20 369 621 413.97 357 472 S44 AGAACA 50 322 20 765 128 412.65 356 469 S45 GGTGTG 41 169 17 011 770 413.22 350 471 S46 AACTAT 67 691 28 070 176 414.68 357 468 S47 ACTGCG 60 220 25 559 117 424.43 359 474 S48 TGTGTT 94 261 39 467 506 418.7 355 474 S49 TAGGAC 86 904 36 530 365 420.35 353 467 ![]() 图 1 凡纳滨对虾养殖池塘门水平的浮游细菌组成Figure 1. Composition of bacteriaoplankon at phylum level in L. vannamei ponds
图 1 凡纳滨对虾养殖池塘门水平的浮游细菌组成Figure 1. Composition of bacteriaoplankon at phylum level in L. vannamei ponds2.2.2 各池塘优势菌群及相对丰度
利用优势度指数筛选各池塘优势菌群,本文将Y值大于0.01的浮游细菌列为优势菌群。3口池塘共筛选出优势菌群23属,其中池塘A有优势菌群18属,池塘B 15属,池塘C 15属。各池塘优势菌群相对丰度变动情况如图2所示[为更加清晰地展示优势菌群的变动情况,相对丰度数据进行了log2(x+0.01)转换]。根据层级聚类分析,池塘A优势菌群变动情况可提取3类 (图2-a):I类中Gplla和微酸菌属在淡化期 (S2—S4) 含量较低 (0.003~2.928),池塘养殖初期相对丰度较高 (最大值为38.508),之后逐渐降低;II类在淡化期和池塘养殖中、后期相对丰度较高;III类优势菌群相对丰度多在池塘养殖初期和后期 (S19—S25) 较高 (最高可达49.9%)。池塘B (图2-b) 中I类优势菌群在养殖期不同阶段均可达到较高的相对丰度 (最高值分别可达41.3%和39.5%);II类在养殖中后期 (S36—S48) 可占据一定相对丰度 (3.8%~21.1%);III类多在养殖前期呈现较高的相对丰度 (最高值可达18.2%)。池塘C (图2-c)中I类优势菌群多在养殖中期 (S33—S41) 出现较高相对丰度 (最高可达19.3%);II类优势菌群则在养殖初期 (最高值为38.2%) 和后期 (38.5%) 相对丰度较高;III类优势菌群在养殖前期相对丰度较高 (最高值为7.5%)。
2.3 浮游细菌群落Alpha多样性
利用PD值、H'值和物种丰富度对不同池塘alpha多样性的变动情况进行了计算 (图3)。池塘A、B和C的PD指数均值分别为82.81 (61.94~111.65)、74.65 (35.55~106.19) 和59.56 (24.45~78.14),方差分析表明池塘A和C间差异极显著 (P<0.01),其他各池塘间差异不显著 (P>0.05)。H'值各池塘均值分别为4.28 (3.04~4.96)、4.01 (2.64~5.06) 和3.71 (2.48~4.59),池塘A与C之间差异均极显著 (P<0.01)。各池塘物种丰富度均值分别为843 (变幅介于597~1054)、656 (变幅介于334~942)和620 (变幅介于243~743),池塘A与B和C之间差异均极显著 (P<0.01),池塘B和C之间差异不显著 (P>0.05)。
![]() 图 3 3口池塘的α多样性系数分析注:箱体上中下线分别为75、50 (中位数) 和25分位数,轴须线最长不超过1.5倍箱体范围,黑色空心圆表示平均数;差异显著性用* (P<0.05)、** (P<0.01)以及*** (P<0.001) 表示;图中的样本量:A:n=22、B:n=12、C:n=12。Figure 3. α diversity index analysis of bacterioplankton in three pondsNote: The upper, middle and lower lines of the box are 75, 50 (Median) and 25 quantiles, respectively. The maximum length of whiskers shall not exceed 1.5 times of the box range. The black hollow circles represent the average values. The significant differences were represented by * (P<0.05), ** (P<0.01) and *** (P<0.001). The numbers of replicated samples in this figure are: A: n=22; B: n=12; C: n=12.
图 3 3口池塘的α多样性系数分析注:箱体上中下线分别为75、50 (中位数) 和25分位数,轴须线最长不超过1.5倍箱体范围,黑色空心圆表示平均数;差异显著性用* (P<0.05)、** (P<0.01)以及*** (P<0.001) 表示;图中的样本量:A:n=22、B:n=12、C:n=12。Figure 3. α diversity index analysis of bacterioplankton in three pondsNote: The upper, middle and lower lines of the box are 75, 50 (Median) and 25 quantiles, respectively. The maximum length of whiskers shall not exceed 1.5 times of the box range. The black hollow circles represent the average values. The significant differences were represented by * (P<0.05), ** (P<0.01) and *** (P<0.001). The numbers of replicated samples in this figure are: A: n=22; B: n=12; C: n=12.2.4 浮游细菌群落Beta多样性
利用rankindex函数筛选出最优的manhattan差异系数进行NMDS分析,如图4所示,各池浮游细菌样方95%置信区间 (不同颜色椭圆所示) 有较多重叠,且这些样方多为养殖的中、后期,边缘分布的诸多样方多为养殖初期,表明各池塘浮游细菌群落在对虾养殖初期差异较大,至中、后期差异减小。ANOSIM分析结果显示,各池塘浮游细菌群落结构总体上差异不显著 (P>0.05)。
![]() 图 4 凡纳滨对虾养殖池塘浮游细菌群落NMDS分析Figure 4. NMDS analysis of bacterioplankton community in L. vannamei pond
图 4 凡纳滨对虾养殖池塘浮游细菌群落NMDS分析Figure 4. NMDS analysis of bacterioplankton community in L. vannamei pond3口池塘调查期间共鉴出浮游细菌433属,其中3口池塘共有属414个,池塘A特有属88个,池塘B特有属7个,池塘C特有属4个 (图5)。利用LEfSe分析法,对各池塘浮游细菌群落相对丰度在各分类水平上的差异进行了分析,LDA得分阈值设为4时 (P=0.05),共筛选出18个分类单元,其中属于池塘A的差异分类单元4个,属于池塘B和C的差异分类单元均为7个。如图6所示,差异分析结果包含各分类水平的差异菌群信息,其中池塘A的差异菌群为Lacibacter sp.、鞘脂杆菌纲、成对杆菌属和微杆菌科;池塘B的差异菌群为暖绳菌纲、暖绳菌目、SAR11、动球菌科、鞘脂单胞菌科、远洋杆属和α-变形菌纲;池塘C的差异菌群为类芽孢八叠球菌、嗜甲基菌科、嗜甲基菌目、黄杆菌纲、浮霉菌门、分枝杆菌属和分支杆菌科。
![]() 图 5 各池浮游细菌属水平上共有和特有属的数量Figure 5. Number of common and endemic genera of bacterioplankton at genus level in each pond
图 5 各池浮游细菌属水平上共有和特有属的数量Figure 5. Number of common and endemic genera of bacterioplankton at genus level in each pond![]() 图 6 各池浮游细菌群落的线性判别分析 (菌群LDA>4)Figure 6. Linear discriminant analysis of bacterioplankton community in each pond (Bacterial flora with LDA>4)
图 6 各池浮游细菌群落的线性判别分析 (菌群LDA>4)Figure 6. Linear discriminant analysis of bacterioplankton community in each pond (Bacterial flora with LDA>4)2.5 优势菌群分布与主要理化因子的关系
对14个环境变量与优势菌群 (Y>0.02) 进行冗余分析 (RDA)。前两轴累计解释总变异的37.8%。经P值校正,全模型前2约束轴显著 (P<0.05)。经ordiR2step函数向前选择,最终保留4个环境变量,其中PO4-P与第一主成分轴 (RDA1) 相关性最大 (R=0.775),其次是ALK (R=−0.604) 和DO (R=0.568),Sul与第二主成分轴相关性最大 (R=0.737)。
如图7所示,GpIIa、芽殖杆菌属、湖栖菌属和红杆菌属与ALK呈正相关,与DO和PO4-P呈负相关;不动杆菌属和黄杆菌属与PO4-P呈高度正相关,与ALK呈高度负相关;气单胞菌属和鞘脂菌属与Sul和DO均呈正相关,Spartobacteria genera incertae sedis和多核杆菌与Sul呈负相关。池塘A养殖前、中期的样方主要集中在排序图左部,显示优势菌群主要在ALK浓度较高的样方分布,其中GpIIa是主导的优势菌群,池塘A后期的样方PO4-P浓度高。池塘B和C在养殖初期和后期的样方多数PO4-P浓度较高,且H'较低 (H'均值分别为3.08和3.50);养殖中期的样方,H'较高 (H'均值为4.30,P<0.01)。
![]() 图 7 优势菌群分布和群落多样性 (H') 与主要理化因子之间的关系 (II型标尺)注:Aci. 不动杆菌属;Aer. 气单胞菌属;Fla. 黄杆菌属;Gem. 芽殖杆菌属;GpI. GpIIa;Ilu. 微酸菌属;Lim. 湖栖菌属;Pol. 多核杆菌;Rho. 红杆菌属;Sed. 沉积物杆状菌属;Spa. 发光细菌属;Sph. 鞘脂菌属。Figure 7. Species associations of dominan flora and diversity (H') with environmental factors (Scaling II)Note: Aci. Acinetobacter sp.; Aer. Aeromonas sp.; Fla. Flavobacterium sp.; Gem. Gemmobacter sp.; GpI. GpIIa; Ilu. Ilumatobacter sp.; Lim. Limnohabitans sp.; Pol. Polynucleobacter sp.; Rho. Rhodobacter sp.; Sed. Sediminibacterium sp.; Spa. Spartobacteria genera incertae sedis; Sph. Sphingomonas sp..
图 7 优势菌群分布和群落多样性 (H') 与主要理化因子之间的关系 (II型标尺)注:Aci. 不动杆菌属;Aer. 气单胞菌属;Fla. 黄杆菌属;Gem. 芽殖杆菌属;GpI. GpIIa;Ilu. 微酸菌属;Lim. 湖栖菌属;Pol. 多核杆菌;Rho. 红杆菌属;Sed. 沉积物杆状菌属;Spa. 发光细菌属;Sph. 鞘脂菌属。Figure 7. Species associations of dominan flora and diversity (H') with environmental factors (Scaling II)Note: Aci. Acinetobacter sp.; Aer. Aeromonas sp.; Fla. Flavobacterium sp.; Gem. Gemmobacter sp.; GpI. GpIIa; Ilu. Ilumatobacter sp.; Lim. Limnohabitans sp.; Pol. Polynucleobacter sp.; Rho. Rhodobacter sp.; Sed. Sediminibacterium sp.; Spa. Spartobacteria genera incertae sedis; Sph. Sphingomonas sp..3. 讨论
3.1 凡纳滨对虾淡化养殖池塘浮游细菌群落结构特征及其对理化因子的响应
本次调查的新疆凡纳滨对虾淡化养殖池塘共鉴出浮游细菌30门,各池塘相对丰度占比前7的门类 (相对丰度占比>1%) 组成相同,分别为变形菌门、放线菌门、蓝细菌门、拟杆菌门、疣微菌门、浮霉菌门和厚壁菌门。这与已有报道的不同凡纳滨对虾养殖水体中优势浮游细菌组成在门分类水平上较为相似[13-15,23]。
群落中优势种群的存在对于生态系统结构与功能的稳定至关重要,包含系统代谢所需的主要生物结构与功能信息,并影响生态系统的演替[23]。如本次调查发现,Lacibacters sp. 既是池塘A的优势菌群也是LEfSe分析识别到的一个生物标记,Lacibacters sp. 经常发现于富营养化水体中[24],其不能进行反硝化和硝酸盐还原,但能水解复杂的有机物,使降解的有机物能够被微生物群落充分吸收利用[25]。表明池塘A水生态系统在有机物循环利用转化中具有优势,更有利于形成良好的养殖环境。
研究表明水生态系统中浮游细菌群落对水环境变化极其敏感并可迅速做出响应[26-27],这种响应主要涉及到现有群落相对丰度的变化[28]。众多相关研究均显示不同的结果,认为水温、pH、DO、硅酸盐、氮、磷和COD或这些因子的不同组合等是影响浮游细菌分布的主要环境因素[29-31]。研究结果的差异除了与环境因子变化幅度不同有关,还可能是不同水体浮游细菌群落由于受到固有的种间关系影响,而形成了复杂群落的结构异质性[32]。本研究中各池塘优势菌群在组成上有较高的相似性,但优势菌群在分布和相对丰度变动上有较大差异 (图2)。而张皓[15]的调查却发现不同水样间优势菌在组成及丰度上均存在明显差异。这可能与采样频次、养殖密度和水环境状况等不同有关。调查发现在养殖前期、中期,优势菌群主要由营光合自养的GpIIa、红杆菌属及化能异养的芽殖杆菌属、微小杆菌属等有益菌株组成。后期条件致病菌不动杆菌属、黄杆菌属、假单胞菌属和气单胞菌属含量增加 (相对丰度最高值分别达18.2%、6.2%、11.2%和39.5%),从RDA分析结果可以看出,这些菌群主要与PO4-P和Sul呈正相关,与GpIIa、红杆菌属和芽殖杆菌属这些益生菌呈负相关,表明后期由于水体营养盐含量增加、水质恶化导致条件致病菌大量繁殖,有益菌生长严重受限。张皓[15]的研究也表明高氨氮、高有机物条件下,假单胞菌和黄杆菌属条件致病菌丰度增加。可见,当水环境状况变差,条件致病菌的发生率会增加,极易引发病害。另外,本研究发现随着ALK的增加,有益菌含量增加。ALK可缓存水体酸碱度,其变动改变了微生物酶的活性和膜的通透性,从而影响其生长状况。郭远涛[33]研究也表明随着ALK的消耗,细菌硝化作用减弱,氨氮去除率不再升高。本次调查各池塘ALK变动范围介于54.2~186.2 mg·L−1,此范围内ALK的增加可促进有益菌的生长。
3.2 凡纳滨对虾养殖池塘浮游细菌群落多样性特征
本次调查H'指数变动范围介于2.48~5.06,各池塘间H'指数差异不显著,这与杨淑芳[14]的调查结果近似。根据物种生态位保守假说[34],即物种在进化过程中生态位是保守的,亲缘关系越相近的物种其生态位越相似。不同物种会通过占有资源生态位的分化实现共存[35],PD值越小亲缘关系越相近,更趋向相似的生态位,池塘A浮游细菌物种丰富度、H'值和PD值均大于池塘C (P<0.05或P<0.01),因此相较池塘A和C的浮游细菌群落呈现更低的物种共存和系统发育多样性特征。
根据NMDS分析,各池浮游细菌群落在凡纳滨对虾养殖初期差异较大,至中、后期差异减小,在统计学上各池塘浮游细菌群落结构的变动差异并不显著。Fan等[36]对不同养殖品种的池塘浮游细菌群落的研究发现,池塘中养殖品种的不同是形成不同浮游细菌群落的主要因素,本研究是同一养殖场的凡纳滨对虾养殖池塘,未呈现显著的群落结构变动。虽然各池塘浮游细菌群落整体上差异不显著,但LEfSe分析仍识别到18个生物标记,这些类群显示出各池塘浮游细菌在各分类单元的差异菌群。如远洋杆属仅在后期出现,Sphingomonadaceae则仅在前期出现,这些菌群的分布可能受到水体营养及有机质状况的影响。
稳定的微生物环境对维持对虾的健康生长至关重要,一般认为多样性是反映群落稳定性的重要指标[37],高多样性指数一般表征更加稳定的群落特征[38]。本研究H'多样性与环境因子RDA分析表明,养殖初期和后期的样方,H'值较低 (均值分别为3.08和3.50,P<0.01);养殖中期的样方,H'值较高 (均值为4.30,P<0.01,图7),这与胡晓娟等[39]的研究结果近似。养殖初期,由于水体浮游细菌群落刚刚构建,H'值较低,随着营养元素的注入,H'值升高,至后期水体呈严重富营养化状态,有机物升高,H'值降低,条件致病菌增多,群落稳定性降低。Yang等[16]的研究也表明,水体富营养化会破坏浮游细菌群落的稳定性。根据中度干扰假说[40],中等程度的干扰使多样性维持在较高水平,可以容纳更多的物种迁入和定居;本研究符合中度干扰假说这一现象。由此可见,凡纳滨对虾养殖后期是水质调控的关键节点,此时应加强人为干预,对水质进行合理调控,如抗应激、杀灭有害菌藻、底质改良、培育有益微生物群落,重新构建生态系统。
-
![]()
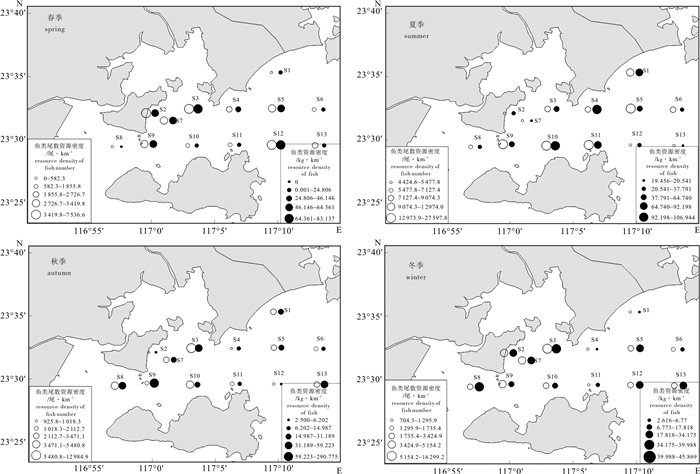
图 2 柘林湾海域鱼类各水层、食性和适温性种类数组成
Figure 2. Number of fish species of different water course, food habit and thermophile habit in Zhelin Bay
![]()
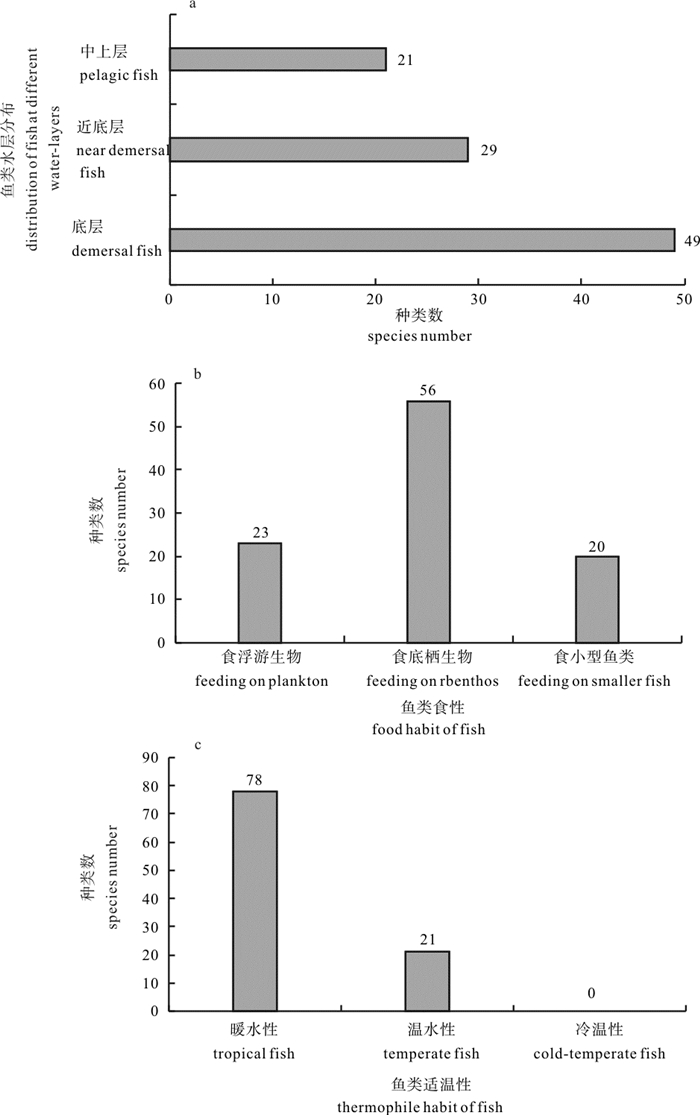
图 3 鱼类群落空间生态类型种数随季节变动情况
Figure 3. Change of number of ecological species of fish in four seasons
![]()
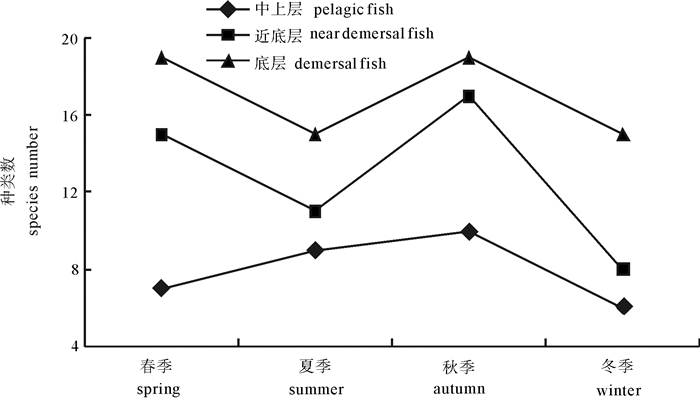
图 4 鱼类适温性类型种数随季节变动情况
Figure 4. Change of number of thermophile of fish in four seasons
![]()
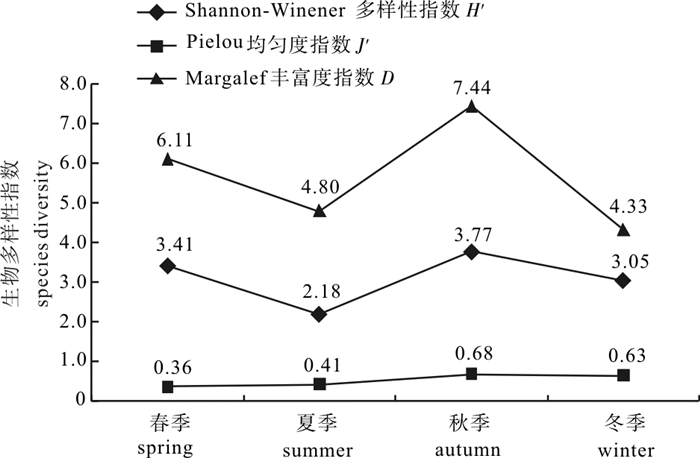
图 7 南海柘林湾鱼类生物多样性指数季节变动
Figure 7. Diversity indices of fishery species in Zhelin Bay of the South China Sea
![]()
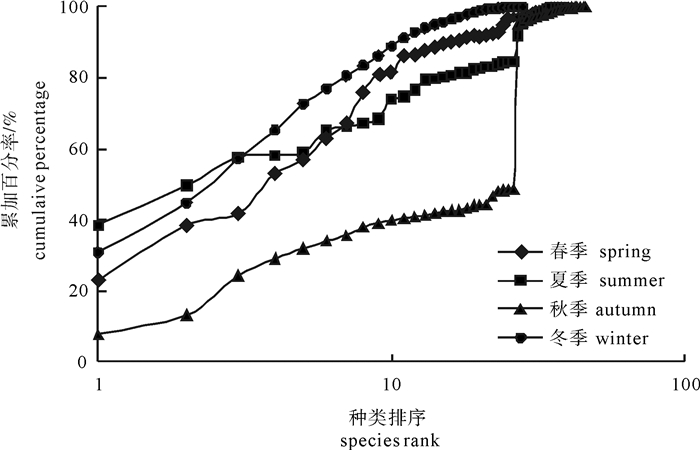
图 8 南海柘林湾鱼类生物量优势度曲线
Figure 8. K-dominance curves by weight in Zhelin Bay of the South China Sea
![]()
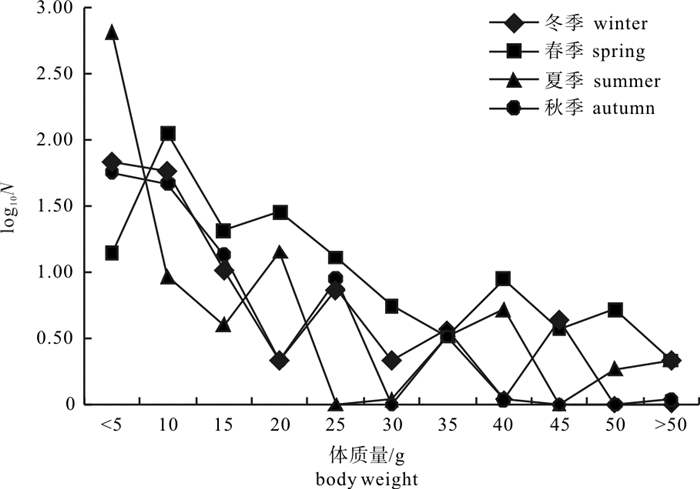
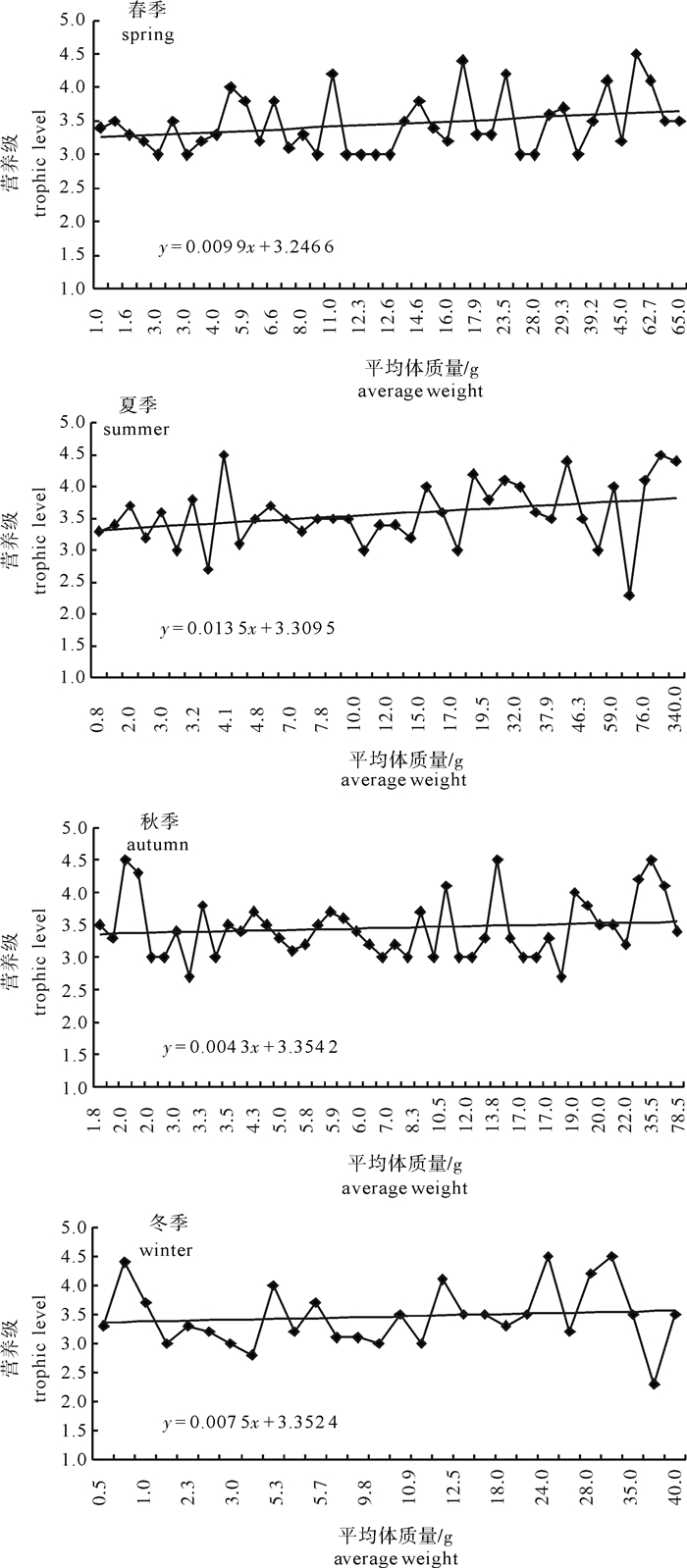
图 9 柘林湾海域4个季节鱼类营养级-平均体质量关系曲线
Figure 9. Relationship curves of trophic level-average body weight for fish in four seasons in Zhelin Bay
表 1 柘林湾海域周年各季度月质量百分比前5位鱼类
Table 1 Top five fishes in terms of biomass percentages in Zhelin Bay
春季spring % 夏季summer % 秋季autumn % 冬季winter % 拟矛尾 
22.17 鹿斑鲾Leiognathus ruconius 38.70 花鲈Lateolabrax japonicus 21.22 拟矛尾 30.12 皮氏叫姑鱼Johnius belengeri 15.92 杜氏棱鳀Thrissa dussumieri 11.32 龙头鱼Harpodon nehereus 4.95 龙头鱼Harpodon nehereus 14.77 龙头鱼Harpodon nehereus 11.59 白姑鱼Argyrosomus argentatus 8.00 矛尾 3.44 皮氏叫姑鱼Johnius belengeri 12.54 白姑鱼Argyrosomus argentatus 8.82 四指马鲅Eleutheronema tetradactylum 7.04 拟矛尾 2.52 钝孔 8.20 花 
5.47 带鱼Trichiurus haumela 6.33 皮氏叫姑鱼Johnius belengeri 2.21 带鱼Trichiurus haumela 7.58  下载: 导出CSV
下载: 导出CSV
表 2 柘林湾海域周年各季度月数量百分比前5位鱼类
Table 2 Top five fishes in terms of abundance percentages in Zhelin Bay
春季spring % 夏季summer % 秋季autumn % 冬季winter % 拟矛尾 41.87 鹿斑鲾Leiognathus ruconius 54.06 矛尾 34.00 拟矛尾 29.47 皮氏叫姑鱼Johnius belengeri 12.52 杜氏棱鳀Thrissa dussumieri 24.60 拟矛尾 15.84 灰康吉鳗Conger cinereus 27.51 鳓Ilisha elongata 8.20 白姑鱼Argyrosomus argentatus 8.51 龙头鱼Harpodon nehereus 6.62 矛尾 13.56 龙头鱼Harpodon nehereus 4.17 尖吻小公鱼Anchoviella heteroloba 2.59 皮氏叫姑鱼Johnius belengeri 5.44 钝孔 6.09 红狼牙 4.17 静鲾Leiognathus insidiator 1.56 孔 5.20 皮氏叫姑鱼Johnius belengeri 4.52
下载: 导出CSV
表 3 南海柘林湾海域各季节鱼类营养级
Table 3 Trophic level of fish of Zhelin Bay in the South China Sea in different seasons
春季spring 夏季summer 鱼种
species营养级
trophic level鱼种
species营养级
trophic level斑 3.0 中华小沙丁鱼Sardinella nymphaea 2.7 花 3.0 尖吻小公鱼Stolephorus heteroloba 3.3 鳓Ilisha elongata 3.8 赤鼻棱鳀Thrissa kammalensis 3.3 印度鳓Ilisha indica 3.8 杜氏棱鳀Thrissa dussumieri 3.7 康氏小公鱼Anchoviella commerson 3.3 多齿蛇鲻Saurida tumbil 4.4 赤鼻棱鳀Thrissa kammalensis 3.3 尖尾鳗Uroconger lepturus 3.5 七丝鲚Coilia grayi 3.3 海鳗Muraenesox cinereus 4.1 多齿蛇鲻Saurida tumbil 4.4 裸鳍虫鳗Muraenichthys gymnopterus 4.0 龙头鱼Harpodon nehereus 3.5 艾氏蛇鳗Ophichthus evermanni 4.5 黑尾前肛鳗Dysomma melanttrum 4.0 四指马鲅Eleutheronema tetradactylum 4.4 鳗鲇Plotosus anguillaris 3.5 六指马鲅Polynemus sextarius 3.7 六带石斑鱼Epinephelus sexfasciatus 3.8 六带石斑鱼Epinephelus sexfasciatus 3.8 多鳞 
3.4 网纹石斑鱼Epinephelus cholorstigma 4.0 杜氏叫姑鱼Johnius dussumieri 4.1 短尾大眼鲷Priacanthus macracanthus 4.1 皮氏叫姑鱼Johnius belengeri 3.3 斑鳍天竺鱼Apogonichthys carinatus 3.5 红牙Otolithes rubber 3.6 四线天竺鲷Apogon quadrifasciatus 3.5 勒氏短须石首鱼Umbrina russelli 4.1 中线天竺鲷Apogon kiensis 3.5 白姑鱼Argyrosomus argentatus 4.5 沟鲹Atropus atropus 3.6 浅色黄姑鱼Nibea chui 3.4 丽叶鲹Caranx kalla 3.8 棘头梅童鱼Collichthys lucidus 3.7 蓝圆鲹Decapterus maruadsi 3.4 短棘鲾Leiognathus equulus 3.0 竹筴鱼Trachurus japonicus 3.4 短吻鲾Leiognathus brevirostris 3.0 白姑鱼Argyrosomus argentatus 4.5 黄斑鲾Leiognathus bindus 3.3 短吻鲾Leiognathus brevirostris 3.0 胡椒鲷Plectorhynchus pictus 3.0 静鲾Leiognathus insidiator 3.2 李氏 
3.2 鹿斑鲾Leiognathus ruconius 3.0 小带鱼Eupleurogrammus muticus 4.2 日本金线鱼Nemipteras japonicus 3.6 带鱼Trichiurus haumela 4.2 吕宋绯鲤Upeneus luzonius 3.4 斑纹舌 3.0 李氏 3.2 长丝 3.0 黄斑蓝子鱼Siganus oramin 2.3 美岸栉 3.0 带鱼Trichiurus haumela 4.2 拟矛尾 3.1 印度无齿鲳Ariomma indica 3.6 红狼牙 3.5 刺鲳Psenopsis anomala 4.0 孔 3.5 犬牙细棘 3.0 褐菖鲉Sebastiscus marmoratus 3.2 拟矛尾 3.1 褐斑栉鳞鳎Aseraggodes kobensis 3.5 矛尾 3.5 娥眉条鳎Zebrias quagga 3.5 圆鳞斑鲆Pseudorhombus levisquamis 3.5 半滑舌鳎Cynoglossoides semilaevis 3.2 纤羊舌鲆Arnoglossus tenuis 3.5 大鳞舌鳎Cynoglossus macrolepidotus 3.2 棕斑腹刺鲀Gastrophysus spadiceus 3.0 横纹东方鲀Fugu oblongus 3.2 红鳍东方鲀Fugu rubripes 3.0 网纹东方鲀Fugu reticularis 3.0 秋季autumn 冬季winter 鱼种
species营养级
trophic level鱼种
species营养级
trophic level裘氏小沙丁鱼Sardinella jussieu 2.7 康氏小公鱼Stolephorus commerson 3.3 中华小沙丁鱼Sardinella nymphaea 2.7 赤鼻棱鳀Thrissa kammalensis 3.3 康氏小公鱼Stolephorus commerson 3.3 杜氏棱鳀Thrissa dussumieri 3.7 赤鼻棱鳀Thrissa kammalensis 3.3 凤鲚Coilia mystus 3.0 黄吻棱鳀Thrissa vitirostris 3.7 龙头鱼Harpodon nehereus 3.5 黄鲫Setipinna taty 3.2 灰康吉鳗Conger cinereus 4.4 凤鲚Coilia mystus 3.0 黑尾前肛鳗Dysomma melanttrum 4.0 七丝鲚Coilia grayi 3.3 前�骨鲻Osteomugil ophuyseni 2.3 龙头鱼Harpodon nehereus 3.5 杜氏叫姑鱼Johnius dussumieri 4.1 尖尾鳗Uroconger lepturus 3.5 皮氏叫姑鱼Johnius belengeri 3.3 海鳗Muraenesox cinereus 4.1 白姑鱼Argyrosomus argentatus 4.5 大头蚓鳗Moringua macrocephalus(Bleeker) 4.0 斑鳍白姑鱼Argyrosomus pawak 4.5 六指马鲅Polynemus sextarius 3.7 棘头梅童鱼Collichthys lucidus 3.7 花鲈Lateolabrax japonicus 3.4 鹿斑鲾Leiognathus ruconius 3.0 六带石斑鱼Epinephelus sexfasciatus 3.8 少鳞服 
3.0 中线天竺鲷Apogon kiensis 3.5 李氏 3.2 多鳞 3.4 带鱼Trichiurus haumela 4.2 丽叶鲹Caranx kalla 3.8 睛斑阿胡 2.8 布氏鲳鲹Trachinotus blochii 3.7 斜带栉眼 3.5 杜氏叫姑鱼Johnius dussumieri 4.1 拟矛尾 3.1 皮氏叫姑鱼Johnius belengeri 3.3 六丝矛尾 3.1 白姑鱼Argyrosomus argentatus 4.5 矛尾 3.0 斑鳍白姑鱼Argyrosomus pawak 4.5 红狼牙 3.5 大头白姑鱼Argyrosomus macrocephalus 4.5 孔 3.5 短吻鲾Leiognathus brevirostris 3.0 神农吻 3.5 黄斑鲾Leiognathus bindus 3.3 钝孔 3.5 鹿斑鲾Leiognathus ruconius 3.0 半滑舌鳎Cynoglossoides semilaevis 3.2 短棘银鲈Gerres lucidus 3.5 横纹东方鲀Fugu oblongus 3.2 细鳞 
3.0 赤刀鱼Cepola schlegeli 3.4 六带拟鲈Parapercis sexfasciata 3.4 李氏 3.2 沙带鱼Lepturacanthus savala 4.3 带鱼Trichiurus haumela 4.2 巴布亚沟 3.0 南方沟 3.0 犬牙细棘 3.0 拟矛尾 3.1 矛尾 3.0 孔 3.0 褐菖鲉Sebastiscus marmoratus 3.2 大鳞鳞鲬Onigocia macrolepis 3.5 鲬Platycephalus indicus 3.6 卵鳎Solea ovata 3.5 大鳞舌鳎Cynoglossus macrolepidotus 3.2 日本细鳞鲀Stephanolepis japonicus 3.0 注:鱼类营养级数据来自FishBase网站。
Note:The trophic data are taken from FishBase website.
下载: 导出CSV
-
[1] 于海婷, 丁月晏, 线薇微, 等.荣成湾渔业资源群落结构季节变化特征[J].海洋湖沼通报, 2013(2):67-75. http://www.cnki.com.cn/Article/CJFDTOTAL-HYFB201302011.htm [2] 乔永民, 黄长江.柘林湾表层沉积物重金属分布及污染初步评价[J].海洋科学, 2006, 30(3):41-45. http://www.oalib.com/paper/4669901 [3] 马继波, 董巧香, 黄长江.粤东大规模海水增养殖区柘林湾浮游细菌的时空分布[J].生态学报, 2007, 27(2):477-485. http://www.oalib.com/references/19444378 [4] 史赟荣, 晁敏, 全为民, 等.2010年春季长江口鱼类群落空间分布特征[J].中国水产科学, 2011, 18(5):1141-1151. http://industry.wanfangdata.com.cn/dl/Detail/Periodical?id=... [5] 金显仕, 邓景耀.莱州湾渔业资源群落结构和生物多样性的变化[J].生物多样性, 2000, 8(1):65-72. http://www.wenkuxiazai.com/doc/887242e96294dd88d0d26ba1-3.html [6] 王雪辉, 邱永松, 杜飞雁, 等.北部湾鱼类群落格局及其与环境因子的关系[J].水产学报, 2010, 34(10):1579-1586. http://www.docin.com/p-1015987319.html [7] 陈嘉欣, 区又君, 廖银萍, 等.北部湾雷州近海春、夏季鱼类群落结构初探[J].生态科学, 2006, 25(4):354-358. http://www.wenkuxiazai.com/doc/03a94ecdb9f3f90f76c61bc7.html [8] 陈国宝, 李永振, 陈新军.南海主要珊瑚礁水域的鱼类物种多样性研究[J].生物多样性, 2007, 15(4):373-381. http://industry.wanfangdata.com.cn/dl/Detail/Periodical?id=... [9] 陈国宝, 李永振.南海主要珊瑚礁科鱼类的组成与分布[J].南方水产, 2005, 1(3):18-25. http://www.cqvip.com/QK/60642A/200503/27478445.html [10] 汪振华, 赵静, 王凯, 等.马鞍列岛岩礁生境鱼类群落结构时空格局[J].生态学报, 2013, 33(19):6218-6226. http://www.cnki.com.cn/Article/CJFDTOTAL-STXB201319034.htm [11] 汪振华, 王凯, 赵静, 等.枸杞岛潮下带沙地生境鱼类群落结构和季节变化[J].应用生态学报, 2011, 22(5):1332-1342. http://www.wenkuxiazai.com/doc/5e23f036192e45361066f51c.html [12] 马彩华, 游奎, 李凤岐, 等.南海鱼类生物多样性与区系分布[J].中国海洋大学学报(自然科学版), 2006, 36(4):665-670. http://www.doc88.com/p-990978065223.html [13] 乔延龙, 林昭进, 邱永松.北部湾秋、冬季渔业生物群落结构特征的变化[J].广西师范大学学报(自然科学版), 2008, 26(1):100-104. http://edu.wanfangdata.com.cn/Periodical/Detail/gxsfdxxb200801025 [14] CLARKE K R.Comparisons of dominance curves[J].J Exp Mar Bio Ecol, 1990, 138(1/2):143-157. http://d.wanfangdata.com.cn/NSTLQK_10.1016-0022-0981(90)90181-B...
[15] 任一平, 徐宾铎, 叶振江, 等.青岛近海春、秋季渔业资源群落结构特征的初步研究[J].中国海洋大学学报(自然科学版), 2005, 35(5):792-798. http://www.wenkuxiazai.com/doc/4233473702020740be1e9b42.html [16] 杜建国, 陈彬, 卢振彬, 等.泉州湾海域鱼类多样性及营养级变化[J].生物多样性, 2010, 18(4):420-427. http://med.wanfangdata.com.cn/Paper/Detail/PeriodicalPaper... [17] 王迎宾, 郑基, 郑献之, 等.舟山渔场禁渔线以外海域单拖网鱼类群落结构变动分析[J].南方水产科学, 2012, 8(1):8-15. http://www.schinafish.cn/CN/abstract/abstract8934.shtml [18] 单秀娟, 孙鹏飞, 金显仕, 等.黄海典型断面渔业资源结构的季节变化[J].水产学报, 2013, 37(3):425-435. http://www.cqvip.com/QK/90183X/201303/45244549.html [19] 单秀娟, 金显仕.长江口近海春季鱼类群落结构的多样性研究[J].海洋与湖沼, 2011, 42(1):32-40. doi: 10.11693/hyhz201101005005 [20] AOYAMA T.The South China Sea fisheries(demersal resources)[M].Rome:UNDP and FAO, 1973:59-67.
[21] 王跃中, 袁蔚文.南海北部底拖网渔业资源的数量变动[J].南方水产, 2008, 4(2):26-33. http://www.oalib.com/references/19444090 [22] 程济生, 俞连福.黄、东海冬季底层鱼类群落结构及多样性变化[J].水产学报, 2004, 28(1):29-34. http://www.oalib.com/paper/5092642 [23] TANG Q S.Effects of long-term physical and biological perturbations on the contemporary biomass yields of the Yellow Sea ecosystem[M]//Large marine ecosystems:stress, mitigation, and sustainability.Washington, D.C.:AAAS Press, 1993:79-93.
[24] JIN X S.Seasonal changes of the demersal fish community of the Yellow Sea[J].Asian Fish Sci, 1995, 8:177-190. http://www.doc88.com/p-7357701577598.html
[25] JIN X, XU B, TANG Q.Fish assemblage structure in the East China Sea and southern Yellow Sea during autumn and spring[J].J Fish Biol, 2003, 62(5):1194-1205. doi: 10.1046/j.1095-8649.2003.00116.x
[26] JIN X S, TANG Q S.Changes in fish species diversity and dominant species composition in the Yellow Sea[J].Fish Res, 1996, 26(3/4):337-352. https://www.sciencedirect.com/science/article/pii/016578369500422X
[27] 纪炜炜, 李圣法, 陈雪忠.鱼类营养级在海洋生态系统研究中的应用[J].中国水产科学, 2010, 17(4):878-887. http://www.wenkuxiazai.com/doc/02a7071902020740be1e9b98.html -
期刊类型引用(4)
1. 侯明勇,张燕萍,余建芳,阙祥尧,吴子君,徐先栋,宋超,章海鑫. 环鄱阳湖地区池塘水体细菌群落结构特征及构建机制. 江西农业大学学报. 2025(01): 178-192 .  百度学术
百度学术
2. 张露,朱建强,李滔,章叶飞,万肖,刘章勇,杨军. 微生物菌剂对克氏原螯虾稻田养殖水体浮游植物群落结构和水质的影响. 水产学杂志. 2024(04): 91-99 . 百度学术
3. 余鑫瑞,吴桃,王浩,闫海明,韩剑钧,管卫兵. 凡纳滨对虾盐碱水养殖池塘浮游细菌群落变化特征. 中国水产科学. 2024(08): 940-953 . 百度学术
4. 顾颖,伏光辉,姚永琪,梁宝贵,叶仁智,王超,卢璐,孙苗苗. 海州湾细菌群落结构多样性及环境因子分析. 生命科学研究. 2023(06): 512-520+527 . 百度学术
其他类型引用(0)











计量
- 文章访问数: 3980
- HTML全文浏览量: 1367
- PDF下载量: 781
- 被引次数: 4
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
