
Microstructure and texture characteristics analysis of grass carp (Ctenopharyngodon idellus) muscle at different stages of crispness
-
摘要:
脆肉鲩作为广东特色水产品之一,其养殖中常出现“不脆”、“半脆”现象,严重制约其产业发展。为探索可鉴定脆度的指标,文章结合生产(饲喂120 d蚕豆+30 d饲料),每30 d采集1次肌肉,检测其感官脆度、肌纤维结构、质构特性的变化。随着脆化时间增加,蚕豆组草鱼感官脆度不断增加(P < 0.05),肌纤维直径逐渐减小、密度逐渐增大(P < 0.05);60 d后蚕豆组草鱼质构特性(硬度、弹性、咀嚼性、内聚性、胶黏性)均显著高于对照组(P < 0.05)。转投30 d饲料对感官脆度、肌纤维结构、质构特性均无显著影响(P>0.05)。相关性分析表明,硬度、弹性、咀嚼性、胶黏性与感官脆度的相关性更高(R2 >0.90,P < 0.05)。结果表明,硬度、弹性、咀嚼性、胶黏性可用于脆肉鲩的脆度评价,转投30 d饲料不影响其脆度。
Abstract:Crisp grass carp (Ctenopharyngodon idellus) is one of the characteristic fishery products in Guangdong Province, but its "no crispness" and "semi crispness" have seriously restricted the industry′s development. In order to explore the evaluation method of crispness, we combined a production process (120 d faba bean + 30 d artificial feed). The muscle samples were collected once every 30 d, and the changes of sensory crispness, texture characteristics and muscle fiber structure were detected. The results demonstrate that sensory crispness of grass carp increased continuously with the increase of production time (before 120 d) in faba bean group, while the diameter of muscle fiber decreased and the density increased gradually (P < 0.05);after 60 d, the texture characteristics (hardness, springiness, chewiness, cohesiveness and gumminess) in faba bean group were significantly better than those of the control (P < 0.05). Feeding artificial diet of 30 d did not affect the sensory crispness, texture characteristics and muscle fiber structure (P>0.05). The correlation analysis shows that the correlation among hardness, springiness, chewiness, gumminess and sensory crispness was much higher (R2>0.90, P < 0.05). It is concluded that the hardness, springiness, chewiness and gumminess can be used to evaluate the crispness, and 30-day feed did not affect the crispness.
-
Keywords:
- crisp grass carp /
- grass carp /
- crispness /
- muscle fiber /
- microstructure /
- texture characteristics
-
Dkkl1 (Dickkopf-like 1) 是Dickkopf (DKK) 基因家族中的一员,属于分泌型糖蛋白,最初是在非洲爪蟾 (Xenopus laevis) 中被鉴定为胚胎头部诱导剂和Wingless (Wnt) 拮抗剂[1]。已有研究证明Dkkl1可以在脊椎动物的多种细胞类型中抑制Wnt诱导的β-catenin的稳定性[2-3]。在小鼠 (Mus musculus) 中,Dkkl1由单个基因编码,其表达首先在2细胞期和4细胞期中检测到[4],在胚胎第15天时,Dkkl1基因在发育中的骨骼和眼睛中表达,但在性腺原基中不表达。然而在成年小鼠中,Dkkl1基因的表达只限于睾丸,Dkkl1 mRNA在发育中的精母细胞中丰富表达,首先在发育中的顶体中表达,然后在成熟精子的顶体中积累[5]。不仅如此,Dkkl1还是一种N-糖基化蛋白,在精母细胞成熟过程中参与精子合成[6],它标志着发育中的精母细胞和精子细胞,通过靶向敲除Dkkl1基因,小鼠胚胎正常发育,形成可育后代[7]。但是,Dkkl1基因的缺失会导致精子体外受精能力严重受损,而这种作用在体内受精中似乎得到了某种因素的补偿。此外,Dkkl1基因还与人 (Homo sapiens) 的弱精子症[8]和不育症[9]密切相关。上述研究表明,Dkkl1基因在哺乳动物睾丸发育和精子发生过程中扮演着十分重要的角色,但是目前关于Dkkl1基因在龟鳖类动物中的研究还鲜有报道。
爬行纲在物种进化过程中具有特殊的进化位置,是第一批真正摆脱水环境依赖而登陆的脊椎动物,也是羊膜动物中最早的一种,既能进化为哺乳类,又能演化为鸟类,在动物学研究中占有举足轻重的地位[10]。中华鳖 (Pelodiscus sinensis) 隶属于爬行纲、龟鳖目、鳖科、中华鳖属,是我国淡水特种养殖中的重要品种,因其独特的营养价值[11]和药用价值[12]而深受市场青睐,直接经济价值超过百亿。随着年龄的增长,中华鳖睾丸发育呈现出不同的形态特征,大致可分为4个时期:精原细胞期、精母细胞期、精子细胞期和精子期。1冬龄雄性中华鳖精巢中的细胞分散,曲细精管内主要由精原细胞组成;2冬龄曲细精管中的生精上皮除了有精原细胞外,初级精母细胞也逐渐分化形成;3冬龄精巢已具备各种类型的精细胞,曲细精管腔内出现大量精子[13-14]。中华鳖的睾丸发育和精子发生受多基因的调控,例如:Li等[15]发现中华鳖中Hmgb2基因的表达呈现睾丸特异性,主要存在于圆形精子细胞和精子中,证实其在中华鳖的精巢发育和精子生成过程中具有重要功能;Lei等[16]发现Spats1是一个雄性表达特异的基因,其主要存在于中华鳖的初级精母细胞、次级精母细胞和精子中,与中华鳖的精子发生和释放密切相关;Zhou等[17]使用RNA干扰技术成功敲除Amh基因,证明其在促进中华鳖的睾丸发育和精子发生方面具有必要和充分的作用。然而,除了关键基因之外,外源性激素也可以在一定程度上调控睾丸发育和精子发生[18-19],且在不同发育时期发挥着不同的作用[20]。例如:意大利壁蜥 (Podarcis sicula) 在高浓度17β-雌二醇 (E2) 和低浓度甲基睾酮 (MT) 中会阻滞精子发生[21],而在低浓度E2和高浓度MT中可诱导精子发生的恢复[22]。锯盖鱼 (Centropomus undecimalis) 在15和30 mg·kg–1的MT下可刺激睾丸的发育和生长并加速精子发生[23];产后15~30 d的大鼠暴露于E2下会延迟精子发生[24]。然而,近年有关Dkkl1基因的研究十分匮乏,且现有的研究还远不足以阐明中华鳖精子发生的背后机制。因此本研究克隆了中华鳖Dkkl1基因的cDNA序列,对其序列特征进行详细分析,探索其在不同体组织和不同时期性腺中的表达模式,以及其对外源性激素 [E2和17α-甲基睾酮 (17α-MT)] 处理的响应,为深入探讨Dkkl1基因在中华鳖睾丸发育和精子发生中的功能研究提供基础数据。
1. 材料与方法
1.1 实验材料的收集
本研究所用中华鳖采购自广东省惠州市财兴实业有限公司,选取1、2、3冬龄健康中华鳖,麻醉后放血。取3冬龄中华鳖的心脏、肝脏、脾脏、肾脏、脑、肌肉、卵巢和1、2、3冬龄的精巢组织于无酶冻存管中,并将其迅速放入液氮中速冻,于−80 ℃超低温冰箱保存,每种组织均取3个生物重复。
1.2 总RNA提取及cDNA第一链合成
使用Trizol法提取中华鳖各组织的总RNA,提取RNA的浓度和质量由NanoQTM核酸检测仪和1% (w) 琼脂糖凝胶电泳检测。使用HiScript® III RT SuperMix for qPCR (+gDNA wiper) 反转录试剂盒 (Vazyme,中国) 去除基因组DNA污染,合成cDNA第一链。
1.3 中华鳖Dkkl1 cDNA的克隆
参考NCBI GenBank公布的中华鳖Dkkl1基因序列信息 (XM_014572465) 设计引物Dkkl1-F/Dkkl1-R (表1),以3冬龄中华鳖精巢cDNA为模板,将Dkkl1基因的开放阅读框 (Open reading frame, ORF) 区域进行扩增,并使用1%琼脂糖凝胶电泳对PCR产物进行检测。目的片段使用Gel Extraction Kit (Omega,中国) 胶回收试剂盒进行回收,连接至pMD19-T (TaKaRa,中国) 载体上,将连接产物转化至DH5α感受态细胞 (TaKaRa,中国) 中。最后,从转化后的细胞中筛选出阳性克隆,并送至广州天一辉远基因科技有限公司进行测序。
表 1 中华鳖 Dkkl1 cDNA 克隆和表达分析的引物Table 1 Primers used for cDNA cloning and expression analysis of P. sinensis Dkkl1名称
Name序列 (5'—3')
Sequences (5'–3')用途
PurposeDkkl1-F GGGACAGGGAAGGGAAAC cDNA克隆 Dkkl1-R GCAAGAAAGACCAAGGAGTAGG Dkkl1-qF ATGGCTAGCAGCCTGTGTCT 实时荧光定量和半定量分析 Dkkl1-qR GACCTGGCAAAGAGATGGAG EF1αF ACTCGTCCAACTGACAAGCCTC 内参基因 EF1αR CACGGCGAACATCTTTCACAG 1.4 Dkkl1基因的生物信息学分析
使用ExPASy (https://web.expasy.org/translate/) 在线网站预测开放阅读框[25],采用DNAMAN软件进行氨基酸序列的翻译和同源性比较[26]、ClustalW (BioEdit) 软件进行氨基酸序列的多重比对[27-28],通过ProtParam分析Dkkl1蛋白的理化性质[29]和SOPMA分析Dkkl1蛋白二级结构[30],利用SWISS-MODLE同源建模构建Dkkl1蛋白质三级结构[31]。
1.5 组织表达分析
运用Primer软件设计实时荧光定量PCR (RT-qPCR) 所需引物Dkkl1-qF/Dkkl1-qR,同时选择中华鳖的Ef1α基因作为内参基因[32] (表1)。以中华鳖不同组织的cDNA作为模板,利用Applied Biosystems StepOnePlus Real-Time PCR Systems (Applied Biosystems,新加坡)对其进行实时荧光定量PCR分析。反应体系为20 μL,其中2×iTaq Universal SYBR Green (BIO-RAD,美国) 10 μL,引物Dkkl1-qF/Dkkl1-qR各0.5 μL,cDNA模板2 μL,双蒸水 (ddH2O) 7 μL。半定量分析使用与RT-qPCR相同的特异性引物 (Dkkl1-qF/Dkkl1-qR),采用各组织的cDNA作为模板进行普通PCR (RT-PCR) 扩增。扩增产物通过1%琼脂糖凝胶分离并进行拍照 (GenoSens2200,中国)。
1.6 成体雄性中华鳖外源性激素处理
选取体表无伤、活跃的3冬龄雄性中华鳖33只,其中30只平均分为2组 (E2处理组和17α-MT处理组),从腿部肌肉注射E2和17α-MT,质量浓度为50 μg·μL−1,剂量为10 mg·kg−1[16];另外3只注射等剂量的溶剂作为空白对照组,分别于激素处理后第6、第12、第24、第48小时和第7天随机采集3只雄性性腺,置于液氮中进行总RNA提取和实时荧光定量PCR分析。
1.7 数据分析
使用2−ΔCt法于第6、第12、第24、第48小时计算目的基因的相对表达量[15],每个实验至少独立重复3次,所有数据均以“平均值±标准误(
$\bar x\pm s_{\bar x} $ )”表示。采用SPSS 20.0软件进行单因素方差分析,P<0.05表示具有显著性差异。2. 结果
2.1 中华鳖Dkkl1基因克隆结果及其编码蛋白氨基酸序列分析
克隆获得的中华鳖Dkkl1 cDNA序列长为823 bp,其中3' 非编码区 (UTR) 长67 bp,5' UTR长90 bp,ORF长666 bp,共编码222个氨基酸(图1)。中华鳖Dkkl1蛋白的相对分子质量为24.924 77 kD,理论等电点为10.8,不稳定系数为58.71,脂溶系数为84.91,总平均亲水系数为−0.418,表明Dkkl1蛋白是一种稳定性较差、亲水性较强的碱性蛋白质。其二级结构主要由α-螺旋、β-折叠、无规则卷曲和延伸链组成,其中α-螺旋占33.78%,β-折叠占5.86%,无规则卷曲占46.85%,延伸链占13.51%,三级结构主要以α-螺旋和无规则卷曲为主。氨基酸序列同源性比对结果显示:其与中华草龟(Chinemys reevesii)的相似性较高 (81%),与棱皮龟 (Dermochelys coriacea)的相似性较低 (70%),与绿海龟(Chelonia mydas)、三趾箱龟(Terrapene carolina triunguis)、巴西红耳龟(Trachemys scripta elegans)、西部锦龟(Chinemys picta bellii)的同源性分别为80%、79%、78%、78% (图2)。
![]() 图 1 中华鳖 Dkkl1 基因核苷酸序列及其推导的氨基酸序列Fig. 1 Nucleotide of P. sinensis Dkkl1 gene and its encoded protein sequence
图 1 中华鳖 Dkkl1 基因核苷酸序列及其推导的氨基酸序列Fig. 1 Nucleotide of P. sinensis Dkkl1 gene and its encoded protein sequence![]() 图 2 中华鳖与其他物种的 Dkkl1 氨基酸序列同源相似性比对注:所有的氨基酸序列来自于NCBI数据库:巴西红耳龟,XM_034791724.1;绿海龟,XM_037888484.1;三趾箱龟,XM_026659251.1;中华草龟,XM_039510142.1;西部锦龟,XM_024113109.1;棱皮龟,XM_038381465.2。黑色阴影为氨基酸序列比对相同的位点,灰色阴影为相近位点。Fig. 2 Homologous comparison of Dkkl1 amino acid sequence of P. sinensis with other speciesNote: All amino acid sequences come from NCBI database: T. scripta elegans, XM_034791724.1; C. mydas, XM_037888484.1; T. carolinatriunguis, XM_026659251.1; C. reevesiis, XM_039510142.1; C. picta bellii, XM_024113109.1; D. coriacea, XM_038381465.2. The black and gray shadows represent the same and similar sites, respectively.
图 2 中华鳖与其他物种的 Dkkl1 氨基酸序列同源相似性比对注:所有的氨基酸序列来自于NCBI数据库:巴西红耳龟,XM_034791724.1;绿海龟,XM_037888484.1;三趾箱龟,XM_026659251.1;中华草龟,XM_039510142.1;西部锦龟,XM_024113109.1;棱皮龟,XM_038381465.2。黑色阴影为氨基酸序列比对相同的位点,灰色阴影为相近位点。Fig. 2 Homologous comparison of Dkkl1 amino acid sequence of P. sinensis with other speciesNote: All amino acid sequences come from NCBI database: T. scripta elegans, XM_034791724.1; C. mydas, XM_037888484.1; T. carolinatriunguis, XM_026659251.1; C. reevesiis, XM_039510142.1; C. picta bellii, XM_024113109.1; D. coriacea, XM_038381465.2. The black and gray shadows represent the same and similar sites, respectively.2.2 中华鳖Dkkl1基因的组织表达
RT-qPCR结果显示,中华鳖Dkkl1 mRNA在3冬龄成体精巢中极显著性高表达 (P<0.001),而在心脏、肝脏、脾脏、肾脏、脑、肌肉和卵巢中几乎不表达 (图3-a)。RT-PCR结果显示,该基因仅在3冬龄中华鳖成体精巢组织中检测到目的条带,而在其他体组织中均未检测到 (图3-b)。
![]() 图 3 中华鳖 Dkkl1 基因在 3 冬龄成体组织中的表达情况注:***. P<0.001;M. DL2000 Marker。Fig. 3 Expression of Dkkl1 gene in tissues of 3-winter-age adult P. sinensisNote: ***. P<0.001; M. DL2000 Marker.
图 3 中华鳖 Dkkl1 基因在 3 冬龄成体组织中的表达情况注:***. P<0.001;M. DL2000 Marker。Fig. 3 Expression of Dkkl1 gene in tissues of 3-winter-age adult P. sinensisNote: ***. P<0.001; M. DL2000 Marker.2.3 中华鳖不同发育时期精巢中Dkkl1基因的表达变化
通过RT-qPCR检测中华鳖Dkkl1基因在不同发育时期精巢中的表达变化。结果显示,随着中华鳖年龄的增长,其精巢中Dkkl1基因的表达量逐渐上升,并在3冬龄时达到顶峰。此外,Dkkl1基因在2冬龄和3冬龄精巢中的表达量极显著高于1冬龄 (P<0.001),而2冬龄和3冬龄精巢之间的表达差异不显著 (P>0.05) (图4)。
![]() 图 4 中华鳖 Dkkl1 基因在不同发育时期精巢中的表达情况注:**. P<0.01表示差异极显著。Fig. 4 Expression of P. sinensis Dkkl1 gene in different developmental periods of testisNote: **. P<0.01 represents very significant difference.
图 4 中华鳖 Dkkl1 基因在不同发育时期精巢中的表达情况注:**. P<0.01表示差异极显著。Fig. 4 Expression of P. sinensis Dkkl1 gene in different developmental periods of testisNote: **. P<0.01 represents very significant difference.2.4 E2和17α-MT处理对中华鳖精巢Dkkl1基因表达的影响
对雄性中华鳖进行E2和17α-MT 激素处理后,其精巢中Dkkl1基因的表达量显著降低 (P<0.05)。E2处理后,精巢中Dkkl1基因的表达量持续下降,至第48小时几乎为0,但是在7 d后得到部分恢复;17α-MT处理后,精巢中Dkkl1基因的表达量同样受到显著性抑制 (P<0.05),在第48小时达到最低值,但在第7天显著上升 (P<0.05) (图5)。
![]() 图 5 中华鳖精巢中 Dkkl1 基因对 17β-雌二醇和17α- 甲基睾酮处理后的表达响应注:不同小写字母表示组间存在显著性差异(P<0.05)。Fig. 5 Expression response of Dkkl1 gene in testis of P. sinensis to E2 and 17α-MT treatmentNote: Different lowercase letters represent significant differences between groups (P<0.05).
图 5 中华鳖精巢中 Dkkl1 基因对 17β-雌二醇和17α- 甲基睾酮处理后的表达响应注:不同小写字母表示组间存在显著性差异(P<0.05)。Fig. 5 Expression response of Dkkl1 gene in testis of P. sinensis to E2 and 17α-MT treatmentNote: Different lowercase letters represent significant differences between groups (P<0.05).3. 讨论
本实验通过克隆得到中华鳖Dkkl1基因的cDNA序列,序列长823 bp,共编码222个氨基酸,氨基酸序列比对和NJ进化树结果表明其与爬行类亲缘关系最近。RT-qPCR和RT-PCR结果显示Dkkl1 mRNA在精巢组织中高表达且极显著高于卵巢等其他组织 (P<0.001),表明Dkkl1基因表达在中华鳖中具有性别二态性和组织特异性。这与在小鼠和人类中发现的两性差异表达模式类似。Dkkl1基因的组织分布研究表明,Dkkl1 mRNA仅在睾丸中表达丰富,而在附睾和其他组织中几乎不表达。此外,人类基因芯片结果显示Dkkl1基因在成年男性睾丸中的杂交信号强度是胎儿睾丸的405.56倍,对人体多个组织的RT-PCR分析表明,Dkkl1 mRNA仅在睾丸中表达,Western blot分析也表明Dkkl1基因主要在人睾丸中表达[5]。综上所述,推测Dkkl1基因在中华鳖睾丸形成或功能维持上可能具有重要的调控作用。
同时,本实验发现,随着中华鳖年龄的增长,其精巢中Dkkl1基因的表达量逐渐上升,并在3冬龄时达到顶峰,Dkkl1基因在中华鳖成体睾丸中的表达与其睾丸发育和精子发生的过程大致吻合。1冬龄中华鳖的睾丸主要由精原细胞组成;2冬龄时逐渐分化为初级精母细胞和次级精母细胞,并且在生精管中观察到少量精子;3冬龄时已经完全性成熟,睾丸中产生大量成熟精子[33]。Dkkl1基因这种特殊的表达模式与睾丸发育成熟程度呈正相关,并且很可能参与了精子发生过程。因此,在生产实践中通过Dkkl1基因的相对表达量可预估中华鳖睾丸的发育成熟程度,这可为选育高繁殖力和优秀的雄性亲本提供新的生物指标。
精子发生是雌激素与雄激素共同调节的结果[34]。E2是一种天然的雌激素[35],17α-MT是一种具有雄激素效应的内分泌干扰物[36],这两种物质已被广泛用于与睾丸发育和精子发生有关的功能研究[37-38]。本实验通过向雄性中华鳖注射外源性激素E2和17α-MT后,发现两者均可显著抑制Dkkl1基因的表达。相比于17α-MT,E2对Dkkl1基因的抑制作用更强,在注射12 h后,其表达量几乎为零。有研究表明,外源性雌激素可通过影响雌激素受体途径诱导精子发生障碍,并对精子的质量和受精潜力产生负面影响[39],低剂量的E2可延迟精子产生,而高剂量的E2则会完全抑制精子产生[40]。Dkkl1基因在精子发生过程中被定位在成熟的精子或精子的顶体中[4],同时还发现Dkkl1基因在小鼠胎盘发育和精子发生过程中表达,并且具有促进精子穿透透明带的能力,将其敲除后,小鼠体外受精能力显著降低[6]。因此,外源性激素E2处理可能是抑制Dkkl1基因表达的关键因素。雌、雄激素通过不同的受体途经影响精子发生,而雄激素大多是通过雄激素特异性受体AR介导的[41]。本实验中,通过注射外源性激素17α-MT导致Dkkl1基因的表达受到显著抑制。Dakhova等[42]发现Dkkl1基因是睾酮产生的负调控因子,敲除Dkkl1基因导致类固醇生成酶基因Cyp11a和Cyp17表达量增加,使睾酮的合成局部上调。而注射外源性激素17α-MT会导致生物体内睾酮水平瞬间升高[43],从而可能抑制中华鳖Dkkl1基因的转录,以响应体内睾酮水平的升高。
4. 结论
本实验通过克隆得到中华鳖Dkkl1基因的cDNA片段,对其进行生物学信息分析,发现中华鳖在进化关系上与爬行类动物相近。运用RT-PCR和RT-qPCR对Dkkl1基因在雌、雄中华鳖各组织和精巢不同发育时期的表达差异进行了分析,确定Dkkl1基因在3冬龄成体精巢中显著高表达,并且随着年龄的增长其表达量逐渐升高。通过注射外源性激素E2和17α-MT,发现其可显著抑制Dkkl1基因在精巢中的表达。由以上结果可以推论,Dkkl1基因在中华鳖睾丸发育和成熟过程中起着重要作用,可能与精子发生密切相关,且可受外源性激素调控,这为今后龟鳖类动物睾丸发育及精子发生机制的研究拓展了新思路。
-
![]()
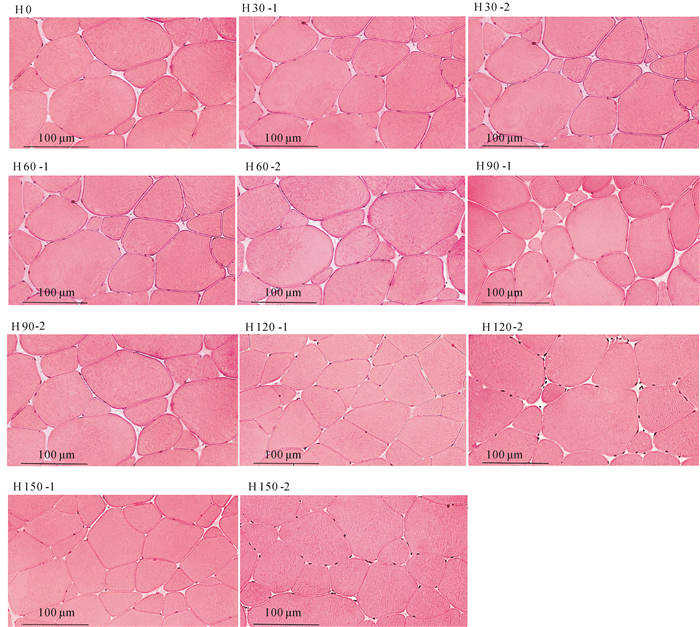
图 1 草鱼肌肉的横切面显微结构(×400)
H0、H30、H60、H90、H120、H150分别代表第0、第30、第60、第90、第120、第150天的肌肉横切;-1代表蚕豆组,-2代表对照组。
Figure 1. Transverse section microstructure of grass carp muscle (×400)
H0, H30, H60, H90, H120 and H150 denote transverse section microstructure on the 0th, 30th, 60th, 90th, 120th, 150th day, respectively; "-1" stands for faba bean group; "-2" stands for control group.
![]()
图 2 草鱼肌肉的纵切显微结构(×400)
Z0、Z30、Z60、Z90、Z120、Z150分别代表第0、第30、第60、第90、第120、第150天的肌肉纵切;-1代表蚕豆组,-2代表对照组。
Figure 2. Longitudinal section microstructure of grass carp muscle (×400)
Z 0, Z30, Z60, Z90, Z120 and Z150 denote longitudinal section microstructure on the 0th, 30th, 60th, 90th, 120th, 150th day, respectively; "-1" stands for faba bean group; "-2" stands for control group.
![]()
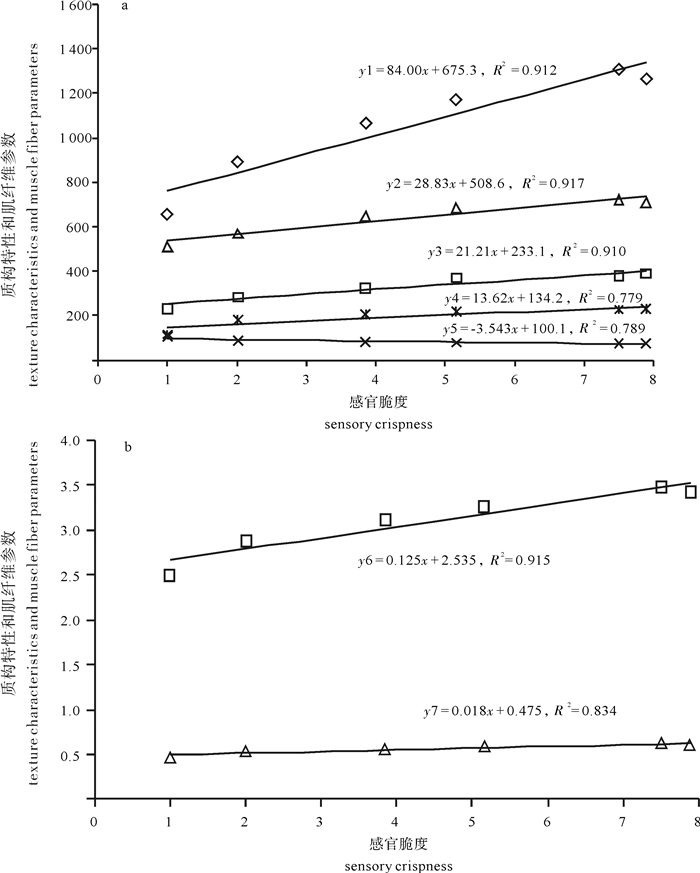
图 3 感官脆度和质构特性及肌纤维参数的线性关系
y1.咀嚼性;y2.硬度;y3.胶黏性;y4.肌纤维密度;y5.肌纤维直径;y6.弹性; y7.内聚性
Figure 3. Linear correlation between sensory crispness and parameters of texture characteristics and muscle fiber parameters
y1. chewiness; y2. hardness; y3. gumminess; y4. muscle fiber density; y5. muscle fiber diameter; y6. springiness; y7. cohesiveness
表 1 不同脆化阶段草鱼肌纤维直径和密度
Table 1 Diameter and density of muscle fiber of grass carp at different crisping periods
项目item 脆化时间/d crisping time 0 30 60 90 120 直径/μm diameter 对照组 104.58±6.54a 105.26±4.00a 103.59±6.65a 103.42±3.12a 104.98±7.21a 蚕豆组 88.20±2.87b 82.12±3.50c 78.01±4.87d 75.17±1.92d 密度/No.·mm-2 density 对照组 116±11e 117±7e 114±8e 116±5e 117±7e 蚕豆组 181±11d 204±10c 219±13b 231±14a 注:同一行上标字母不同表示差异性显著(P<0.05),后表同此。
Note:Different superscript letters in the same row indicate significant difference between treatments (P<0.05).The same case in the following table. 下载: 导出CSV
下载: 导出CSV
表 2 不同脆化阶段草鱼肌肉TPA指标值
Table 2 TPA index values of grass carp muscle at different crisping periods
项目item 脆化时间/d crisping time 0 30 60 90 120 硬度/g hardness 对照组 508.52±66.89c 504.47±59.54c 510.54±66.43c 506.38±67.53c 511.36±59.22c 蚕豆组 569.60±67.02bc 646.00±45.95b 685.28±73.02ab 710.73±60.79a 弹性springiness 对照组 2.50±0.19c 2.34±0.13c 2.67±0.24c 2.55±0.33c 2.47±0.17c 蚕豆组 2.88±0.06b 3.11±0.04ab 3.26±0.31a 3.43±0.42a 咀嚼性/g chewiness 对照组 655.14±83.66c 644.86±54.24d 654.25±84.76d 658.11±53.34d 662.63±64.12d 蚕豆组 890.32±21.89c 1 067.11±214.14b 1 170.85±127.10ab 1 263.61±122.06a 内聚性cohesiveness 对照组 0.46±0.04c 0.46±0.02c 0.45±0.04c 0.48±0.03c 0.46±0.09c 蚕豆组 0.53±0.02b 0.57±0.02ab 0.59±0.05a 0.60±0.06a
下载: 导出CSV
-
[1] 谢骏, 郁二蒙, 王广军, 等."脆肉鲩"的养殖关键技术及其产业化开发[J].科学养鱼, 2009(5):41-42. http://www.wenkuxiazai.com/doc/c807c8c86137ee06eff91813.html [2] 刘庆营.草鱼脆化养殖新技术[J].农村实用技术, 2008(6):53. http://www.wenkuxiazai.com/doc/5a621127a32d7375a4178050.html [3] 李忠铭, 冷向军, 李小勤, 等.脆化草鱼生长性能、肌肉品质、血清生化指标和消化酶活性分析[J].江苏农业科学, 2012, 40(3):186-189. http://www.docin.com/p-527044645.html [4] 程纯明.微变革, 脆肉鲩要从13斤进入15斤的新历程, 凭借的是什么?[J].当代水产, 2015, 40(9):48-49. http://industry.wanfangdata.com.cn/yj/Magazine?magazineId=nlsc&... [5] 刘怡.《海洋与渔业》走进"中国脆肉鲩之乡"中山东升镇[J].海洋与渔业, 2014(3):30-33. http://www.cnki.com.cn/Article/CJFDTotal-HYUY201403024.htm [6] 林婉玲, 杨贤庆, 李来好, 等.脆肉鲩质构与感官评价的相关性研究[J].现代食品科技, 2013, 29(1):1-7. http://www.cqvip.com/QK/92448A/201301/44546154.html [7] YU E M, LIU B H, WANG G J, et al. Molecular cloning of type I collagen cDNA and nutritional regulation of type I collagen mRNA expression in grass carp[J]. J Anim Physiol Anim Nutr (Berl), 2014, 98(4):755-765. doi: 10.1111/jpn.2014.98.issue-4
[8] 肖调义, 刘建波, 陈清华, 等.脆肉鲩肌肉营养特性分析[J].淡水渔业, 2004, 34(3):28-30. http://www.cnki.com.cn/Article/CJFDTOTAL-DSYY200403008.htm [9] 朱志伟, 李汴生, 阮征, 等.脆肉鲩鱼肉与普通鲩鱼鱼肉理化特性比较研究[J].现代食品科技, 2008, 24(2):109-112, 119. http://www.doc88.com/p-6015982449717.html [10] 甘承露, 郭姗姗, 荣建华, 等.脆肉鲩肌肉主要营养成分的分析[J].营养学报, 2010, 32(5):513-515. http://www.cqvip.com/QK/95640X/201005/35713240.html [11] 伦峰, 冷向军, 李小勤, 等.投饲蚕豆对草鱼生长和肉质影响的初步研究[J].淡水渔业, 2008, 38(3):73-76. http://www.cqvip.com/qk/93836X/200803/27676191.html [12] 陈清华, 肖调义, 吴松山, 等.脆肉鲩肌肉游离氨基酸初步分析[J].水利渔业, 2004, 24(6):8-10. http://www.wenkuxiazai.com/doc/63a284f1f8c75fbfc77db2db.html [13] 谭乾开, 黎华寿.脆化草鱼(Ctenopharyngodon idellus C.et V)的病理生理生态学[J].生态学报, 2006, 26(8):2749-2756. http://www.cnki.com.cn/Article/CJFDTOTAL-STXB200608044.htm [14] 谭乾开, 黎华寿.食物与水环境因子对草鱼(Ctenopharyngodon idellus C.et V)脆化过程的影响[J].生态学报, 2006, 26(7):2409-2415. http://www.cnki.com.cn/Article/CJFDTOTAL-STXB200607045.htm [15] YANG S L, LI L H, QI B, et al.Quality evaluation of crisp grass carp (Ctenopharyngodon idellus C. et V) based on instrumental texture analysis and cluster analysis[J].Food Anal Methods, 2015, 8(8):2107-2114. doi: 10.1007/s12161-015-0101-2
[16] 郁二蒙, 谢骏, 卢炳国, 等.脆肉鲩与普通草鱼肌肉显微结构观察[J].南方农业学报, 2014, 45(4):671-675. http://www.cqvip.com/QK/90177A/201404/50011561.html [17] LIN W L, ZENG Q X, ZHU Z W.Different changes in mastication between crisp grass carp (Ctenopharyngodon idellus C.et V) and grass carp (Ctenopharyngodon idellus) after heating:the relationship between texture and ultrastructure in muscle tissue[J].Food Res Int, 2009, 42(2):271-278. doi: 10.1016/j.foodres.2008.11.005
[18] 李道友. 草鱼脆化养殖过程中的肌肉特性及脆性表征[D]. 武汉: 华中农业大学, 2011: 39-48. http://cdmd.cnki.com.cn/Article/CDMD-10504-1011405247.htm [19] 关磊, 朱瑞俊, 李小勤, 等.普通草鱼与脆化草鱼的肌肉特性比较[J].上海海洋大学学报, 2011, 20(5):748-753. http://www.wenkuxiazai.com/doc/636bdd3516fc700abb68fc8c.html [20] 林婉玲, 关熔, 曾庆孝, 等.影响脆肉鲩鱼背肌质构特性的因素[J].华南理工大学学报(自然科学版), 2009, 37(4):134-137. http://industry.wanfangdata.com.cn/dl/Detail/Periodical?id=... [21] 刘建波, 陈开健, 佘曙明, 等.脆肉鲩肌肉超微结构分析[J].水利渔业, 2005, 25(3):65-67. http://www.cnki.com.cn/Article/CJFDTOTAL-SCAN200503031.htm [22] JOHNSTON I A, ALDERSON R, SANDHAM C, et al. Muscle fibre density in relation to the colour and texture of smoked Atlantic salmon (Salmo salar L.)[J].Aquaculture, 2000, 189(3/4):335-349. https://www.sciencedirect.com/science/article/pii/S0308814603005417
[23] GAO J C, PIGOTT G M, REINE B.Gel forming additive effects on properties of thermally induced minced fish gel[J]. J Food Sci, 1999, 64(3):414-417. doi: 10.1111/jfds.1999.64.issue-3
[24] URESTI R M, TELLEZ L J, RAMIREZ J A, et al.Use of dairy proteins and microbial transglutaminase to obtain low-salt fish products from filleting waste from silver carp(Hypophthalmichthys molitrix)[J].Food Chem, 2004, 86(2):257-262. doi: 10.1016/j.foodchem.2003.09.033
[25] HUIDOBRO F R, MIGUEL E, BLAZQUEZ B, et al. A comparison between two methods (Warner-Bratzler and texture profile analysis) for testing either raw meat or cooked meat[J].Meat Sci, 2005, 69(3):527-536. doi: 10.1016/j.meatsci.2004.09.008
[26] 伍芳芳, 林婉玲, 李来好, 等.草鱼脆化过程中肌肉品质变化[J].南方水产科学, 2014, 10(4):70-77. http://www.schinafish.cn/CN/abstract/abstract9140.shtml [27] AYALA M D, ALBORS O L, BLANCO A, et al. Structural and ultrastructural changes on muscle tissue of sea bass, Dicentrarchus labrax L., after cooking and freezing[J].Aquaculture, 2005, 250(1/2):215-231.
[28] HURLING R, RODELL J B, HUNT H D. Fiber diameter and fish texture[J].J Texture Stud, 1996, 27(6):679-685. doi: 10.1111/jts.1996.27.issue-6
[29] BUGEON J, LEFEVRE F, FAUCONNEAU B. Fillet texture and muscle structure in brown trout (Salmo trutta) subjected to long-term exercise[J].Aquacult Res, 2003, 34(14):1287-1295. doi: 10.1046/j.1365-2109.2003.00938.x
[30] PERIAGO M J, AYALA M D, LOPEZ A O, et al. Muscle cellularity and flesh quality of wild and farmed sea bass, Dicentrarchus labrax L[J].Aquaculture, 2005, 249(1):175-188.
[31] JOHNSTON I A, MANTHRI S, BICKERDIKE R, et al. Growth performance, muscle structure and flesh quality in out-of-season Atlantic salmon (Salmo salar) smolts reared under two different photoperiod regimes[J].Aquaculture, 2004, 237(1):281-300. https://www.researchgate.net/profile/Tom_Hansen3/2?page=2&sorting=...
[32] JOHNSTON I A, LI X, VIEIRA V A, et al. Muscle and flesh quality traits in wild and farmed Atlantic salmon[J].Aquaculture, 2006, 256(1):323-336. https://www.sciencedirect.com/science/article/pii/S0044848606001517
-
期刊类型引用(0)
其他类型引用(1)









计量
- 文章访问数: 4448
- HTML全文浏览量: 1293
- PDF下载量: 812
- 被引次数: 1
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
