
Antioxidant stability and functional properties of enzymatic hydrolysates from purple back flying squid (Symplectoteuthis oualaniensis)
-
摘要:
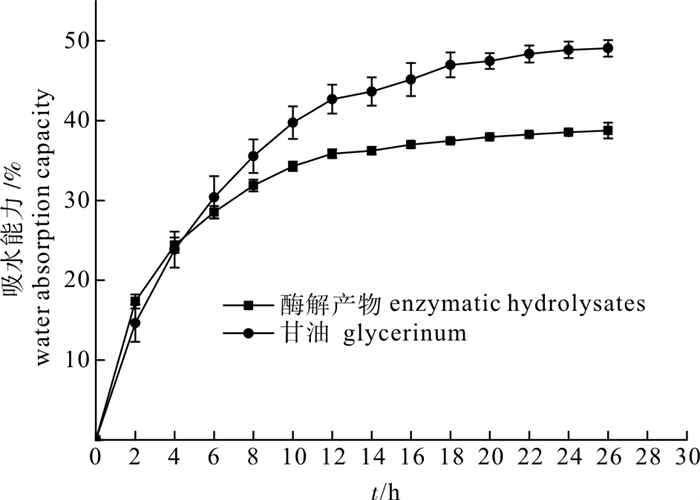
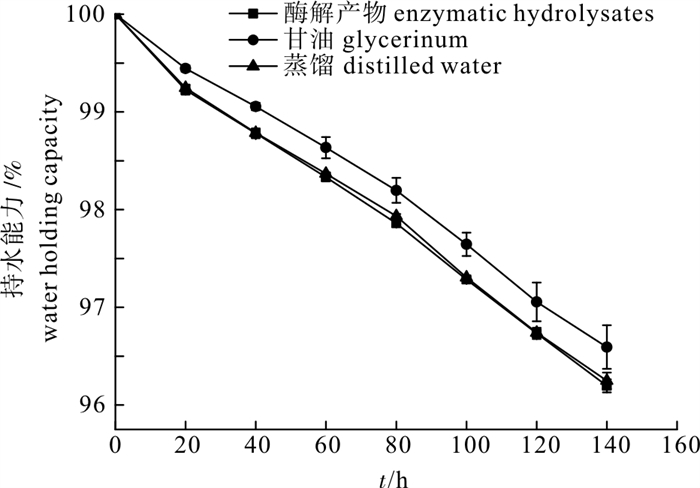
以鸢乌贼(Symplectoteuthis oualaniensis)胴体为原料,采用木瓜蛋白酶进行酶解,比较了酶解液中不同分子量组分的抗氧化特性,探讨了温度、pH和光照对其抗氧化活性的影响,以及其功能特性(包括乳化性和乳化稳定性、起泡性和泡沫稳定性、吸水性、持水性和吸油性等)。结果表明,鸢乌贼酶解产物具有一定的耐热性,在酸性偏中性的环境中能较好地保持其抗氧化活性,日光照射会降低其抗氧化能力。此外,酶解产物还具有较强的乳化性、起泡性、吸水吸油能力等功能特性。
Abstract:Purple back flying squids (Symplectoteuthis oualaniensis) were hydrolyzed by papain. We compared the antioxidant properties of enzymatic hydrolysates with different molecular weights components and investigated the effects of temperature, pH and light on the antioxidant stability and functional properties (emulsifying properties, emulsion stability, foaming properties, foaming stability, water absorption capacity, water holding capacity and oil absorption capacity) of enzymatic hydrolysates. The results show that enzymatic hydrolysates had certain heat resistance; the activity was maintained well in acidic or neutral environment, and light could reduce its antioxidant capacity. In addition, enzymatic hydrolysates had good emulsifying properties, foaming properties as well as water and oil absorption capacity.
-
Keywords:
- purple back flying squid /
- enzymatic hydrolysates /
- antioxidant /
- stability
-
发酵是用于食品保存的一种常见加工方式,在改善食品营养和功能特性方面起着重要作用[1],其中发酵鱼作为一种传统的加工水产制品,主要依靠微生物在厌氧环境下和相对稳定的温度中进行长期发酵而成[2]。发酵鱼制品不仅营养丰富,还具有新鲜鱼所没有的独特风味和口感,在中国、日本、泰国等亚州国家食用广泛,具有一定的市场需求。
发酵鳜鱼(Siniperca chuatsi)又称桶鲜鱼、臭桂鱼等,是安徽徽州菜的代表菜之一,传统发酵鳜鱼加工方式是将新鲜或冷冻的鳜鱼经腌制后密封并在室温环境下发酵7~15 d[3],蛋白质水解释放氨基酸、脂解产生游离脂肪酸等均是发酵成熟过程中重要的生化反应[4]。经腌制发酵后的发酵鳜鱼具有“闻起来臭,吃起来香”的特点[5],深受消费者的喜爱。近年来,发酵鳜鱼在营养品质、理化性质、菌群结构等多个方面逐渐引起了学者们的注意,其中贺永玲等[6]对不同发酵阶段的臭鳜鱼进行营养成分探究;王伟等[7]比较了发酵鳜鱼与新鲜鳜鱼的营养品质。而目前市售的发酵鳜鱼主要以小作坊加工方式为主,受传统自然发酵生产条件的限制且技术水平和集约化程度不高,难以保证产品品质的稳定性及安全性;同时,不同厂家加工工艺的不同使其生产的发酵鳜鱼产品品质有所差异。为此,本文选取两家安徽企业生产的发酵鳜鱼作为研究对象,通过检测营养成分和安全性指标以评估市售发酵鳜鱼的营养特点和食用安全性,为其加工技术的改进提供理论依据。
1. 材料与方法
1.1 材料与试剂
新鲜鳜鱼、1号发酵鳜鱼和2号发酵鳜鱼均购于广州市华润万家超市。
石油醚,氯化钾,三氯乙酸,乙二胺四乙酸 (EDTA),2-硫代巴比妥酸,冰乙酸,亚硝酸盐,对氨基苯磺酸,氯化钠等试剂均为分析纯,购自上海摩楷生物科技有限公司;37种脂肪酸标准品,8种生物胺标准品,丹磺酰氯,乙腈等均为色谱纯,购自广州领驭生物科技有限公司。
1.2 方法
1.2.1 样品处理
分别取新鲜鳜鱼和发酵鳜鱼背部肌肉,用流动水冲洗干净、绞碎后混合均匀,−18 ℃冻藏备用。
1.2.2 基本营养成分的测定
水分测定参照GB 5009.3—2016《食品中水分的测定》;灰分测定参照GB 5009.4—2016《食品中灰分的测定》;粗蛋白测定参照GB 5009.5—2016《食品中蛋白质的测定》;粗脂肪测定参照GB 5009.6—2016《食品中脂肪的测定》。
1.2.3 氨基酸测定
16种氨基酸测定参照GB 5009.124—2016《食品中氨基酸的测定》;色氨酸测定参照GB/T 15400—2018《饲料中色氨酸的测定》。
1.2.4 脂肪酸测定
参考韩迎雪等[8]的方法稍作修改。准确称取2.0 g绞碎鱼肉于均质杯中,加入氯仿-甲醇 (V∶V=2∶1) 混合溶液 (含有0.01%2,6-二叔丁基-4-甲基苯酚),均质1 min,用氯仿-甲醇混合溶液定容至30 mL,4 ℃静置1 h后用滤纸过滤,向滤液中加入0.2倍0.85%生理盐水,振荡混匀后,4 000 r·min−1离心15 min,弃去上层液体,下层脂质溶液氮吹吹干,加入2 mL 14%三氟化硼-甲醇溶液,60 ℃水浴30 min后取出,冷却至室温,加入1 mL正己烷和1 mL蒸馏水,充分振荡混匀,静置分层后吸取上层有机层,过0.22 μm有机滤膜,待GC-MS上机分析。利用NIST 0.5谱库数据库及MS图库中标准谱图进行脂肪酸分析,并采用面积归一化法计算脂肪酸的相对含量。
1.2.5 矿物质与重金属元素测定
称取0.500 g样品于聚四氟乙烯消解管内,加入10 mL浓硝酸,经微波消解后冷却至室温,加超纯水定容至50 mL。采用电感耦合等离子体质谱法测定样品中矿物质及重金属元素[9]。
1.2.6 硫代巴比妥酸值 (TBA) 测定
参照陈丽丽等[10]和魏颖等[11]的方法稍作修改。准确称取10.00 g绞碎鱼肉于均质杯中,加入50 mL 7.5%三氯乙酸混合溶液 (含0.1% EDTA),均质1 min,在4 ℃冰箱内静置30 min后用滤纸过滤2遍。移取5 mL滤液于比色管内,加入5 mL 0.02 mol·L−1TBA溶液,沸水浴保持40 min后取出,冷却至室温,加入5 mL氯仿,充分振荡混匀,静置待分层后取上层清液,测定532 nm和600 nm处吸光值,并计算TBA值。
1.2.7 挥发性盐基氮 (T-VBN) 测定
参照张婷等[12]的方法稍作修改。准确称取5.0 g绞碎鱼肉于均质杯中,加入45 mL 0.6 mol·L−1高氯酸溶液,均质2 min,4 ℃冰箱内静置40 min后,10 000 r·min−1离心10 min,取滤液,用自动凯式定氮仪进行分析。
1.2.8 亚硝酸盐测定
参照国标GB 5009.33—2016《食品中亚硝酸盐与硝酸盐的测定》。
1.2.9 生物胺测定
生物胺的提取与衍生参照GB 5009.208—2016《食品中生物胺的测定》,采用高效液相色谱法进行生物胺测定并制作生物胺标准曲线对样品中8种生物胺进行定量分析。
1.2.10 氨基酸营养价值评价方法
根据FAO/WHO 1973建议的氨基酸评分标准模式和全鸡蛋蛋白模式进行营养评价,蛋白质的氨基酸评分 (Amino acid score,AAS)、化学评分 (Chemical score,CS) 和必需氨基酸评分 (Essential amino acid index,EAAI) 按以下所列公式求得[13]:
$$ {\rm{AAS = }} \dfrac{{\text{所测样品中某种氨基酸}}}{{{\rm{FAO}}/{\rm{WHO}}{\text{评分标准模式中同种氨基酸含量}}}} $$ $${ {\rm{CS = }}\frac{\text{所测样品中某种氨基酸含量}}{\text{全鸡蛋蛋白质中同种氨基酸含量}}} $$ $$ {\rm{EAAI = }}\sqrt[n]{{\left( {\frac{{{\text{缬氨酸}}{{t}} \times {\text{苏氨酸}}{{t}} \times \cdots \times {\text{赖氨酸}}{{t}}}}{{{\text{缬氨酸}}{{s}} \times {\text{苏氨酸}}{{s}} \times \cdots \times {\text{赖氨酸}}{{s}}}}} \right)}} \times 100 $$ 式中n为参与计算的必需氨基酸数目;t为被考察的蛋白质中氨基酸质量分数 (mg·g−1);s为标准蛋白质中相对应的氨基酸质量分数 (mg·g−1)。
1.3 数据处理
采用Excel 2010、SPSS 17.0统计软件对获得的数据进行分析处理。
2. 结果与分析
2.1 发酵鳜鱼的营养组成分析
不同厂家发酵鳜鱼的加工工艺各有不同,如发酵时间、发酵温度和食盐添加量等。新鲜鳜鱼的水分质量分数为81.85%,1号发酵鳜鱼为77.67%,2号为79.34%,两款发酵鳜鱼的水分含量均有所下降,这与发酵过程添加一定量的食盐有关,食盐渗入鱼体后导致渗透压加强,部分水分流失,使得水分含量下降[7]。发酵鳜鱼干样中基本营养成分见表1,发酵鳜鱼中灰分含量均高于新鲜鳜鱼,其中1号发酵鳜鱼干样中灰分含量明显高于2号,这与食盐添加量不同有关;两款发酵鳜鱼中粗蛋白和粗脂肪含量均有一定差异,这可能与加工工艺、所选用鳜鱼的个体大小、肥瘦、生存环境 (产地) 等不同有关,如李燕等[14]发现3种不同品种鳜鱼干样中基本营养成分含量存在一定差异。
表 1 发酵鳜鱼肌肉中基本营养成分质量分数 (干基)Table 1. Nutrients contents in muscle of fermented mandarin fish (dry mass)% 营养成分
Nutrients新鲜鳜鱼
Fresh mandarin fish1号发酵鳜鱼
No.1 Fermented mandarin fish2号发酵鳜鱼
No.2 Fermented mandarin fish灰分 Ash 5.22±0.07a 12.83±0.30b 10.69±0.17c 粗蛋白 Crude protein 85.93±0.73a 73.24±1.41b 77.30±1.36c 粗脂肪 Crude fat 9.89±0.70a 13.44±0.24b 11.76±0.35c 注:同行中均值具有不同字母者为差异显著 (P<0.05);后表同此 Note: The values with different superscript letters in the same row are significantly different (P<0.05). The same case in the following tables 2.2 发酵鳜鱼肌肉中氨基酸分析
2.2.1 氨基酸组成分析
鱼肉蛋白质的初步水解主要受内源性组织蛋白酶作用,进而在微生物蛋白酶的影响下进一步降解为小分子肽和游离氨基酸[4]。从表2可看出,经发酵,鳜鱼肌肉中氨基酸总量均有所增加,说明促进了游离氨基酸的释放。必需氨基酸中赖氨酸、亮氨酸、苯丙氨酸和缬氨酸等含量显著增加,其中赖氨酸作为人体必需氨基酸在植物蛋白中相对缺乏[15],发酵鳜鱼中较高的赖氨酸含量可补充这一营养成分;而亮氨酸可促进骨骼肌的生长发育与蛋白质形成[16];非必需氨基酸中谷氨酸、天冬氨酸、丙氨酸和酪氨酸等含量显著增加,甘氨酸与脯氨酸含量无明显差异,其中谷氨酸、天冬氨酸、丙氨酸和甘氨酸均为重要的鲜味氨基酸,蛋白质中鲜味氨基酸的含量对鱼肉鲜美程度起到关键作用,发酵鳜鱼鲜味氨基酸总量的增加,有助于其鲜味的提升。有研究发现微生物对个别游离氨基酸的变化具有较大的影响[17],1号发酵鳜鱼中精氨酸含量显著高于2号,但从整体而言,两款发酵鳜鱼中氨基酸总含量及其组成的差异并不明显,这可能是由于鳜鱼肉组织蛋白酶对蛋白质水解起到主要作用,而微生物起到次要作用。两款发酵鳜鱼必需氨基酸 (EAA) /总氨基酸 (TAA) 分别为 (42.02±0.28)%、(42.17±0.14)%,略高于新鲜鳜鱼 (41.68±0.01)%;EAA/非必需氨基酸 (NEAA) 分别为 (85.29±1.07)%、(85.47±0.60)%,略高于新鲜鳜鱼 (84.09±0.18)%,均符合FDA/WHO的推荐要求 (FAO/WHO的理想模型中EAA/TAA约40 %,EAA/NAEE约60%[7])。
表 2 发酵鳜鱼肌肉中氨基酸含量 (湿基)Table 2. Amino acids content in muscle of fermented mandarin fish (wet mass)g·kg−1 氨基酸
Amino acid新鲜鳜鱼
Fresh mandarin fish1号发酵鳜鱼
No.1 Fermented mandarin fish2号发酵鳜鱼
No.2 Fermented mandarin fish苏氨酸* Thr 7.78±0.05a 8.21±0.04b 8.20±0.08b 缬氨酸* Val 7.81±0.01a 8.47±0.08b 8.51±0.05b 蛋氨酸* Met 5.43±0.02a 5.91±0.05b 5.89±0.01b 异亮氨酸* Ile 7.29±0.01a 7.84±0.04b 7.86±0.06b 亮氨酸* Leu 13.27±0.01a 14.34±0.04b 14.34±0.12b 苯丙氨酸* Phe 6.83±0.04a 7.44±0.09b 7.50±0.06b 赖氨酸* Lys 16.41±0.01a 17.18±0.05b 17.24±0.11b 色氨酸* Try 1.46±0.04a 1.63±0b 1.61±0.03b 组氨酸** His 3.50±0.05a 3.93±0.01b 3.95±0.04b 精氨酸** Arg 10.44±0.04a 10.8±0.03b 10.37±0.05a 天冬氨酸# Asp 16.83±0.09a 18.24±0.11b 18.23±0.16b 丝氨酸 Ser 6.85±0.04a 7.26±0.04b 7.22±0.08b 谷氨酸# Glu 26.38±0.03a 28.03±0.04b 28.05±0.25b 脯氨酸 Pro 4.32±0.11a 4.35±0.27a 4.40±0.08a 甘氨酸# Gly 8.34±0.10a 8.25±0.33a 8.24±0.27a 丙氨酸# Ala 10.10±0.06a 10.75±0.04b 10.76±0.16b 酪氨酸 Tyr 5.98±0.04a 6.38±0.07b 6.33±0.09b 氨基酸总量 TAA 158.98±0.50a 168.97±0.42b 168.65±1.62b 必需氨基酸含量 EAA 66.26±0.19a 71.00±0.30b 71.12±0.44b 半必需氨基酸含量 CEAA 13.93±0.08a 14.73±0.04b 14.31±0.08c 非必需氨基酸含量 NEAA 78.79±0.40a 83.25±0.69b 83.22±1.10b 鲜味氨基酸含量 DAA 61.65±0.28a 65.26±0.45b 65.27±0.83b 必需氨基酸/总氨基酸 EAA/TAA (%) 41.68±0.01 42.02±0.28 42.17±0.14 必需氨基酸/非必需氨基酸 EAA/NEAA (%) 84.09±0.18 85.29±1.07 85.47±0.60 注:本实验未检测胱氨酸;*. 必需氨基酸;**. 半必需氨基酸;#. 鲜味氨基酸 Note: No cystine was detected. *. Essential amino acids; **. Semiessential amino acid; #. Delicious amino acids 2.2.2 氨基酸营养价值评价
营养学中,氨基酸评分 (AAS) 与化学评分 (CS) 是对食物中蛋白质进行营养评价的重要指标,而必需氨基酸指数 (EAAI) 则可反映出样品中必需氨基酸含量与标准蛋白质的接近程度[18]。发酵鳜鱼肌肉AAS、CS、EAAI均高于新鲜鳜鱼,说明发酵后,鳜鱼的氨基酸营养价值有一定提升 (表3)。不同加工工艺对发酵鱼的氨基酸营养价值也有一定影响,这可能是由于两种发酵工艺使得蛋白质的降解程度不同。两款发酵鳜鱼AAS均超过1,符合FAO/WHO的建议标准,且2号发酵鳜鱼AAS均高于1号;同时2号发酵鳜鱼中苏氨酸、亮氨酸和赖氨酸CS更高,苯丙氨酸+酪氨酸CS接近1,且EAAI (97.07) 高于1号发酵鳜鱼EAAI (94.72),属于优质蛋白源[18],同时与鸡蛋蛋白氨基酸模式更为接近,其氨基酸营养价值更高。
表 3 发酵鳜鱼肌肉必需氨基酸组成评价Table 3. Evaluation of essential amino acids composition of fermented mandarin fish muscle必需氨基酸
Essential amino acidFAO/WHO评分模式
FAO score mode/(mg·g−1)鸡蛋蛋白
Egg protein/(mg·g−1)新鲜鳜鱼
Fresh mandarin fish1号发酵鳜鱼
No.1 Fermented mandarin fish2号发酵鳜鱼
No.2 Fermented mandarin fishAAS CS AAS CS AAS CS 苏氨酸 Thr 250 292 1.25 1.07 1.25 1.07 1.28 1.10 缬氨酸 Val 310 411 1.01 0.76 1.05 0.79 1.07 0.81 异亮氨酸 Ile 250 331 1.17 0.88 1.19 0.90 1.24 0.93 亮氨酸 Leu 440 534 1.21 1.00 1.24 1.02 1.27 1.05 苯丙氨酸+酪氨酸 Phe + Tyr 380 565 1.35 0.91 1.39 0.93 1.42 0.96 赖氨酸 Lys 340 441 1.93 1.49 1.93 1.49 1.98 1.53 色氨酸 Try 60 99 1.00 0.61 1.02 0.62 1.04 0.63 必需氨基酸指数 EAAI 92.45 94.72 97.07 注:AAS. 氨基酸评分;CS. 化学评分 Note: AAS. Amino acid score; CS. Chemical score 2.3 发酵鳜鱼肌肉中脂肪酸含量分析
脂解是发酵鱼制品在加工过程中重要的生化反应,脂质经脂肪酶和磷脂酶的作用,释放出游离脂肪酸,其中脂肪细胞和肌肉中的内源酶及细菌中分泌的酶均参与脂解作用[19]。发酵鳜鱼肌肉中不饱和脂肪酸含量增加而饱和脂肪酸含量有所下降,这表明发酵促进了饱和脂肪酸的降解,有助于不饱和脂肪酸的释放 (表4)。如Xu等[20]发现从传统发酵鱼 (酸鱼) 中分离出的菌株木糖葡萄球菌 (Staphylococcus xylosus) 135和酿酒酵母 (Saccharomyces cerevisiae) 31均具有较好的脂解作用,且主要释放的游离脂肪酸为多不饱和脂肪酸。棕榈酸 (C16:0)、硬脂酸 (C18:0)、油酸 (C18:1)、棕榈油酸 (C16:1)、DHA (C22:6)、花生四烯酸 (C20:4) 和亚油酸 (C18:2) 等是发酵鳜鱼中主要的游离脂肪酸,但受到pH、温度、环境湿度及发酵菌种等条件的多重影响,采用不同发酵工艺加工的发酵鳜鱼中游离脂肪酸含量常存在一定差异。2号发酵鳜鱼中饱和脂肪酸含量普遍低于1号,其中C16:0和C14:0显著低于1号,饱和脂肪酸的减少有助于降低三高疾病的发生几率[21]。不饱和脂肪酸中,1号的C16:1、C20:4、EPA和DHA等含量显著高于2号,其中EPA和DHA被认为是人和动物生长发育的必需脂肪酸[22],对高血压、哮喘和心血管疾病等病症具有一定帮助[23],且其EPA+DHA含量最高 [(15.58±0.25)%],具有较高的食用价值和保健作用。而2号发酵鳜鱼C18:2含量远高于1号,同时C18:1和C20:3等含量也较高,C18:1和C18:2可抑制小肠对胆固醇的吸收,促进肝脏内胆固醇的降解与排除,改变体内胆固醇的分布[24],两者含量的升高可能是由于其内源性脂肪酶活性较高,并优先释放亚油酸和油酸,既是脂肪酶对甘油三酯SN-3位置的特异性作用,又与磷脂酶对磷脂的活性有关[25]。
表 4 发酵鳜鱼肌肉中脂肪酸组成及相对含量Table 4. Fatty acids composition and relative content in muscle of fermented mandarin fish% 脂肪酸
Fatty acid新鲜鳜鱼
Fresh mandarin fish1号发酵鳜鱼
No.1 Fermented mandarin fish2号发酵鳜鱼
No.2 Fermented mandarin fish十一碳酸 (C11:0) Undecanoate acid 0.02±0.01a 0.07±0.03a 0.04±0.03a 十三碳酸 (C13:0) Tridecanoic acid 0.08±0.04a 0.08±0.03a 0.06±0a 肉豆蔻酸 (C14:0) Myristic acid 1.39±0.10a 1.29±0.04a 0.97±0.01b 十五碳酸 (C15:0) Pentadecanoic acid 0.85±0.12a 0.96±0.05ab 0.67±0.05ac 棕榈酸 (C16:0) Palmitic acid 22.31±0.38a 20.91±0.32a 18.77±0.71b 十七碳酸 (C17:0) Heptadecanoic acid 1.56±0.05a 2.10±0.3ab 1.28±0.06ac 硬脂酸 (C18:0) Stearic acid 8.75±0.20a 8.74±0.62a 8.47±0.14a 十九碳酸 (C19:0) Nonadecanoic acid 0.21±0.01a 0.27±0.07a 0.15±0.01a 花生酸 (C20:0) Arachidic acid 0.17±0a 0.13±0.01b 0.16±0.02ab 棕榈油酸 (C16:1) Palmitoleic acid 4.86±0.36a 5.9±0.06b 2.82±0.19c 十七碳一烯酸 (C17:1) Heptadecenoic acid 0.26±0.02 ND ND 油酸 (C18:1) Oleic acid 25.36±0.10a 24.13±0.17b 26.56±0.45c 二十碳一烯酸 (C20:1) Eicosenoic acid ND 0.94±0.05 0.99±0.03 二十二碳烯酸 (C22:1) Cetoleic acid ND ND 0.12±0.03 亚油酸 (C18:2) Linoleic acid 9.48±0.72a 6.57±0.25a 20.49±0.13b 亚麻酸 (C18:3) Linolenic acid 0.45±0.04 ND ND 二十碳二烯酸 (C20:2) Eicosadienoic acid 0.49±0.13a 0.42±0.08a 0.62±0.11a 二十碳三烯酸 (C20:3) Eicosatrienoic acid 1.15±0.02a 1.25±0.01a 2.57±0.59b 花生四烯酸 (C20:4) Arachidonic acid 11.10±0.24a 10.13±0.09b 5.63±0.14c 二十碳五烯酸 (C20:5) Eicosapentaenoic acid 1.6±0.03a 3.55±0.05b 1.25±0.02c 二十二碳三烯酸 (C22:3) Docosatrienoic acid ND 0.53±0.40 ND 二十二碳六烯酸 (C22:6) Docosahexaenoic acid 9.92±0.21a 12.03±0.31b 8.37±0.14c EPA+DHA 11.52±0.24a 15.58±0.25b 9.62±0.06c 饱和脂肪酸 SFA 35.34±0.39 34.56±0.17 30.58±1.00 单不饱和脂肪酸 MUFA 30.47±0.48 30.96±0.07 30.49±0.7 多不饱和脂肪酸 PUFA 34.19±0.09 34.48±0.24 38.94±0.30 注:ND. 未检测出 Note: ND. Undetected 2.4 发酵鳜鱼肌肉中矿物质元素分析
发酵鳜鱼所含矿物质元素中钠 (Na) 含量最高,其次为钾 (K)、镁 (Mg)、钙 (Ca)、锌 (Zn)、铁 (Fe)等 (表5),由于加工过程中食盐的加入,使得发酵鳜鱼的Na含量明显高于新鲜鳜鱼,且由于1号发酵鳜鱼在腌制发酵过程中添加了较高的食盐量,使其Na含量高于2号,同时由于盐度越大,钠离子含量急剧增加,而产生的渗透压越大,水分渗出越快,使得发酵鳜鱼中钾离子、镁离子、锌离子、铁离子随水分流失,所以相对含量有所下降;而Ca的含量有所增加,可能由于在发酵菌的作用下,促进了蛋白质降解为肽段,有利用于螯合游离钙离子,所以产品中Ca含量增加;1号发酵鳜鱼中钙离子含量更高,其发酵条件更适宜微生物活动。同时发酵鳜鱼肌肉中含有一定量锰 (Mn)、硒 (Se) 等微量元素,其含量略低于新鲜鳜鱼,Se是人体必需的矿物质元素,在抗炎、抗氧化和调节免疫功能等方面均发挥着重要作用[26]。
表 5 发酵鳜鱼肌肉中矿物质元素质量分数 (干基)Table 5. Mineral elements content in muscle of fermented mandarin fish (dry mass)mg·kg−1 矿物质元素
Mineral element新鲜鳜鱼
Fresh mandarin fish1号发酵鳜鱼
No.1 Fermented mandarin fish2号发酵鳜鱼
No.2 Fermented mandarin fish钠 Na 1 832.23±17.89a 42 865.9±172.75b 35 625.84±44.19c 钾 K 14 778.62±731.17a 5 131.98±140.48b 6 710.36±127.02c 镁 Mg 1 934.08±46.88a 1 008.88±31.32b 1 196.88±33.10c 钙 Ca 119.77±11.97a 286.22±14.92b 236.41±18.17c 锌 Zn 82.27±7.38a 41.05±11.94b 63.33±1.71ab 铁 Fe 55.61±11.82a 33.89±4.16b 21.77±6.57c 锰 Mn 1.84±0.09a 1.23±0.43a 1.09±0.08a 硒 Se 2.81±0.04a 2.19±0.07b 2.65±0.06c 2.5 发酵鳜鱼安全性指标分析
2.5.1 发酵鳜鱼肌肉中TBA分析
脂肪氧化是发酵鱼制品在加工过程中常见的现象之一,而TBA在一定程度上可反映出鱼肉的脂肪氧化程度。两款发酵鳜鱼均发生了较明显的脂肪氧化 (图1-a),由于发酵周期较长,鱼体与空气接触后发生缓慢氧化;而氧化程度可能与发酵微生物作用有关,如木糖葡萄球菌作为发酵菌时,其脂解活性较高,促进游离脂肪酸释放的同时也易引起强烈的脂肪酸氧化[20],且2号发酵鳜鱼经发酵后增加的脂肪酸多为多不饱和脂肪酸,因其不稳定性更易发生氧化。脂肪氧化程度过高将影响鱼肉的品质,同时也可促进醛类、酸类、酯类、酮类等挥发性物质的生成,如在发酵鳜鱼中检出丁酸、乙酸、壬醛、辛醛、胡椒酮等多种挥发性物质[27],使其具有鱼腥味、油脂味、奶油香等多种气味,这有助于发酵风味的形成。
![]() 图 1 发酵鳜鱼肌肉中硫代巴比妥酸值、挥发性盐基氮和亚硝酸盐含量Figure 1. TBA, T-VBN and nitrite content in muscle of fermented mandarin fish
图 1 发酵鳜鱼肌肉中硫代巴比妥酸值、挥发性盐基氮和亚硝酸盐含量Figure 1. TBA, T-VBN and nitrite content in muscle of fermented mandarin fish2.5.2 发酵鳜鱼肌肉中T-VBN分析
T-VBN可用于衡量水产品腐败程度,主要受细菌或酶的作用使蛋白质分解产生氨及胺类等碱性物质。自然发酵的发酵鳜鱼中微生物十分复杂,李燕等[28]发现传统发酵鳜鱼中存在维氏气单胞菌 (Aeromonas veronii)、不可培养气单胞菌、热杀索丝菌 (Brochothrix thermosphacta) 等有害菌,而这些有害菌在发酵过程中得以生长繁殖,不仅加快了鱼肉蛋白降解变质,对产品的安全性也产生一定影响。两款发酵鳜鱼产品T-VBN明显增加,其中1号发酵鳜鱼T-VBN最高,可能是其发酵条件更适于有害菌的生长,经发酵作用后促使鱼肉蛋白降解程度加深,而其产生三甲胺、吲哚等含氮化合物,导致鳜鱼的新鲜度有所下降,另外这类物质具有的“腥臭”气味有助于发酵鳜鱼风味的形成,其“似臭非臭”的独特气味与其他发酵鱼制品有所不同 (图1-b)。
2.5.3 发酵鳜鱼肌肉中亚硝酸盐含量分析
食品中腌制发酵制品中常含有亚硝酸盐,其可抑制肉毒梭状芽胞菌 (Clostridium botulinum) 等有害菌的生长,从而延长货架期;同时亚硝酸盐在胃酸等酸性环境中可转化生成具有致癌性的N-亚硝基化合物[29],不利于人体健康。两款发酵鳜鱼肌肉中亚硝酸盐含量均明显增加,但仍符合我国食品中亚硝酸盐的限量标准 (图1-c)。发酵制品中亚硝酸盐含量增加的原因很多,包括微生物作用、盐浓度、水分和温度等,其中盐浓度是影响亚硝酸盐生成的重要因素,有研究指出低盐高水分的腌制鱼较适宜具备还原能力的微生物生存,从而使得亚硝酸盐含量偏高[30],且盐浓度越高,对具有硝酸盐还原能力的细菌抑制作用也越强,亚硝酸盐生成量也越少[31]。2号发酵鳜鱼中食盐含量低于1号且水分偏高,使其硝酸盐还原菌作用加强,导致亚硝酸盐生成量显著增加。
2.5.4 发酵鳜鱼肌肉中生物胺分析
自然发酵鱼中微生物结构复杂,其中有害细菌在发酵过程中易产生有毒代谢产物如生物胺,而生物胺主要通过氨基酸脱羧酶的作用以及酮、醛的还原胺化反应形成[32]。发酵鳜鱼肌肉中色胺、β-苯乙胺、腐胺、尸胺、组胺、酪胺等含量均有明显增加,且生物胺总量均显著高于新鲜鳜鱼 (表6),但仍在FDA限量范围内[33] (食品中生物胺总量≤1 000 mg·kg−1,其中组胺质量分数≤50 mg·kg−1,酪胺质量分数≤100 mg·kg−1)。色胺、腐胺、尸胺和酪胺是发酵鳜鱼中主要的生物胺,腐胺、尸胺和酪胺也是鲢 (Hypophthalmichthys molitrix) 香肠在发酵过程中形成的主要生物胺[34],其中腐胺和尸胺可与亚硝酸盐反应生成致癌物亚硝胺[35];而组胺、酪胺、β-苯乙胺和色胺对神经系统和血管系统可能有一定的影响[36],且由于尸胺、腐胺、酪胺等生物胺的存在使组胺的毒性增强[37]。对比两款发酵鳜鱼,1号发酵鳜鱼中色胺、尸胺含量显著低于2号,可能原因是起作用的相关微生物较少;而其组胺含量显著高于2号,组胺是由于组氨酸脱羧酶的作用使组氨酸降解产生的,肠杆菌科、假单胞菌科和弧菌 (Vibrio) 等均含有这种酶[38],其原因可能是1号发酵鳜鱼的加工环境更适于这类细菌的生长。
表 6 发酵鳜鱼肌肉中生物胺质量分数 (湿基)Table 6. Biogenic amines content in muscle of fermented mandarin fish (wet mass)mg·kg−1 生物胺
Biogenic amine新鲜鳜鱼
Fresh mandarin fish1号发酵鳜鱼
No.1 Fermented mandarin fish2号发酵鳜鱼
No.2 Fermented mandarin fish色胺 Tryptamine 2.81±0.21a 24.34±1.20b 36.98±0.17c β-苯乙胺 β-pheny-lethylamine 1.22±0.07a 9.71±0.07b 11.61±1.49b 腐胺 Putreseine 1.69±0.11a 46.56±0.04b 47.21±2.79 b 尸胺 Cadaverine 4.30±4.18 a 34.33±0.65b 65.48±4.66c 组胺 Histamine 5.37±0.09a 25.52±2.30b 10.62±0.08c 酪胺 Tyramine 10.89±0.49a 28.69±1.29b 31.45±0.83b 亚精胺 Spermidine 4.61±0.05a 5.23±0.52a 5.58±0.61a 精胺 Spermine 8.53±0.63a 8.98±0.71a 8.80±1.12a 总生物胺 Total biogenic amine 39.41±4.07a 183.36±0.54b 217.73±8.36c 2.5.5 发酵鳜鱼肌肉中重金属含量分析
必需的矿物质元素在人体正常生理活动中不可或缺,而铬 (Cr)、砷 (As)、铅 (Pb)等重金属元素并不参与生理活动且摄入后易造成重金属的积累而对人体造成危害[39]。两款发酵鳜鱼干样中Cr和镉 (Cd) 含量与新鲜鳜鱼相比无显著性差异,而As和汞 (Hg) 含量均略低于新鲜鳜鱼,Pb含量略高于新鲜鳜鱼 (表7),各重金属含量均符合我国食品中重金属限量标准 (按湿基换算)。相比两款发酵鳜鱼,1号发酵鳜鱼中Cr、Cd 和Pb含量略高于2号,而As和Hg含量较低,这可能与其原料鱼的生存环境、投喂饲料成分有关,造成重金属富集量略有差异,故发酵鳜鱼的生产加工同样要注重原材料的选择。
表 7 发酵鳜鱼肌肉中重金属质量分数 (干基)Table 7. Heavy metals content in muscle of fermented mandarin fish (dry mass)mg·kg−1 重金属
Heavy metal新鲜鳜鱼
Fresh mandarin fish1号发酵鳜鱼
No.1 Fermented mandarin fish2号发酵鳜鱼
No.2 Fermented mandarin fish铬 Cr 2.623±0.168a 2.602±0.24a 2.041±0.278a 砷 As 0.134±0.022a 0.022±0b 0.101±0a 镉 Cd 0.012 6±0.001a 0.005 9±0a 0.002 3±0a 汞 Hg 0.054±0.003a 0.014 0±0.001b 0.017 3±0.009b 铅 Pb 0.770±0.091a 1.216±0.036b 1.154±0.033b 3. 结论
两款发酵鳜鱼氨基酸种类齐全,鲜味氨基酸含量均高于新鲜鳜鱼,从而提升了鱼肉的鲜美滋味,其氨基酸模式与必需氨基酸指数略高于新鲜鳜鱼,且2号发酵鳜鱼必需氨基酸指数高于1号;发酵鳜鱼中脂肪酸种类较全且总不饱和脂肪酸含量高于新鲜鳜鱼,而两款发酵鳜鱼多不饱和脂肪酸含量有较大差异,2号发酵鳜鱼中多不饱和脂肪酸含量更高;发酵鳜鱼中均含有多种矿物质元素,发酵后Na和Ca含量明显增加。整体而言,发酵鳜鱼属于营养较为全面且味道良好的发酵鱼制品。在食用安全性方面,两款发酵鳜鱼重金属含量均未超标,而TBA、T-VBN、亚硝酸盐和总生物胺含量均高于新鲜鳜鱼,其中1号发酵鳜鱼中T-VBN和组胺含量高于2号,而亚硝酸盐与总生物胺含量均低于2号。
现阶段常用的发酵鳜鱼加工方式较为传统且不统一,不同地区原料鱼自身携带的微生物也存在一定差异,同时受加工条件、环境因素的影响,发酵鳜鱼常常存在不同程度的脂肪氧化、蛋白质降解和生物胺积累等问题,使其产品品质较不稳定,产品间差异性也较大。此外,目前所参照的水产食品安全标准大多针对鲜、冻动物性水产品,尚未建立健全与发酵鳜鱼或发酵鱼制品相关的食用安全标准,难以对发酵鱼制品品质进行系统性评价。因此,亟需深入研究以建立并完善发酵鳜鱼品质控制技术,制定相关的规范标准,控制不良因素的发生,加强其稳定性和安全性,全面提升产品品质,促进发酵鳜鱼企业化发展。
-
![]()
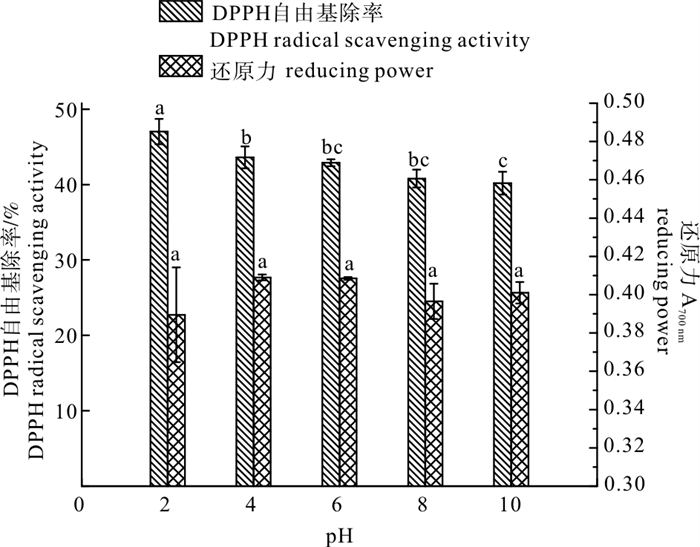
图 1 pH对酶解产物抗氧化稳定性的影响
Figure 1. Effect of pH on antioxidant stability of enzymatic hydrolysates
表 1 不同分子量组分的抗氧化活性
Table 1 Antioxidant activity of different molecular weights peptides
分子量
molecular weightDPPH自由基清除率IC50/mg·mL-1
DPPH radical scavenging activity·OH清除率IC50/mg·mL-1
·OH scavenging activity还原力A700 nm
reducing power酶解液enzymatic hydrolysate 6.08±0.21a 15.52±0.08a 0.662±0.085a >5 kD 6.58±0.07b 16.05±0.74b 0.472±0.025b < 5 kD 5.04±0.12c 15.16±0.16c 0.717±0.067c  下载: 导出CSV
下载: 导出CSV
表 2 温度对酶解产物抗氧化稳定性的影响
Table 2 Effect of temperature on antioxidant stability of enzymatic hydrolysates
温度/℃
temperaturet/min DPPH自由基清除率/%
DPPH radical scavenging activity还原力A700 nm
reducing power25 30 54.52±2.39aA 0.341±0.026aA 60 54.33±0.94aA 0.352±0.034aA 90 51.87±2.31aA 0.361±0.02aA 120 51.68±0.6aA 0.34±0.012aA 40 30 54.79±0.11aA 0.352±0.013aA 60 52.19±1.46bA 0.350±0.018aA 90 51.77±0.47bcA 0.343±0.020aA 120 49.77±0.13cA 0.328±0.029aA 60 30 50.93±0.2aB 0.334±0.014aA 60 47.08±2.22bB 0.335±0.185aAB 90 47.63±0.65bB 0.331±0.025aAB 120 45.07±0.04bB 0.331±0.013aA 80 30 45.67±0.15aC 0.332±0.028aA 60 45.40±0.9aBC 0.307±0.044aBC 90 40.47±0.34bC 0.303±0.031aBC 120 42.15±2.06bC 0.276±0.024bB 100 30 44.27±1.14aC 0.328±0.033aA 60 41.99±0.01bC 0.291±0.011abC 90 40.90±0.09bC 0.285±0.030bC 120 36.66±0.67cD 0.279±0.024bB 注:同列标有不同小写字母表示相同温度不同时间下差异显著(P < 0.05),标有不同大写字母表示不同温度相同时间下差异显著(P < 0.05),标有相同字母表示差异不显著(P>0.05)。
Note:Different small letters within the same column mean significant difference under the same temperature and different time (P < 0.05);different capital letters mean significant difference at different temperatures and at the same time (P < 0.05);the same letters indicate no significant difference (P>0.05).
下载: 导出CSV
表 3 不同光照处理对酶解产物抗氧化稳定性的影响
Table 3 Effect of light on antioxidant stability of enzymatic hydrolysates
不同光照处理
different light processingt/h DPPH自由基清除率/%
DPPH radical scavenging activity还原力A700 nm
reducing power避光dark place 1 50.28±0.72aA 0.405±0.023aA 2 48.28±0.72aA 0.412±0.044aA 3 49.87±1.46aA 0.384±0.026aA 4 48.19±0.58aA 0.396±0.027aA 5 47.10±3.48aA 0.360±0.013aA 室内自然光natural light 1 49.86±0.43aA 0.357±0.008aB 2 47.25±0.12bAB 0.356±0.024aA 3 45.57±1.45abA 0.365±0.024aA 4 46.42±0.23abA 0.336±0.019aB 5 44.19±2.29abB 0.357±0.023aA 日光照射solar radiation 1 41.88±0.12aB 0.318±0.033aB 2 41.06±0.73aB 0.289±0.039aB 3 40.59±1.16abB 0.320±0.023aA 4 39.65±0.51abA 0.324±0.015aA 5 39.39±0.57bC 0.338±0.024aA 注:同列标有不同小写字母表示相同光照处理不同时间下差异显著(P < 0.05),标有不同大写字母表示不同光照处理相同时间下差异显著(P < 0.05),标有相同字母表示差异不显著(P>0.05)。
Note:Values with different small letters in the same column indicate significant difference with the same light processing at different time (P < 0.05);values with different capital letters indicate significant difference at different light processing time (P < 0.05);values with the same letters indicate no significant difference (P > 0.05).
下载: 导出CSV
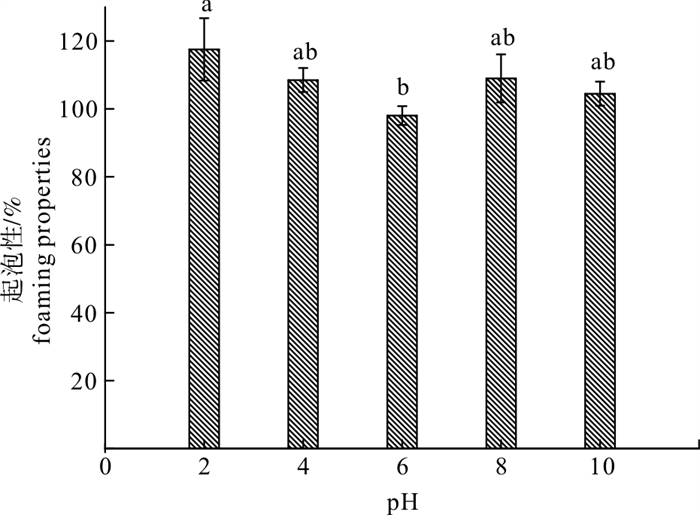
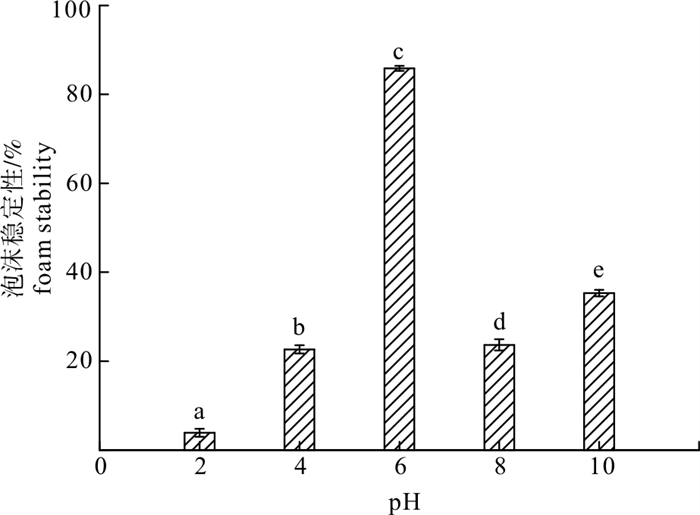
表 4 酶解产物的乳化活性和乳化稳定性
Table 4 Emulsifying property and emulsion stability of enzymatic hydrolysates
pH 2 4 6 8 10 乳化活性指数/cm2·g-1 EAI 74.58±3.84a 55.71±0.4b 40.83±1.04c 106.42±4.17d 154.76±2.83e 乳化稳定性指数/min ESI 19.95±2.52a 28.71±1.37ab 33.69±1.90b 23.98±3.41a 51.93±3.51c
下载: 导出CSV
-
[1] 江艳娥, 陈作志, 林昭进, 等. 南海中部海域鸢乌贼耳石形态特征分析[J]. 南方水产科学, 2014, 10(4): 85-90. doi: 10.3969/j.issn.2095-0780.2014.04.014 [2] 晏磊, 张鹏, 杨吝, 等. 月相对南海灯光罩网鸢乌贼渔获率的影响分析[J]. 南方水产科学, 2015, 11(3): 16-21. doi: 10.3969/j.issn.2095-0780.2015.03.003 [3] 张鹏, 杨吝, 张旭丰, 等. 南海金枪鱼和鸢乌贼资源开发现状及前景[J]. 南方水产, 2010, 6(1): 68-74. doi: 10.3969/j.issn.1673-2227.2010.01.012 [4] FUJⅡ T, AHN J Y, KUSE M, et al. A novel photoprotein from oceanic squid (Symplectoteuthis oualaniensis) with sequence similarity to mammalian carbon-nitrogen hydrolase domains[J]. Biochem Bioph Res Commun, 2002, 293(2): 874-879. doi: 10.1016/S0006-291X(02)00296-6
[5] MASAKI K. Chromophores in photoproteins of a glowing squid and mollusk[J]. Biosci Biotechnol Biochem, 2014, 78(5): 731-736. doi: 10.1080/09168451.2014.915724
[6] WANG Q, XUE C, LI Z, et al. Analysis of DHA-rich phospholipids from egg of squid Sthenoteuthis oualaniensis[J]. J Food Compos Anal, 2008, 21(4): 356-359. doi: 10.1016/j.jfca.2008.02.004
[7] 隋晓, 冯晓梅, 岳荣岩, 等. 亚临界R134a脱除鸢乌贼卵中胆固醇的工艺研究[J]. 核农学报, 2015, 29(6): 1129-1134. doi: 10.11869/j.issn.100-8551.2015.06.1129 [8] 马静蓉, 杨贤庆, 马海霞, 等. 南海鸢乌贼肌原纤维蛋白的热稳定性研究[J]. 食品与发酵工业, 2015, 41(5): 80-84. doi: 10.13995/j.cnki.11-1802/ts.201505014 [9] JE J Y, LEE K H, LEE M H, et al. Antioxidant and antihypertensive protein hydrolysates produced from tuna liver by enzymatic hydrolysis[J]. Food Res Int, 2009, 42(9): 1266-1272. doi: 10.1016/j.foodres.2009.06.013
[10] 王晶, 吴燕燕, 李来好, 等. 响应面法优化微波辅助酶解合浦珠母贝蛋白工艺[J]. 食品科学, 2014, 35(10): 11-17. doi: 10.7506/spkx1002-6630-201410003 [11] ZHAO Q, XIONG H, SELOMULYA C, et al. Enzymatic hydrolysis of rice dreg protein: effects of enzyme type on the functional properties and antioxidant activities of recovered proteins[J]. Food Chem, 2012, 134(3): 1360-1367. doi: 10.1016/j.foodchem.2012.03.033
[12] 王晶, 吴燕燕, 李来好, 等. 合浦珠母贝肉酶解产物的抗氧化稳定性与功能特性[J]. 食品工业科技, 2014, 35(13): 67-71. doi: 10.13386/j.issn1002-0306.2014.13.005 [13] ZENG M, ADHIKARI B, HE Z, et al. Improving the foaming properties of soy protein isolate through partial enzymatic hydrolysis[J]. Dry Technol, 2013, 31(13/14): 1545-1552.
[14] 祝婧, 刘磊, 张名位, 等. 不同分子量海鲈鱼胶原蛋白肽组分的功能特性比较[J]. 现代食品科技, 2014, 30(12): 113-118. doi: 10.13982/j.mfst.1673-9078.2014.12.019 [15] 牛慧慧. 蛋清肽制备及其生物功能性研究[D]. 武汉: 华中农业大学, 2011: 89. 10.7666/d.y2004217 [16] HE X H, LIU H Z, LIU L, et al. Effects of high pressure on the physicochemical and functional properties of peanut protein isolates[J]. Food Hydrocolloid, 2014, 36: 123-129. doi: 10.1016/j.foodhyd.2013.08.031
[17] 游丽君. 泥鳅蛋白抗氧化肽的分离纯化及抗疲劳、抗癌功效研究[D]. 广州: 华南理工大学, 2010: 110. https://cdmd.cnki.com.cn/Article/CDMD-10561-2010228252.htm [18] 孙常雁, 李德海, 刘骞, 等. 乳清蛋白肽美拉德反应产物的抗氧化活性与稳定性[J]. 食品科学, 2014, 35(1): 104-109. doi: 10.7506/spkx1002-6630-201401020 [19] 游丽君, 赵谋明, JOE R, 等. 加工和贮藏条件对泥鳅多肽抗氧化活性的影响[J]. 江苏大学学报(自然科学版), 2009, 30(6): 549-553. doi: 10.3969/j.issn.1671-7775.2009.06.003 [20] 胡晓, 武琼, 杨贤庆, 等. 裂壶藻渣酶解产物的抗氧化稳定性[J]. 食品科学, 2015, 36(11): 21-26. doi: 10.7506/spkx1002-6630-201511005 [21] 吴燕燕, 王晶, 李来好, 等. 添加剂和光照对合浦珠母贝肉酶解液抗氧化活性的影响[J]. 食品工业科技, 2015, 36(2): 149-156. doi: 10.13386/j.issn1002-0306.2015.02.024 [22] 孙燕婷, 黄国清, 孙萍, 等. 大豆分离蛋白溶解性和乳化性影响因素研究[J]. 粮油食品科技, 2011, 19(1): 32-35. [23] 杜艳萍, 刘春雷, 闵伟红, 等. 长白山榛仁分离蛋白及其主要组分的功能性质研究[J]. 现代食品科技, 2015, 31(5): 109-115. doi: 10.13982/j.mfst.1673-9078.2015.5.018 [24] SEENA S, SRIDHAR K R. Physicochemical, functional and cooking properties of under explored legumes, Canavalia of the southwest coast of India[J]. Food Res Int, 2005, 38(7): 803-814. https://www.sciencedirect.com/science/article/pii/S0963996905000700
[25] SURESH K K, GANESAN K, SELVARAJ K, et al. Studies on the functional properties of protein concentrate of Kappaphycus alvarezii (Doty) Doty: an edible seaweed[J]. Food Chem, 2014, 153: 353-360. https://www.sciencedirect.com/science/article/pii/S0308814607008102
-
期刊类型引用(3)
1. 向劲,吴启藩,宋锐,彭治桃,李金龙,高峰,谢敏,程小飞. 循环水养殖池塘中浮游动植物功能群特征及水质评价. 水产学杂志. 2024(01): 81-89 .  百度学术
百度学术
2. 王保,洪涛,刘可群,汤阳,叶佩,姜润. 夏季晴天鱼塘水温垂直变化规律及原因分析. 中国农学通报. 2024(12): 150-158 . 百度学术
3. 贺克雕,马瑞君,范莹,杨世美,和杰梅,段昌群. 泸沽湖水生生物群落及水生态环境质量研究. 西部林业科学. 2023(04): 32-39 . 百度学术
其他类型引用(5)




计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 8
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
