
Composition and biodiversity of catches by bottom trawl fishery in southwestern continent shelf of Nansha area
-
摘要:
根据2013年对南沙西南陆架海域的4个航次底拖网调查数据,分析了渔获组成及生物多样性指数。结果显示,4个航次调查期间共收集并记录渔获物251种,其中鱼类202种、头足类16种、甲壳类33种,渔获种类夏季最多,冬季最少。根据计算相对重要性指数(IRI),短尾大眼鲷(Priacanthus macracanthus)、剑尖枪乌贼(Uroteuthis edulis)和多齿蛇鲻(Saurida tumbil)为优势种(IRI≥500);深水金线鱼(Nemipterus bathybius)、蛇鲭(Gemphylus serpens)、黑
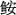
According to the bottom trawl survey data in southwestern continent shelf of Nansha area in different seasons of 2013, we analyzed the species composition and biodiversity indices. We collected 251 species, including 202 fish species, 16 cephalopod species and 33 crustacean species. The total number of species of catch composition was the highest in summer and the lowest in winter. According to the index of relative importance (IRI), the dominant species were Priacanthus macracanthus, Uroteuthis edulis and Saurida tumbil, whose IRIs were higher than 500;the main species were Nemipterus bathybius, Gemphylus serpens, Lophiomus setigerus, Trachinocephalus myops, Trichiurus lepturus, Saurida undosquamis, Decapterus macrosoma, and so on, whose IRIs were 100~500. Cluster analysis indicates that the dominant species structure was more similar between spring and autumn. The Margalef richness index (D) reached the maximum in summer but the the the minimum in winter; the Shannon-Weiner diversity index (H′) reached the maximum in summer but the minimum in autumn; Pielou index (J′) reached the minimum in autumn, but there was little difference among the other three seasons; dominance index (λ) reached the maximum in autumn but the minimum in summer.
-
盐度是影响河口、过河口及海水水生生物生长和发育的重要环境因子,与水生生物的性腺发育关系密切。研究表明,成体金钱鱼 (Scatophagus argus)[1]、印度囊鳃鲶 (Heteropneustes fossilis)[2]、红眼沙鲈(Psammoperca waigiensis)[3-4]、梭鱼 (Liza haematocheila)[5]、凡纳滨对虾 (Litopenaeus vannamei)[6]、中华绒螯蟹(Eriocheir sinensis)[7]等的性腺需要在一定盐度刺激下才可正常发育。
刀鲚 (Coilia nasus) 是长江口重要的洄游性经济鱼类[8],主要分布于中国、韩国沿海以及日本的有明海[9-12],因味道鲜美、肉质细嫩成为重要的经济鱼类[13]。近年来由于水体污染、水利兴修、资源过度开发等因素,进入产卵场的刀鲚种群数量显著降低,以致不能形成常规的渔汛[14-15]。盐度是刀鲚在洄游过程中最直观的影响因素[16],研究显示,进行生殖洄游的鱼类须到达特定盐度才能完成卵巢的正常发育,才会发生产卵行为[17],如鲥 (Tenualosa reevesii)、暗纹东方鲀 (Takifugu obscurus)、大麻哈鱼 (Oncorhynchus keta) 的性腺发育和成熟仅在盐度低于0.5的淡水环境中进行;而日本鳗鲡 (Anguilla japonica)、松江鲈 (Trachidermus fasciatus)的性腺发育与繁殖需要盐度较高的海水环境[17]。近年,Xu等[18]通过代谢组学对刀鲚卵巢发育不同步的问题进行了研究,Yin等[19]通过代谢组学研究了刀鲚在产卵迁移过程中能量的消耗和储存,但未明确指出海、淡水对卵巢发育的影响。为了更好地保护并合理利用刀鲚资源,需要通过人工养殖来维持其资源量,因此,对刀鲚在人工海、淡水环境下卵巢发育机制的研究十分重要。
代谢组学技术作为近年新兴的一种高通量测序技术,在各研究领域尤其是环境对物种的影响中得到广泛应用[20-22],代谢组代表生物细胞、组织、器官或生物体中的完整代谢产物,这些代谢产物可以通过代谢组学的方法进行测定[23]。关于刀鲚卵巢发育的代谢组学研究较少,本研究主要通过液相色谱-质谱联用 (Liquid chromatograph-mass spectrometer, LC-MS) 的代谢组学技术分析筛选养殖在淡水和海水2种环境下刀鲚卵巢组织的差异代谢产物,并对差异代谢产物进行代谢通路富集分析,为进一步研究刀鲚在生殖洄游过程中卵巢组织的发育及其机制以及人工养殖提供参考依据。
1. 材料与方法
1.1 实验材料
实验于2018年12月—2019年2月在上海市水产研究所奉贤基地进行,实验材料选用该基地提供的野生型F3代,Ⅱ龄刀鲚。实验前将刀鲚放入淡水池中 (长5 m×宽3 m×高1 m) 中暂养2周,其后随机分到淡水组 (实验组) 和海水组 (对照组,盐度为25) 水泥池中养殖,每组100尾。海水从河口抽取,盐度约为9,用日晒盐将盐度提高到25。每天08:00、15:00投喂从池塘捞取的活饵 (桡足类、糠虾等),每14 d换水1次。
1.2 实验方法
1.2.1 采样方法
养殖实验结束后,停食1 d。将刀鲚置于30 mg·L−1丁香酚溶液中麻醉5 min后,用干毛巾将鱼体表面擦干,取其性腺,一半装于2 mL的冻存管,迅速置于液氮罐中冷冻,之后于−80 ℃保存以备刀鲚卵巢代谢组学分析;另一半置于Bouin's液中固定24 h,通过石蜡切片和HE染色确定性别后,将海、淡水刀鲚的卵巢分别每3个样混合,每组6个平行,平均体长为 (21.00±2.67) cm,平均体质量为 (25.80±9.05) g。
1.2.2 样本处理
称取刀鲚卵巢样本20 mg ,加入400 μL提取液 [V(甲醇)∶V(水)=4∶1],利用高通量组织破碎仪破碎 (−20 ℃, 50 Hz, 6 min);涡旋 (30 s) 混匀后,低温超声萃取30 min (5 ℃, 40 kHz);将其置于−20 ℃ 30 min;离心(13 000×g, 4 ℃, 15 min),去掉上清液,并加入100 μL复溶液 [V(乙腈)∶V(水)=1∶1] 复溶,同时将所有样本的提取液等体积混匀制备质控样本 (Quality control, QC),每个QC的体积与样本体积相同。将所测样本和QC转移至LC-MS 进样瓶进行上机分析。本次LC-MS 分析的仪器平台为AB SCIEX公司的超高效液相色谱串联飞行时间质谱UPLC-TripleTOF系统。
1.2.3 色谱条件
实验采用美国Milford公司BEH C18色谱柱 (100 mm × 2.1 mm,1.7 μm)。配置流动相A 为水 (含0.1%甲酸),流动相B 为V(乙腈)∶V(异丙醇)=1∶1 (含0.1%甲酸),进行梯度洗脱,流速为0.40 mL·min−1,进样量为20 μL,柱温为40 ℃。
1.2.4 质谱条件
样品质谱信号采集分别采用正负离子扫描模式、离子喷雾电压。质谱检测范围为50~1 000 m.z−1。ESI离子源参数设置雾化气压 (GS1) 50 psi;辅助气压 (GS2) 50 psi;气帘气压:30 psi;温度:500 ℃;喷雾电压:5 000 V (POS模式)或−4 000 V (NEG模式)。
1.3 数据处理与分析
首先采用代谢组学Progenesis QI处理软件 (Waters Corporation, Milford, USA)过滤质谱数据的低质量峰,填充缺失值,归一化,评估QC样本RSD,数据log转换预处理。将原始数据用Progenesis QI进行搜库鉴定,将质谱信息与代谢数据库HMDB和METLIN进行匹配。
采用R软件包Ropls进行多元统计分析:首先将所有实验样品与QC样品比较后进行主成分分析 (Principal Component Analysis, PCA)和正交偏最小二乘判别分析(Orthogonal Partial Least Squares Discrimination Analysis, OPLS-DA),再经200次置换检验以判断模型是否过拟合。
采用t检验并结合OPLS-DA分析中的VIP值,筛选出组间的差异代谢物,筛选条件为差异倍数(Fold-change FC>1)、P(<0.05)和变量投影重要度(Variable importance in the projection, VIP>1),三者取交集。代谢通路的注释分析基于KEGG数据库。
将得到的差异代谢产物在KEGG Compound和HMDB数据库进行化合物分类情况统计。
2. 结果
2.1 多元统计分析结果
首先对淡水组 (N=6) 和海水组 (N=6) 的样品进行PCA和OPLS-DA分析以评估两组间的统计学差异,发现所有样品均处于样本置信区间。十字架表示的质控样本比较集中,说明仪器的稳定性好,检测的数据可靠,可用于PCA和OPLS-DA分析。PCA得分结果显示,淡水组和海水组样品点在空间上具有明显的分离,表明两组间的代谢模式存在差异 (图1-a);OPLS-DA分析可看出两组数据的样品点分别聚为一簇,在空间上呈现分离状态,表明淡水组和海水组间产生了显著差异 (图1-b),模型的评价指标R2=0.971,Q2=0.562,说明所构建的模型良好。为进一步验证模型是否过拟合,采用200次置换检验 (图2),可知Q2于y轴的截距为负,且置换检验得到的R2=0.873 5和Q2=−0.146 4均小于原始值,由此可见模型没有过拟合,可以用于后续差异代谢物筛选的分析。
![]() 图 1 海水组和淡水组的主成分分析 (a) 和正交偏最小二乘判别分析 (b) 得分图Figure 1. PCA (a) and OPLS-DA (b) scores of seawater and freshwater
图 1 海水组和淡水组的主成分分析 (a) 和正交偏最小二乘判别分析 (b) 得分图Figure 1. PCA (a) and OPLS-DA (b) scores of seawater and freshwater2.2 总代谢产物Venn图
对在阳离子和阴离子2种模式下获得的海、淡水养殖刀鲚的卵巢代谢产物数据分别构建Venn图(图3)。在阳离子分离模式下,2组卵巢组织中共有代谢产物310个,其中淡水组中特有代谢产物9个,海水组12个;在阴离子分离模式下,2组共有代谢产物392个,其中淡水组中特有代谢产物也为9个,海水组则降到10个。
2.3 差异代谢产物注释
海水和淡水养殖刀鲚卵巢组织样本中的差异代谢产物的火山图见图4,差异代谢产物具体信息详见表1。与海水组相比,淡水组刀鲚卵巢组织有47个代谢产物表现出显著差异性,其中11个代谢产物有上调表达,36个代谢产物有下调表达,表达差异程度最显著的为碳环血氧烷A2 (Carbocyclic thromboxane A2)、半乳糖神经酰胺 (Galactosyl ceramide),差异倍数分别为10.40、2.78倍。根据表1,淡水组的皮质醇含量比海水组高1.61倍,且呈显著性差异 (P<0.05)。
表 1 差异代谢物信息表Table 1. Differential metabolites information sheet差异代谢物
Differential metabolite分子式
Molecular formula变量投影重要度
VIPFC
Fold changeP 变化趋势
Variation trend磷脂酰肌醇 PI [20:4(5Z,8Z,11Z,14Z)/0:0] C29H49O12P 1.58 0.93 0.017 下降 谷氨酰胺色氨酸 Glutaminyltryptophan C16H20N4O4 1.05 0.96 0.018 下降 L-异亮氨酸 L-Isoleucine C6H13NO2 1.33 0.96 0.005 下降 木麻黄6-α-D-葡萄糖苷 Casuarine 6-alpha-D-glucoside C14H25NO10 1.13 0.94 0.043 下降 甘油一脂 MG(10:0/0:0/0:0) C13H26O4 3.59 0.71 0.025 下降 皮质醇 Cortisol C21H30O5 3.28 1.61 0.008 上升 碳环血氧烷A2 Carbocyclic thromboxane A2 C22H36O3 2.88 10.40 0.007 上升 (±)9-十八碳二烯酸 (±)9-HPODE C18H32O4 2.90 0.58 0.020 下降 N-棕榈酰蛋氨酸 N-palmitoyl methionine C21H41NO3S 1.86 0.75 0.022 下降 13-羟基十八酸 13-hydroxyoctadecanoic acid C18H36O3 1.95 0.86 0.005 下降 咖啡酰环戊醇 Caffeoylcycloartenol C39H56O4 2.07 0.81 0.036 下降 11,13-二十碳二烯酸 15-OxoEDE C20H34O3 2.51 0.70 0.032 下降 2,3-二氢苯并呋喃 2,3-dihydrobenzofuran C8H8O 1.19 1.08 0.033 上升 4-甲酰基吲哚 4-formyl indole C9H7NO 1.34 0.83 0.040 下降 红花素C Safflomin C C30H30O14 2.04 0.85 0.003 下降 6-[(2-羧基乙酰基)氧]-3,4,5-三羟基氧烷-2-羧酸6-[(2-carboxyacetyl)oxy]-3,4,5-trihydroxyoxane-2-carboxylic acid C9H12O10 2.07 1.12 0.001 上升 异戊二烯 Isoputreanine C7H16N2O2 2.33 0.84 0.001 下降 脯氨酸 L-Proline C5H9NO2 1.80 0.91 0.003 下降 羟脯氨酸 Hydroxyprolyl-hydroxyproline C10H16N2O5 3.13 0.72 0.007 下降 5-O-阿魏酰黑精 5-O-Feruloylnigrumin C21H25NO10 1.0 0.95 0.037 下降 壬二酸 Azelaic acid C9H16O4 1.12 0.96 0.012 下降 尿苷 Uridine C9H12N2O6 1.35 0.95 0.001 下降 视黄酯 Retinyl ester C20H30O2 1.24 0.96 0.004 下降 顺-9,10-环氧硬脂酸 cis-9,10-Epoxystearic acid C18H34O3 1.66 0.92 0.025 下降 异柠檬酸盐 Isocitrate C6H8O7 1.55 1.12 0.047 上升 戊二酸 Glutaric acid C5H8O4 2.53 1.38 0.015 上升 6-脱氧噬菌体胺 (6-脱氧花青) 6-Deoxyfagomine C6H13NO2 1.36 0.94 0.012 下降 乙酰-L-酪氨酸 Acetyl-L-tyrosine C11H13NO4 1.38 0.87 0.010 下降 γ-谷氨酰鸟氨酸 Gamma glutamyl ornithine C10H19N3O5 1.87 1.58 0.023 上升 对茴香酸异戊酯 (异戊基异茴香酸酯) Isoamyl p-anisate C13H18O3 1.76 0.67 0.041 下降 3-氧十二酸 3-Oxododecanoic acid C12H22O3 1.01 0.92 0.043 下降 C-2神经酰胺 C-2 Ceramide C20H39NO3 2.21 0.75 0.028 下降 磷脂酰乙醇胺 PE(15:0/16:1(9Z)) C36H70NO8P 1.42 1.10 0.030 上升 半乳糖神经酰胺 Galactosyl ceramide (d18:1/14:0) C38H73NO8 1.97 2.78 0.045 上升 N-棕榈酰甘氨酸 N-Palmitoyl glycine C18H35NO3 1.97 0.86 0.000 下降 二十碳五烯酸 Eicosapentaenoic acid C20H30O2 1.01 0.98 0.004 下降 溶血磷脂酰乙醇胺 LysoPE(0:0/20:2(11Z,14Z)) C25H48NO7P 1.33 1.12 0.037 上升 9-羟基癸酸 9-Hydroxydecanoic acid C10H20O3 1.21 0.92 0.028 下降 甘油一脂 MG(a-13:0/0:0/0:0)[rac] C16H32O4 1.92 0.79 0.040 下降 反式-2-十二碳烯二酸 Traumatic acid C12H20O4 1.37 0.86 0.025 下降 9-氧壬酸 9-Oxo-nonanoic acid C9H16O3 1.15 0.93 0.019 下降 8-羟基-5,6-辛二烯酸 8-Hydroxy-5,6-octadienoic acid C8H12O3 1.04 0.94 0.016 下降 (S)-3-磺酸盐 (S)-3-Sulfonatolactate C3H6O6S 2.57 3.05 0.046 上升 γ-谷氨酰缬氨酸 gamma-Glutamylvaline C10H18N2O5 1.80 0.87 0.008 下降 穗花牡荆苷 Agnuside C22H26O11 1.41 0.91 0.005 下降 假尿苷 Pseudouridine C9H12N2O6 1.19 0.93 0.003 下降 雌三醇7-(6-反式-对-香豆酰基葡萄糖苷)
Eriodictyol 7-(6-trans-p-coumaroylglucoside)C30H28O13 1.44 0.92 0.020 下降 注:FC表示某差异代谢物在淡水组相对于海水组的表达倍数变化,FC>1表示该代谢物上调,FC<1表示该代谢物下调;P<0.05表示差异显著,P<0.01表示差异极显著。 Note: FC value indicates the change of expression multiple of a differential metabolite in FOV group compared with SOV group. FC>1 indicates that the metabolite is upregulated; FC<1 indicates that the metabolite is down-regulated. P<0.05 indicates significant difference, and P<0.01 indicates extremely significant difference. 将上述47个差异表达代谢产物比对到KEGG 数据库进行富集分析,共有8个代谢产物显著富集到12条KEGG通路上 (图5和表2)。显著差异代谢产物的富集通路主要涉及氨酰-tRNA生物合成 (Aminoacyl-tRNA biosynthesis)、嘧啶代谢 (Pyrimidine metabolism)、鞘磷脂代谢 (Sphingolipid metabolism)和三羧酸循环 (TCA cycle)。上调表达的2个代谢产物L-脯氨酸 (L-Proline)和L-异亮氨酸 (L-Isoleucine)参与到氨酰tRNA生物合成通路上。下调表达的2个代谢产物尿苷和假尿苷参与到嘧啶代谢通路中。在实验组卵巢组织中上调表达的半乳糖神经酰胺和异柠檬酸 (Isocitrate) 代谢物分别参与到鞘磷脂代谢和三羧酸循环通路中。
表 2 差异代谢产物的KEGG通路富集表Table 2. KEGG pathway enrichment table of differential metabolites富集的差异代谢产物名称
Enriched differential metabolite name富集个数
Numble富集通路
ID Pathway通路描述
Pathway desciptionP 半乳糖神经酰胺 Galactosyl ceramide (d18:1/14:0) 1 map00600 鞘脂代谢 0.059 脯氨酸 L-Proline 1 map00330 精氨酸和脯氨酸代谢 0.176 异柠檬酸盐 Isocitrate 1 map00020 柠檬酸循环 0.057 戊二酸 Glutaric acid 1 map00310 赖氨酸降解 0.130 皮质醇 Cortisol 1 map00140 类固醇激素生物合成 0.207 尿苷 Uridine;假尿苷 Pseudo uridine 2 map00240 嘧啶代谢 0.011 异柠檬酸盐 Isocitrate 1 map00630 乙醛酸和二羧酸代谢 0.137 L-异亮氨酸 L-Isoleucine;脯氨酸 L-Proline 2 map00970 氨酰tRNA生物合成 0.008 L-异亮氨酸 L-Isoleucine 1 map00290 缬氨酸、亮氨酸和异亮氨酸生物合成 0.064 二十碳五烯酸 Eicosapentaenoic acid 1 map01040 不饱和脂肪酸生物合成 0.097 L-异亮氨酸 L-Isoleucine 1 map00280 缬氨酸、亮氨酸和异亮氨酸降解 0.107 (S)-3-磺酸盐 (S)-3-Sulfonatolactate 1 map00270 半膀氨酸和蛋氨酸代谢 0.141 将得到的差异代谢产物比对到HMDB数据库,统计发现占比最多的是脂质和类脂分子,高达41.18%;其次为有机酸及其衍生物,占比35.29%。
将代谢差异物比对到KEGG Compound数据库,发现聚类最多的是氨基酸和羧酸;比对到KEGG Pathway数据库,发现富集程度最高的分别是脂质代谢通路 (Lipid metabolism)、氨基酸代谢通路和消化系统。
3. 讨论
通过非靶向代谢组学得到刀鲚卵巢在海、淡水环境下47种差异代谢物,发现代谢物主要包括脂质和类脂、有机酸及衍生物、氨基酸及羧酸,这与Xu等[18]对刀鲚卵巢Ⅲ—Ⅴ期发育过程的研究结果相似,差异代谢物主要涉及糖酵解和糖异生、组氨酸代谢、赖氨酸代谢以及脂肪酸的代谢相关通路。由此可见,海、淡水环境对人工养殖刀鲚的卵巢发育具有重要作用。
碳环血氧烷A2是由活化的血小板在止血过程中产生的,具有凝血之前的特性,也是一种已知的血管收缩剂[24-26],本实验淡水组碳环血氧烷A2高表达可能是刀鲚机体组织为应对进入到淡水中盐度降低作出的适应性调节。半乳糖神经酰胺是一种强免疫兴奋剂,在许多体内实验模型中显示出强大的抗肿瘤活性[27],虹鳟 (Oncorhynchus mykiss) 在洄游产卵期会遇到海水中不同的病毒、细菌等进而增强体内免疫系统[28],本研究与此类似,刀鲚为应对环境改变所带来的免疫胁迫压力,也会通过增强特殊的免疫通路来完成洄游繁殖进程。
皮质醇是一类类固醇激素,属于糖皮质激素类,其通过糖异生作用提高血糖水平,也具有抑制免疫系统的功能,有助于脂肪、蛋白质和碳水化合物的代谢[29]。研究表明,在产卵季节,大麻哈鱼的血浆皮质醇水平会有升高现象[30]。皮质醇在薄唇鲻鱼 (Liza ramada) 卵巢卵黄形成早期呈下降趋势,在卵黄中期呈上升趋势,在卵黄后期下降,在产卵前期上升到较高水平,皮质醇的这种周期性变化支持了卵巢的发育和产卵[31]。类固醇代谢在卵巢发育中发挥着重要作用,其主要由内分泌腺如精巢、卵巢和肾上腺等单位合成之后释放到血液循环中,而所有的类固醇激素均由胆固醇合成,Xu等[18]研究指出,胆固醇合成途径在刀鲚卵巢发育过程中 (Ⅲ—Ⅴ期) 逐渐增强。本实验中皮质醇在淡水环境下比海水环境高1.61倍,每年2月初次性成熟的刀鲚开始从沿海区域进入长江等近海淡水区进行生殖洄游,在此过程中卵巢逐渐发育成熟,说明皮质醇在刀鲚洄游过程中参与了卵巢的发育过程,可能进一步促进了卵巢发育。此外,鱼体内的皮质醇具调节渗透压作用,能够刺激鱼类鳃上皮氯细胞密度增加,增强鳃内Na+/K+-ATPase活性,使过多的盐分排除体外[32-34],推测皮质醇在刀鲚的洄游过程中参与了渗透压的调节。
氨酰-tRNA合成酶在翻译中的作用是通过准确地将同源tRNA与相应的氨基酸配对来定义遗传密码,增强生物体在蛋白质翻译水平的准确性[35-36],本实验中下调的差异代谢物L-脯氨酸和L-异亮氨酸被富集到氨酰-tRNA生物合成代谢通路上,表明刀鲚卵巢组织为适应低盐度的环境可能相应地降低了蛋白质的翻译。同样下调表达的尿苷和假尿苷2个差异代谢产物富集到嘧啶代谢通路中,该通路的产物β-氨基丁酸 (β-aminobutyric acid) 可以参与到TCA循环中[37] ,表明淡水养殖环境下刀鲚卵巢组织内嘧啶类代谢受到部分抑制,甚至可影响能量代谢。刀鲚卵巢组织中鞘磷脂代谢在淡水环境中得到提升,被富集到该通路的半乳糖神经酰胺可稳固细胞膜脂质双分子层的机械稳定和化学抗性[38-39],由此可见,在刀鲚洄游过程中神经酰胺类代谢产物在信号转导和细胞识别中发挥重要作用。
4. 结论
本研究使用LC-MS非靶向代谢组学分析方法,研究了养殖在海、淡水中刀鲚的卵巢组织代谢产物的变化,共筛选得到47种差异代谢产物,推测碳环血氧烷A2和半乳糖神经酰胺的高表达,分别是为了更好地适应环境变化作出的适应性调节和增强免疫;淡水环境下皮质醇大量积累,推测皮质醇在刀鲚的洄游过程中参与了渗透压调节,同时可能与卵巢的发育相关;对差异代谢产物进行KEGG通路富集分析,筛选出氨酰-tRNA生物合成、嘧啶代谢和鞘磷脂代谢3条最显著的代谢途径,推测盐度的变化对刀鲚卵巢组织中的氨基酸、嘧啶、鞘磷脂的合成代谢有较大影响。
-
![]()
图 1 各季节优势物种结构聚类分析图
Figure 1. Clustering analysis of dominant species structure in each season
表 1 底层单拖网作业参数
Table 1 Parameter of bottom trawling
季节
season作业时间
time作业次数
number of net拖网时间/h
trawling time春季spring 2013年3月~4月 10 24 夏季summer 2013年6月~7月 9 9 秋季autumn 2013年9月~10月 5 7 冬季winter 2013年11月~12月 4 10  下载: 导出CSV
下载: 导出CSV
表 2 南沙西南陆架海域渔获物组成
Table 2 Species composition of catches in southwest shelf sea of Nansha area
鱼类fish 甲壳类crustacea 头足类cephalopoda 总种类
total species种类
species比例/%
percentage种类
species比例/%
percentage种类
species比例/%
percentage春季spring 68 77.53 8 8.99 12 13.48 89 夏季summer 122 85.31 12 8.39 9 6.29 143 秋季autumn 80 81.63 10 10.20 8 8.16 98 冬季winter 62 78.48 13 16.46 4 5.06 79
下载: 导出CSV
表 3 南沙西南陆架海域优势物种的优势度
Table 3 Dominance index of dominant species in southwest shelf sea of Nansha area
春季spring 夏季summer 秋季autumn 冬季winter 鱼种species IRI 鱼种species IRI 鱼种species IRI 鱼种species IRI 短尾大眼鲷
Priacanthus macracanthus6 181 短尾大眼鲷
Priacanthus macracanthus5 082 短尾大眼鲷
Priacanthus macracanthus7 130 短尾大眼鲷
Priacanthus macracanthus7 362 多齿蛇鲻
Saurida tumbil1 157 剑尖枪乌贼
Loligo edulis1 325 大头狗母鱼
Trachinocephalus myops1 210 蛇鲭
Gemphylus serpens1 601 剑尖枪乌贼
Loligo edulis940 红鳍圆鲹
Decapterus russelli319 鳞首方头鲳
Cubiceps squamiceps1 126 多齿蛇鲻
Saurida tumbil946 深水金线鱼
Nemipterus bathybius580 花斑蛇鲻
Saurida undosquamis263 长体圆鲹
Decapterus macrosoma1 070 无齿鲳
Ariomma brevimanus786 条尾绯鲤
Upeneus bensasi467 粗纹鲾
Leiognathus lineolatus216 黑 

Lophiomus setigerus758 箭天竺鲷
Rhabdamia gracilis571 黑
Lophiomus setigerus460 多齿蛇鲻
Saurida tumbil206 剑尖枪乌贼
Loligo edulis450 日本骨 
Ostichthys japonicus488 尖嘴 
Dasyatis zugei272 单棘躄鱼
Chaunax fimbriatus200 深水金线鱼
Nemipterus bathybius422 尖嘴
Dasyatis zugei469 大棘角鲂 
Pterygotrigla hemisticta230 黑
Lophiomus setigerus184 叉斑狗母鱼
Synodus macrops414 单棘躄鱼
Chaunax fimbriatus464 弓背鳄齿鱼
Champsodon atridorsalis200 带鱼
Trichiurus haumela172 鳃斑叉鼻鲀
Arothron immaculatus338 鳃斑叉鼻鲀
Arothron immaculatus420 青带小公鱼
Stolephorus zollingeri187 尖嘴
Dasyatis zugei141 武士 
Charybdis miles329 剑尖枪乌贼
Loligo edulis387 中国枪乌贼
Loligo chinensis163 杜氏枪乌贼
Loligo duvaucelii139 单棘躄鱼
Chaunax fimbriatus284 叉斑狗母鱼
Synodus macrops280 棘茄鱼
Halieutaea stellata162 日本骨
Ostichthys japonicus116 尖嘴
Dasyatis zugei147 黑
Lophiomus setigerus279 大头狗母鱼
Trachinocephalus myops147 中线天竺鲷
Apogon kiensis134 浅海长尾鲨
Alopias pelagicus275 长体圆鲹
Decapterus macrosoma117 蛇鲭
Gemphylus serpens111 深水金线鱼
Nemipterus bathybius239 带鱼
Trichiurus haumela110 海鳗
Muraenesox cinereus186 霜点石斑鱼
Epinephelus rivulatus181 弓背鳄齿鱼
Champsodon atridorsalis178 南洋羊舌鲆
Arnoglossus polyspilus137 鳒鲆
Psettina iijimae125 武士
Charybdis miles123 花斑蛇鲻
Saurida undosquamis119 红鳍圆鲹
Decapterus russelli112 黄鳍马面鲀
Navodon xanthopterus101
下载: 导出CSV
表 4 南沙西南陆架海域的物种生物多样性指数
Table 4 Biodiversity index of species in southwest shelf sea of the Nansha area
季节season 丰富度指数D 均匀度指数J′ 多样性指数H′ 春季spring 8.696 0.643 9 2.898 夏季summer 14.200 0.622 1 3.088 秋季autumn 10.400 0.576 6 2.650 冬季winter 7.834 0.633 5 2.768
下载: 导出CSV
-
[1] 贾晓平, 李纯厚, 陈作志, 等. 南海北部近海渔业资源及其生态系统水平管理策略[M]. 北京: 海洋出版社, 2012: 332. https://xueshu.baidu.com/usercenter/paper/show?paperid=3eaf8a22770735a48a6c2de01b4b369f&site=xueshu_se&hitarticle=1 [2] 陈作志, 邱永松. 南海区海洋渔业资源现状和可持续利用对策[J]. 湖北农学院学报, 2002, 22(6): 507-510. [3] 张鹏, 杨吝, 张旭丰, 等. 南海金枪鱼和鸢乌贼资源开发现状及前景[J]. 南方水产, 2010, 6(1): 68-74. doi: 10.3969/j.issn.1673-2227.2010.01.012 [4] 张俊, 陈作志, 陈国宝, 等. 南海鸢乌贼水声学测量和评估相关技术研究[J]. 南方水产科学, 2014, 10(6): 1-11. doi: 10.3969/j.issn.2095-0780.2014.06.001 [5] 晏磊, 张鹏, 杨吝, 等. 2011年春季南海中南部海域灯光罩网渔业渔获组成的初步分析[J]. 南方水产科学, 2014, 10(3): 97-103. doi: 10.3969/j.issn.2095-0780.2014.03.015 [6] 陈铮. 南沙群岛西南部陆架区底拖网渔业资源调查所获的鱼类[C]//中国科学院南沙综合考察队, 中国水产科学研究院南海水产研究所. 南沙群岛西南部陆架海区底拖网渔业资源调查研究专集. 北京: 海洋出版社, 1996: 38-52. [7] 李辉权. 南沙群岛西南部陆架海区主要经济鱼类生物学的初步研究[C]//中国科学院南沙综合考察队, 中国水产科学研究院南海水产研究所. 南沙群岛西南部陆架海区底拖网渔业资源调查研究专集. 北京: 海洋出版社, 1996: 53-57. CNKI:SUN:ZSCK.0.1996-01-006 [8] 詹海刚. 南沙群岛西南部陆架海区水文状况[C]//中国科学院南沙综合考察队, 中国水产科学研究院南海水产研究所. 南沙群岛西南部陆架海区底拖网渔业资源调查研究专集. 北京: 海洋出版社, 1996: 92-109. [9] 黄梓荣. 南海北部陆架区头足类的种类组成和资源密度分布[J]. 南方水产, 2008, 4(5): 1-7. doi: 10.3969/j.issn.2095-0780.2008.05.001 [10] 黄梓荣, 陈作志. 南沙群岛西南陆架区多齿蛇鲻的资源变动[J]. 南方水产, 2005, 1(3): 50-56. https://xueshu.baidu.com/usercenter/paper/show?paperid=1u750et0dp670p50qf740t50m3469739&site=xueshu_se&hitarticle=1 [11] PINKAS L, OLIPHANT M S, IVERSON I K. Food habits of albacore, blue fin tuna, and bonito in California waters[J]. Calif Dep Fish Game Fish Bull, 1971, 152(1): 1-105. https://xueshu.baidu.com/usercenter/paper/show?paperid=373e4f6fde34aa74f57ae687ba4b7b50
[12] 陈国宝, 李永振, 陈新军. 南海主要珊瑚礁水域的鱼类物种多样性研究[J]. 生物多样性, 2007, 15(4): 373 -381. doi: 10.3321/j.issn:1005-0094.2007.04.006 [13] MARGALSE R. Information theory in ecology[J]. Gen Sys, 1958(3): 36-71. https://xueshu.baidu.com/usercenter/paper/show?paperid=273dd7a6b376ce8fbc6645d9d4459561
[14] KREBS C J. Ecological methodology[M]. New York: Harper Collins Publishers, 1989: 478-505. doi: 10.1007/978-94-009-4115-1_3
[15] SIMPSON E H. Measurement of diversity[J]. Nature, 1949(163): 688. doi: 10.1136/thx.27.2.261
[16] PIELOU E C. The use of information theory in the study of ecological succession[J]. J Theor Biol, 1966(10): 370-383. doi: 10.1111/brv.12995
[17] 江艳娥, 林昭进, 黄梓荣. 南海北部大陆架区渔业生物多样性研究[J]. 南方水产, 2009, 5(5): 33-37. doi: 10.3969/j.issn.1673-2227.2009.05.006 [18] 张鹏, 曾晓光, 杨吝, 等. 南海区大型罩网渔场渔期和渔获组成分析[J]. 南方水产科学, 2013, 9(3): 74-79. doi: 10.3969/j.issn.2095-0780.2013.03.012 [19] 邹建伟, 林丕文, 王强哲. 南海北部湾2012年捕捞产量估算[J]. 南方水产科学, 2013, 9(4): 76-82. [20] 费鸿年, 何宝全, 陈国铭. 南海北部大陆架底栖鱼群聚的多样度以及优势种区域和季节变化[J]. 水产学报, 1981, 5(1): 1-20. https://xueshu.baidu.com/usercenter/paper/show?paperid=08e614d849bd499745607be0e6bc9591&site=baike [21] MARGALSE R. Perspectives in ecological theory[M]. Chicago: University of Chicago Press, 1968: 77-83. doi: 10.1515/9781400860180
[22] HAEDRICH R L, HAEDRICH S O. A seasonal survey of the fishes in the Mystic River, a polluted estuary in downtown Boston, Massachusetts[J]. Est Coast Mar Sci, 1974(2): 59-73. doi: 10.1016/0302-3524(74)90028-0
[23] 曾晓光, 李娜娜, 杨权, 等. 南沙群岛西南部陆架海域鱼类分类的多样性[J]. 水产学报, 2012, 36(4): 592-600. [24] 杜飞雁, 王雪辉, 李纯厚, 等. 大亚湾大型底栖动物物种多样性现状[J]. 南方水产, 2008, 4(6): 33-41. doi: 10.3969/j.issn.2095-0780.2008.06.005 [25] 颜云榕, 王峰, 郭晓云, 等. 基于3S集成平台的南海渔业信息动态采集与实时自动分析系统研发[J]. 水产学报, 2014, 38(5): 748-758. doi: 10.3724/SP.J.1231.2014.48926 -
期刊类型引用(6)
1. 王子涵,曾聪,姜子禺,曹玲. 东海及其邻近海域受胁鱼类保护空缺分析. 热带海洋学报. 2023(01): 66-86 .  百度学术
百度学术
2. 王新月,陈生熬,王程欣,訾方泽,常德胜,许豪,李大鹏. 叶尔羌高原鳅耳石形态探究及群体判别分析. 渔业科学进展. 2023(04): 201-211 . 百度学术
3. 谢玉,玄文丹,周永东,朱凯,朱文斌,王忠明,徐开达,王嘉浩. 东海海域带鱼矢耳石形态特征与生长研究. 浙江海洋大学学报(自然科学版). 2023(04): 294-302 . 百度学术
4. 潘梦珍,王海山,叶乐,陈治,杨超杰. 崖州湾附近海域金线鱼形态学指标的相关性. 海南热带海洋学院学报. 2020(02): 11-17 . 百度学术
5. 蔡研聪,黄梓荣,李佳俊,许友伟,孙铭帅,陈作志,刘维达. 南海北部近海新记录种——苏门答腊金线鱼资源分布特征. 南方水产科学. 2020(06): 1-11 . 本站查看
6. 杨超杰,唐婷婷,吴东霞,杜良玉,方馨正. 崖州湾附近海域金线鱼繁殖生物学研究. 海南热带海洋学院学报. 2019(05): 6-11 . 百度学术
其他类型引用(5)





计量
- 文章访问数: 3135
- HTML全文浏览量: 176
- PDF下载量: 744
- 被引次数: 11
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
