
A study on sinking and closing performance of falling-net
-
摘要:
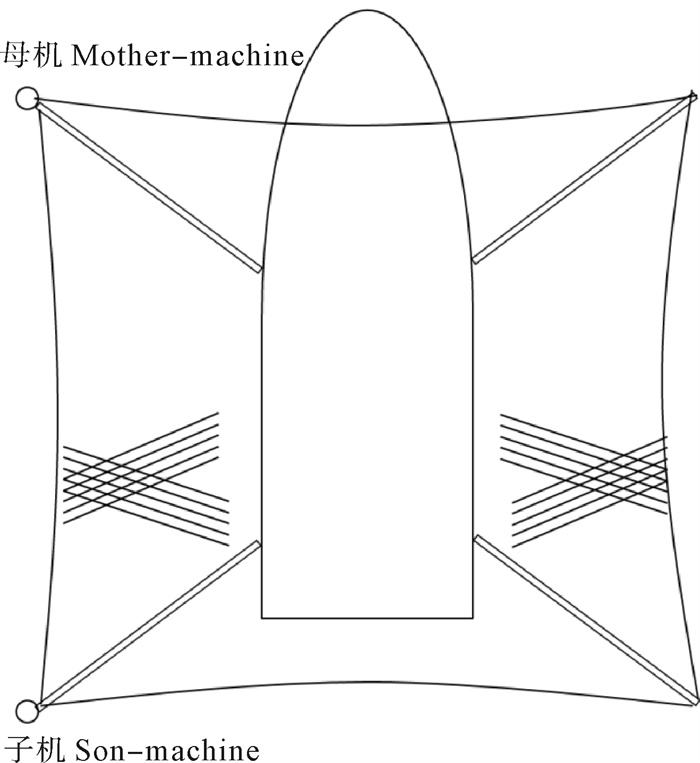
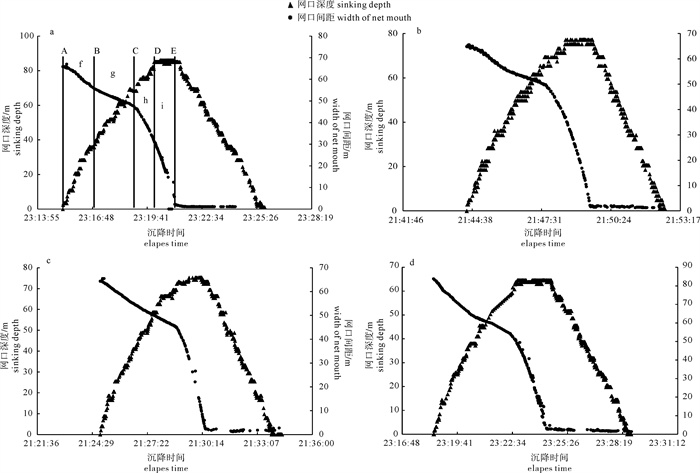
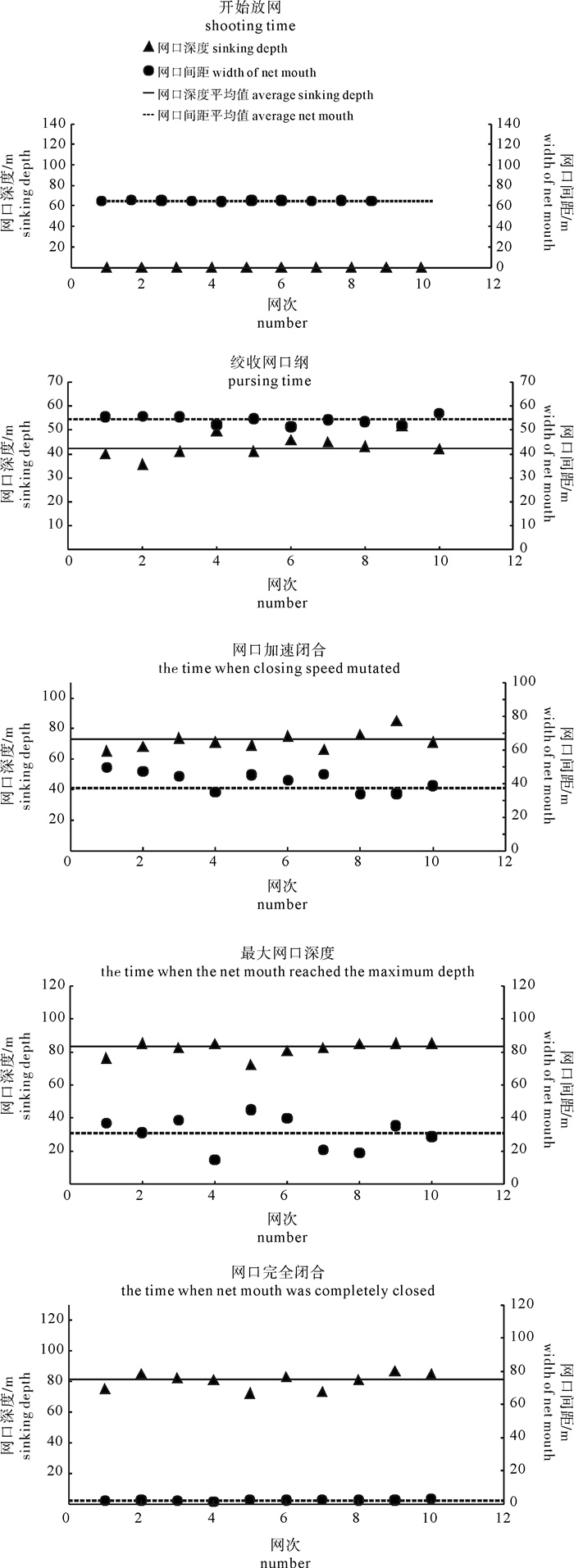
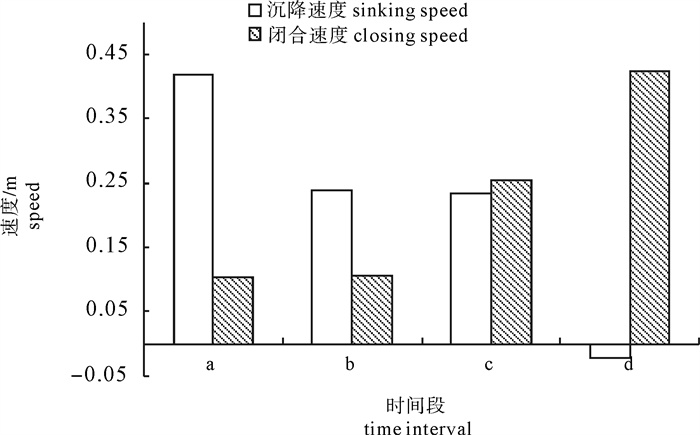
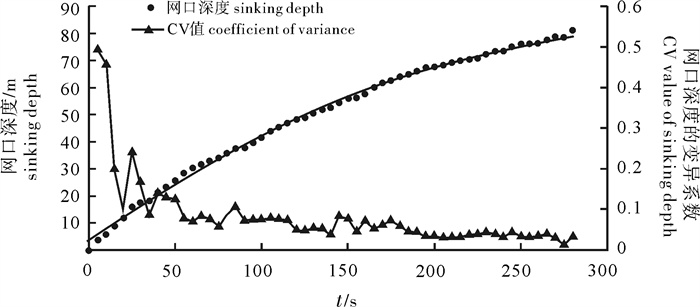
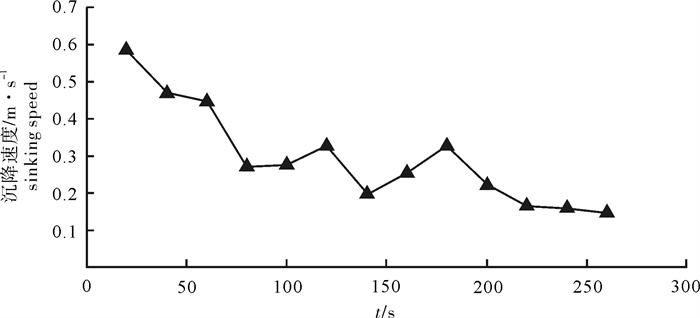
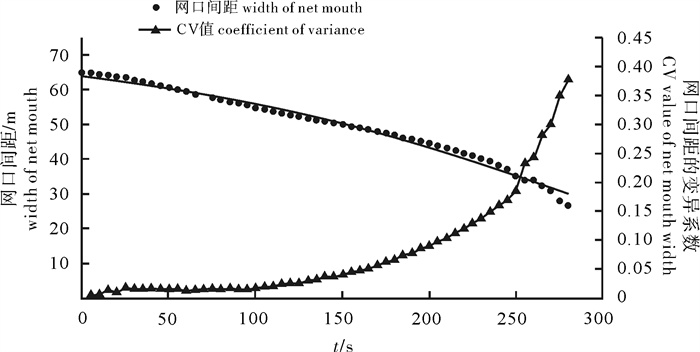
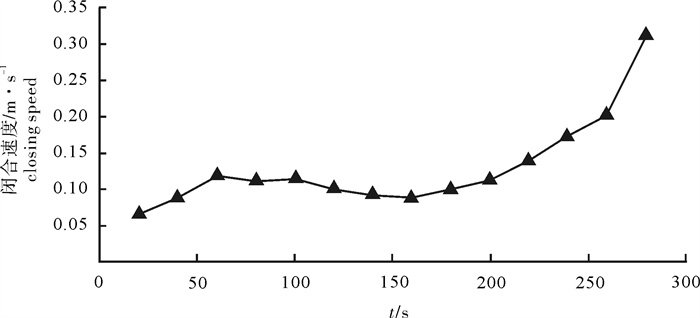
根据2014年11月使用SDKN-500型网位仪所采集的罩网作业时网口深度和间距的实时数据,分析罩网网口沉降与闭合性能。结果发现:1)渔船绞收网口纲后,网口沉降和闭合速度不会马上突变,存在120.7 s的滞后时间;2)网口降至最大深度时(平均82.0 m),仍未完全闭合,此后深度相对稳定,直至网口完全闭合(平均68.0 s)并停滞一段时间(平均9.8 s)后网口才开始上升;3)渔船不同作业状态可分为开始放网(设为0 s)、开始绞收网口纲(平均104.0 s)、网口加速闭合(平均224.7 s)、网口达最大深度(平均266.6 s)、网口完全闭合(平均334.6 s),也是决定罩网性能的5个关键时间节点;4)4个时段的网口沉降速度分别为0.419 m · s-1、0.238 m · s-1、0.233 m · s-1和-0.021 m · s-1,总体呈递减趋势,沉降中期存在波动;5)4个时段的网口闭合速度分别为0.103 m · s-1、0.105 m · s-1、0.253 m · s-1和0.424 m · s-1,总体呈递增趋势;整个沉降过程闭合速度在前期先变大后变小,在沉降后期明显加大。建议提高网口纲绞收速度以避免鱼群逃逸。
Abstract:Based on the real-time data of sinking depth and distance of net mouth of falling-net by SDKN-500 net monitor in November 2014, we analyzed the sinking and closing performance of falling-net. The results show that: 1) The sinking and closing speeds would not mutate when pursing the purse line with retardation time of 120.7s;2) The net mouth was not completely closed when it reached the maximum depth (averagely 82.0 m), and did not rise up until it was completely closed (averagely 68.0 s) and stayed for a while (averagely 9.8 s); 3) The different operating conditions could be divided into the shooting time (0 s), the pursing time (averagely 104.0 s), the time when the closing speed mutated (averagely 224.7 s), the time when the net mouth reached the maximum depth (averagely 266.6 s) and the time when the net mouth was completely closed (average 334.6 s) in sequence, which were the five key time nodes determining the fishing performance of falling-net; 4) The sinking speeds of the four periods were 0.419 m · s-1, 0.238 m · s-1, 0.233 m · s-1 and -0.021 m · s-1, respectively, showing a decreasing trend with fluctuation in the middle of settlement; 5) The closing speeds of the four periods were 0.103 m · s-1, 0.105 m · s-1, 0.253 m · s-1 and 0.424 m · s-1, respectively, showing an increasing trend. The closing speed of net mouth increased firstly and then decreased at the beginning of settlement, and the closing speed showed a significant increasing trend in the later period of settlement. It is suggested to increase the hauling speed to avoid fish escape.
-
Keywords:
- falling-net /
- sinking performance /
- closing performance of net mouth
-
长体圆鲹(Decapterus macrosoma),又名长身圆鲹,隶属于鲈形目、鲹科、圆鲹属,主要分布于中国南海、印度尼西亚、澳洲和日本南部沿海等地[1]。长体圆鲹在中国南海分布较广,是南海灯光围网渔业的主要捕捞对象,具有较高的经济价值[2-5]。目前,国内外学者关于长体圆鲹的研究主要集中在生长繁殖[6]和资源评估[7-8]方面,与种群遗传和分子标记相关的研究报道较少。微卫星标记仅见翟云等[9]开发蓝圆鲹微卫星标记中获得5个跨物种通用标记可于用长体圆鲹,并无专门针对长体圆鲹开发的微卫星标记。种群遗传信息的匮乏,将大大影响对其资源的评估和长期有效的管理。
微卫星分子标记因是共显性标记,具有多态性高、变异性强、数据易统计等突出优点[10],广泛应用于海洋生物遗传结构及遗传多样性分析[11-12]。但由于微卫星标记通用性较差,常常具有极强的种属特异性。鱼类微卫星标记开发中多以二核苷酸重复为主[13-15],普遍认为它们具有较高的遗传变异[16],但是也有部分学者研究认为三、四核苷酸重复位点较二核苷酸重复具有更高的筛选效率和多态性[17-19]。
本研究通过RAD-Seq高通量测序方法开发长体圆鲹二、三核苷酸微卫星分子标记,并对测试群体进行多样性分析,同时比较二、三核苷酸的筛选效率和多态性差异,旨在为长体圆鲹种群遗传结构及遗传多样性分析提供技术基础,并为该资源的评估和管理提供帮助。
1. 材料与方法
1.1 样品采集与基因组DNA提取
长体圆鲹样品采集于中国南海中沙群岛东部海域,共35尾。剪取部分肌肉样品加入无水乙醇保存。每个样品剪取少量肌肉组织,使用海洋动物组织基因组DNA提取试剂盒(天根,北京) 提取基因组DNA,0.8%的琼脂糖凝胶电泳检测DNA提取质量,−20 ℃保存备用。
1.2 高通量测序与引物合成
使用HiSeq2000高通量测序仪(Illumina,USA) 对长体圆鲹基因组DNA进行RAD-seq (测序服务由广州基迪奥生物科技有限公司提供),经生物信息学搜索出微卫星位点[20]。使用Premier 5.0软件在重复单元侧翼序列上选择性设计出112条引物,主要参数为:G-C含量为40%~60%,引物长度为18~25 bp,退火温度为45~60 ℃,预期产物长度180~320 bp。送上海英潍捷基贸易有限公司合成引物。
1.3 引物筛选与分型检测
选取3个样本混合成的基因组DNA为模板,优化PCR反应条件,对引物进行首轮筛选,琼脂糖电泳检测是否能扩增出稳定且均一的目的片段。之后选取8尾个体的基因组DNA作为模板,使用三引物法[21],利用M13荧光接头引物进行PCR扩增,扩增产物送华大基因公司经毛细管电泳进行等位基因分型,检测引物是否具有多态性。PCR反应体系为15 μL,其中包括10×PCR Buffer 1.5 μL,2.5 mmol·L–1 MgCL2 1.2 μL,2 mmol·L–1 dNTPs 2 μL,M13正向引物(10 μmol·L–1) 0.2 μL,M13反向引物(10 μmol·L–1) 0.6 μL,M13通用荧光引物(10 μmol·L–1) 0.5 μL,Taq酶(5 U·μL–1) 0.15 μL,DNA模版1 μL,加双蒸水至15 μL。PCR扩增程序为:94 ℃预变性5 min;94 ℃变性45 s,55~60 ℃退火45 s,72 ℃延伸45 s,35个循环;94 ℃变性45 s,53 ℃退火45 s,72 ℃延伸45 s,8个循环;72 ℃延伸30 min。
1.4 长体圆鲹群体遗传学评价
使用35尾长体圆鲹个体的基因组DNA为模板,对通过筛选的微卫星标记的种群遗传学特征进行评价。PCR反应体系和条件、等位基因分型方法如上。使用软件Genepop 4.0[22]对每个标记的种群遗传学特征值进行计算,包括等位基因数(Na)、表观杂合度(Ho)和期望杂合度(He),进行“哈迪-温伯格”平衡(HWE)检验和连锁不平衡检测,并对P值进行Bonferroni校正。使用Cervus 3.0.7[23]软件计算多态信息含量(PIC)。
2. 结果
2.1 高通量测序结果与微卫星位点分析
RAD-seq高通量测序共获得长体圆鲹基因组原始数据2.06 G,GC含量为41.43%,Q30达93.05%。说明测序结果质量较好,可用于后续分析。搜索后共获得微卫星序列58 180条,一至六核苷酸重复微卫星位点70 508个,其中二核苷酸重复微卫星位点最多(37 646个),占总数的53.39% (表1),说明二核苷酸重复为主要的微卫星类型。二核苷酸重复微卫星位点共有4种重复类型,4种类型重复微卫星数量相差较大,AC/GT类有29 754个,占二核苷酸重复的68.4%;AG/CT类有6 487个,占17.2%;AT/TA有1 340个,占3.6%;GC/CG仅有65个,占0.17%。
表 1 长体圆鲹基因组中不同类型SSR统计Table 1. Different types of SSR statistics in D. macrosoma genome重复单元
repeat unit微卫星数量/个
number of microsatellite占比/%
ratio一核苷酸 mono-nucleotide 8 184 11.61 二核苷酸 di-nucleotide 37 646 53.39 三核苷酸 tri-nucleotide 13 960 19.80 四核苷酸 tetra-nucleotide 7 741 10.98 五核苷酸 penta-nucleotide 2 255 3.20 六核苷酸 hexa-nucleotide 722 1.02 合计 total 70 508 100.00 2.2 PCR引物设计和筛选
选取112条二、三核苷酸重复序列设计引物,其中二核苷酸重复为81对,三核苷酸重复为31对。经过筛选后,共有27对引物通过筛选(表2),27对引物扩增的序列中18个位点为二核苷酸重复,重复次数为9~14次;8个位点为三核苷酸重复,重复次数为6~10次。二核苷酸重复位点检出效率为22.2%,三核苷酸重复位点检出效率为25.8%。
表 2 27对长体圆鲹微卫星引物信息Table 2. Information of 27 pairs of primers in D. macrosoma位点
locus引物序列 (5'−3')
primer sequence重复单元
repeat motif退火温度/℃
annealing temperature期望长度/bp
allele sizeDma03 F:CCACGCCTATTGAGTTACAGA (CA)9 60 186 R:GAGCCAGTGGATGAACAGAGT Dma07 F:GCCCCTGTGGGTGTGTGA (CA)9 60 225 R:GGGTGGTGGGTTCGGTTT Dma12 F:GAACCAGTGCCTACAATAGA (AC)9 60 243 R:CTGCTCACGGTAAGTCCA Dma15 F:ACAGGAAGGAACAGGACAG (TG)10 55 254 R:TATTGAAGTGAAAAAGCCG Dma22 F:CGCTGTTGAAATGAAGAAGA (GT)10 60 317 R:AGTGATGTCGCCTCATAAAT Dma23 F:AAACTGAGGGCGAGATAGAGG (AC)10 55 190 R:CCACAGGCTGAGTAAACCAAC Dma26 F:ATCCCATTCACCGACATAG (TG)10 58 258 R:CTGTGGTATCGTTCCCTGT Dma28 F:TGATTGGCTTCTACTCTGC (AC)10 55 281 R:AGTGGCTTGTTTGACTCTTAT Dma36 F:GGATGTAGTGAAGAGGGGAG (GT)11 55 239 R:CACAATCAGTGTTATGGCAG Dma38 F:GCCAATAAAGGCAAACAGT (CA)11 60 227 R:ATCCGAGACAAAGACATACAA Dma39 F:AGTGTGCTGACTTTTCTCTG (CA)11 55 241 R:TTATTGTTTGTTGTCTGGGT Dma45 F:CTCCTTTTTCTTCTTCCTCT (CA)11 60 281 R:CTACCTGCTCTTCAACTCAT Dma51 F:TGACAGCCTCCACTACTCC (GA)12 55 225 R:GCTAACCAGACACGCAAA Dma54 F:AAAGCCCATCTGTCTCGT (GT)12 60 202 R:TGTTTCAGTCCGTTCCTG Dma58 F:TCAAGAGGGAGTGGGAGC (AC)12 58 279 R:TCAAATGGGTGTTTAGCG Dma64 F:GCTCAGACTGCGTGGACA (TG)13 55 314 R:GCTGGTGAACAACAGGACA Dma72 F:TTCCGCAGGCATAAAAAC (CT)13 58 301 R:CCAAGGTCCGCTACACTA Dma76 F:TTCTCGCTGACCTGCTTG (TG)14 55 253 R:GCGTCCTCGTCGTCTTTC Dma81 F:GAGACACGGTCAGAAAACA (TGC)6 60 216 R:GGAAGTAGGACTCTAGGGG Dma82 F:CTGTCACTCCATTCCTATTCC (GTT)6 58 244 R:CCTACATTTGTGCTTTTGTTC Dma83 F:CTCTAAAGCCGACCTAACC (CTT)6 58 239 R:TGTCTCAACACAGCGAAAC Dma84 F:AAACTAACTCATCACCAG (TGT)6 55 283 R:AAACGACAGGAACTCAAT Dma85 F:CTCACTTTGACCCAACCAG (AGG)6 55 256 R:CCTTTCACCGAGACACCAG Dma131 F:TGCGGATGGGTGGTAGTGT (GGT)8 55 208 R:ATTGCTGGTAGTCGGTGGC Dma132 F:CCCAGTGAGACCAGAACCA (GCT)8 55 268 R:GACCCGTAGACAGGAGAGT Dma135 F:GTTGTTGTTTTTTTCCTT (GCA)9 55 301 R:CATCAGTCTGGCTTTATA Dma145 F:ACGATACAGCAGCCGAAG (TCA)10 60 197 R:AGTGATGTCGCCTCATAAAT 2.3 微卫星标记的种群遗传学评价
使用1个采集自南海东南部海域的长体圆鲹群体对筛选合格的微卫星标记进行种群遗传学评价。所有27个标记在测试群体中共检测到285个等位基因,等位基因数为5~17,Ho为0.342 9~0.857 1,平均为0.631 7;He为0.538 3~0.911 8,平均为0.7968。PIC为0.497~0.886,平均为0.780 9 (表3),表明开发的微卫星位点具有较高的多态性。共有19个标记等位基因频率符合“哈迪-温伯格”平衡。连锁不平衡检测表明各位点间无连锁不平衡现象。
表 3 长体圆鲹微卫星标记的种群遗传学特征Table 3. Characteristics of microsatellite loci in D. macrosoma位点
locusN Na Ho He PHWE PIC Dma03 35 8 0.857 1 0.790 9 0.042 0 0.746 0 Dma07 35 9 0.771 4 0.855 9 0.553 4 0.825 0 Dma12 34 11 0.685 7 0.816 6 0.106 4 0.809 0 Dma15* 35 14 0.542 9 0.911 8 0.000 0 0.890 0 Dma22* 32 14 0.485 7 0.813 2 0.000 0 0.864 0 Dma23 34 17 0.714 3 0.864 8 0.003 3 0.868 0 Dma26* 34 14 0.428 6 0.869 1 0.000 0 0.870 0 Dma28 35 11 0.742 9 0.837 7 0.035 3 0.803 0 Dma36 35 9 0.685 7 0.786 3 0.054 5 0.743 0 Dma38* 34 12 0.628 6 0.851 2 0.000 0 0.849 0 Dma39* 33 12 0.514 3 0.782 0 0.000 0 0.793 0 Dma45 34 12 0.771 4 0.847 8 0.139 2 0.844 0 Dma51* 31 12 0.485 7 0.784 5 0.000 0 0.859 0 Dma54 35 15 0.771 4 0.900 6 0.139 0 0.878 0 Dma58 34 12 0.771 4 0.845 6 0.449 7 0.843 0 Dma64* 31 13 0.428 6 0.788 3 0.000 0 0.865 0 Dma72 35 8 0.628 6 0.713 9 0.155 7 0.659 0 Dma76 35 12 0.685 7 0.908 1 0.003 6 0.886 0 Dma81 35 8 0.628 6 0.795 0 0.045 7 0.752 0 Dma82 35 5 0.485 7 0.538 3 0.023 6 0.497 0 Dma83* 34 8 0.342 9 0.705 8 0.000 0 0.674 0 Dma84 35 7 0.542 9 0.704 8 0.005 9 0.649 0 Dma85 35 8 0.828 6 0.747 8 0.876 9 0.704 0 Dma131 35 9 0.628 6 0.717 6 0.646 6 0.661 0 Dma132 35 8 0.657 1 0.746 6 0.365 5 0.692 0 Dma135 34 10 0.742 9 0.840 5 0.241 8 0.835 0 Dma145 34 7 0.600 0 0.749 3 0.077 4 0.725 0 注:N. 有效样品数;Na. 等位基因数;Ho. 表观杂合度;He. 期望杂合度;PHWE. “哈迪-温伯格”平衡显著性检验P值;PIC. 多态信息含量;*. 经Bonferroni校正后显著背离“哈迪-温伯格”平衡 (校正P<0.001 85) Note: N. effective number of samples; Na. number of alleles; Ho. observed heterozygosity; He. expected heterozygosity; PHWE. Hardy–Weinberg probability test; PIC. polymorphism information content; *. significant deviation from HWE after Bonferroni's correction (adjusted P-value<0.001 85) 3. 讨论
3.1 高通量测序发掘微卫星序列的技术优势
传统微卫星标记开发方法耗时长、花费高、技术难度大。以磁珠富集法为例,标记开发过程中基因组DNA浓度、接头连接效率、富集过程中的杂交温度以及洗脱条件的控制等因素都会影响微卫星筛选的效率[24-25],且最终获得的有效微卫星序列仅几百条[26-27]。相比较而言,高通量测序技术开发微卫星标记,省略了建库、克隆、筛选等,只需提取基因组DNA测序,利用生物信息学手段可直接获取微卫星序列,通常是传统方法获得微卫星序列数目的几百倍[15,28-29],具有高效、便捷、准确的特点,能够满足短时间内大批量微卫星位点的开发需求,比如连锁图谱构建、QTL定位等[30-31]。
本次RAD-seq高通量测序共获得长体圆鲹基因组原始数据2.06 G,GC含量为41.43%,测序质量Q30达93.05%;共获得微卫星序列58 180条,一至六核苷酸重复微卫星位点70 508个。说明测序质量稳定高效,并获得了数量庞大、类型丰富的长体圆鲹微卫星序列,可用于后续长体圆鲹微卫星标记的大规模开发和相关遗传学研究。
3.2 不同核苷酸重复微卫星位点比较
本次高通量测序结果表明在长体圆鲹微卫星位点中二核苷酸重复为主要重复类型,AC/GT类重复数量最为丰富,GC/CG重复较为少见。此结果与大量水产动物微卫星位点研究结果相一致[32-34],差异仅在于比例多少,以及除二核苷酸重复占主要地位外其他核苷酸重复的含量差异。熊良伟等[33]对中华鳑鲏(Rhodeus sinensis)微卫星的分析中,二核苷酸占总微卫星位点的53.59%,其次为单核苷酸重复,二核苷酸重复中AC/GT类占60.63%,GC/CG仅占0.32%。在裸体异鳔鳅鮀 (Xenophysogobio nudicorpa)中[32],二核苷酸重复占总微卫星位点比例高达83.15%,AC/GT类重复占49.36%,GC/CG重复仅有4个。
多数鱼类开发的微卫星标记以二核苷酸重复为主,但研究表明,在人类基因组中三核苷酸重复序列与遗传疾病的发生有关,并且具有较高的多态性和遗传稳定性[35]。部分学者对三、四核苷酸重复微卫星标记的研究结果存在差异。房祖业等[28]对大刺鳅 (Mastacembelus armatus) 二、三、四核苷酸重复微卫星标记的筛选发现二核苷酸重复较三、四核苷酸重复具有更高的筛选效率和多态性;鲁翠云等[17]、谭照君等[18]、李文升等[19]的研究认为三、四核苷酸具有更高的多态性和分型效果。长体圆鲹二、三核苷酸的筛选效率分别为22.2%和29.0%,PIC分别为0.827 4和0.687 7 (表4)。就筛选效率而言,三核苷酸重复略高于二核苷酸重复,但二者相差不大。PIC为衡量种群遗传变异程度的重要指标[36],二核苷酸重复多态性明显高于三核苷酸重复。本文中长体圆鲹二核苷酸重复筛选效率低于三核苷酸重复,但多态性二核苷酸重复明显高于三核苷酸重复。因此,筛选效率和多态性的差异可能由种属差异或其他多种因素导致。
表 4 长体圆鲹二、三核苷酸重复微卫星标记的比较Table 4. Comparison on di- and trinucleotide-repeated microsatellite loci in D. macrosoma序列
sequence引物数
primer number重复次数
repeat number筛选效率
efficiencyPIC 二核苷酸重复
di-nucleotide-repeated18 9~14 22.2% 0.827 4 三核苷酸重复
tri-nucleotide-repeated9 6~10 29.0% 0.687 7 通过筛选的27对引物中18个位点为二核苷酸重复,重复次数为9~14次不等;9个位点为三核苷酸重复,重复次数为6~10次不等,符合Ellegren[37]提出的真核生物微卫星位点重复大部分在30次重复以下。但Ellegren[37]认为二核苷酸重复以15~19次为主,本文中高通量测序获得的二核苷酸重复主要在6~15次。基于Weber[38]的研究结果,重复次数高的微卫星在种群中表现出的多态性较高,龚小玲等[39]对澳洲鳗鲡 (Anguilla australis) 进行标记开发时发现,微卫星重复序列的重复次数过高会影响PCR效果,应选择居中的重复次数为宜。长体圆鲹二核苷酸重复PIC为0.827 4,具有较高多态性,表明选择6~15次的二核苷酸重复是合适的。
3.3 微卫星标记的种群遗传学特征
群体杂合度的高低反映了群体在多个基因座上的遗传变异及群体遗传多样性丰富度[19]。本研究中长体圆鲹中沙群体的平均Ho为0.631 7,平均He为0.796 8,说明长体圆鲹该群体的遗传多样性较高。平均Ho和He存在差异,说明存在杂合子缺失或者纯合子过剩的情况。PIC也是衡量群体遗传多样性的重要指数,Botstein等[36]认为基因标记PIC>0.5为高度多态位点,0.25<PIC<0.5为中度多态位点,PIC<0.25为低度多态性位点,通常不作为遗传多样性分析。本文中长体圆鲹位点除1个为中度多态外,其他位点均为高度多态位点。表明开发所得的长体圆鲹微卫星标记在中沙群体中具有较好的遗传稳定性和丰富的遗传多样性。
在所有27个位点中有8个位点偏离了“哈迪-温伯格”平衡,这些位点不适合进一步的遗传分析。近亲杂交、无效等位基因、种群退化和自然选择等因素皆可能导致微卫星位点偏离HWE[15]。
-
![]()
图 5 4个时间段网口沉降与闭合速度
Figure 5. Sinking and closing speed of net mouth at four time intervals
-
[1] 杨吝, 卢伙胜, 吴壮, 等. 南海区海洋渔具渔法[M]. 广州: 广东科技出版社, 2002: 164-168. [2] 张鹏, 杨吝, 张旭丰, 等. 南海金枪鱼和鸢乌贼资源开发现状及前景[J]. 南方水产, 2010, 6(1): 68-73. doi: 10.3969/j.issn.1673-2227.2010.01.012 [3] 晏磊, 张鹏, 杨吝, 等. 2011年春季南海中南部海域灯光罩网渔业渔获组成的初步分析[J]. 南方水产科学, 2014, 10(3): 97-103. doi: 10.3969/j.issn.2095-0780.2014.03.015 [4] 张鹏, 曾晓光, 杨吝, 等. 南海区大型罩网渔场渔期和渔获组成分析[J]. 南方水产科学, 2013, 9(3): 74-79. doi: 10.3969/j.issn.2095-0780.2013.03.012 [5] 邱永松, 张鹏. 南海大洋性渔业资源开发利用对策建议[C]. 热带海洋科学学术研讨会暨第八届广东海洋湖沼学会、第七届广东海洋学会会员代表大会论文及摘要汇编. 广州: 广东省科学技术协会科技交流部, 2013: 199-203. https://cpfd.cnki.com.cn/Article/CPFDTOTAL-GDKL201305001026.htm [6] 杨吝, 张旭丰, 张鹏, 等. 南海区海洋小型渔具渔法[M]. 广州: 广东科技出版社, 2007: 103-115. [7] 杨吝, 张旭丰, 谭永光, 等. 南海北部灯光罩网渔获组成及其对渔业资源的影响[J]. 南方水产, 2009, 5(4): 41-46. doi: 10.3969/j.issn.1673-2227.2009.04.008 [8] 邹建伟, 陈立峰, 林蒋进. 南海外海灯光罩网主要渔场分布及变动研究——基于广西渔船的生产监测统计[J]. 南方水产科学, 2014, 10(4): 78-84. doi: 10.3969/j.issn.2095-0780.2014.04.013 [9] 颜云榕, 冯波, 卢伙胜. 中、西沙海域2种灯光作业渔船的捕捞特性及其技术效率分析[J]. 南方水产, 2009, 5(6): 59-64. doi: 10.3969/j.issn.1673-2227.2009.06.011 [10] 冯波, 许永雄, 卢伙胜. 南沙北部灯光罩网与金枪鱼延绳钓联合探捕[J]. 广东海洋大学学报, 2012, 32(4): 54-58. https://xueshu.baidu.com/usercenter/paper/show?paperid=42230c90c74c793caed758cd81c178fc [11] 杨权, 李永振, 张鹏, 等. 基于灯光罩网法的南海鸢乌贼声学评估技术研究[J]. 水产学报, 2013, 37(7): 1032-1039. doi: 10.3724/SP.J.1231.2013.38513 [12] 晏磊, 张鹏, 杨吝, 等. 南海灯光罩网沉降性能研究[J]. 上海海洋大学学报, 2014, 23(1): 146-153. https://xueshu.baidu.com/usercenter/paper/show?paperid=7c3cde045035a7ed323dc0ccfb4eb144&site=xueshu_se&hitarticle=1 [13] 冯维山. 围网下纲沉降特性试验研究[J]. 大连水产学院学报, 1990, 5(3): 37-43. https://xueshu.baidu.com/usercenter/paper/show?paperid=c20be56bccf9b5a4fdfab7eca99d5b04&site=xueshu_se&hitarticle=1 [14] MISUND O, DICKSON W, BELTESTAD A. Optimization of purse seines by large-meshed sections and low lead weight. Theoretical considerations, sinking speed measurements and fishing trials[J]. Fish Res, 1992, 14(4): 305-317. doi: 10.1016/0165-7836(92)90039-V
[15] 冯维山. 围网下纲提升特性研究[J]. 大连水产学院学报, 1993, 8(1): 29-36. https://xueshu.baidu.com/usercenter/paper/show?paperid=25f890fd8bc6b390b20042f81417a5c6&site=xueshu_se [16] SUK J K, JEONG S P. An analysis on the sinking characteristics of model purse seine by different netting material[J]. Kor Soc Fish Tech, 1995, 31(4): 372-379.
[17] SUK J K, JEONG S P. An analysis of sinking resistance for purse seine[J]. Kor Soc Fish Tech, 1998, 34(3): 275-234.
[18] KIM Y H. Geometry of the model purse seine in relation to enclosed volume during hauling operation[J]. Fish Aquat Sci, 2000, 3(2): 156-162. https://www.semanticscholar.org/paper/Geometry-of-the-Model-Purse-Seine-in-Relation-to-Yong-Hae/90875fbfcc024cfe61178f0c75a33f86916920e4
[19] HYUN Y K, CHUN W L, JONG K S, et al. Dynamic simulation of the behavior of purse seine gear and sea-trial verification[J]. Fish Res, 2007, 88(1/2/3): 109-119. doi: 10.1016/j.fishres.2007.08.007
[20] 王春雷. 中西太平洋1664.5m×394.3m金枪鱼围网沉降性能研究[D]. 上海: 上海海洋大学, 2008. 10.7666/d.y1660028 [21] YONG H K, MYEONG C P. The simulation of the geometry of a tuna purse seine under current and drift of purse seiner[J]. Ocean Eng, 2009, 36(14): 1080-1088. doi: 10.1016/j.oceaneng.2009.06.011
[22] HOSSEINI S A, LEE C W, KIM H S, et al. The sinking performance of the tuna purse seine gear with large-meshed panels using numerical method[J]. Fish Sci, 2011, 77(4): 503-520. doi: 10.1007/s12562-011-0371-6
[23] 兰光查. 基于模型试验的金枪鱼围网沉降性[D]. 上海: 上海海洋大学, 2011. [24] 许柳雄, 王敏法, 叶旭昌, 等. 金枪鱼围网沉降特性[J]. 中国水产科学, 2011, 18(5): 1161-1169. doi: 10.3724/SP.J.1118.2011.01161 [25] 许柳雄, 兰光查, 叶旭昌, 等. 下纲重量和放网速度对金枪鱼围网下纲沉降速度的影响[J]. 水产学报, 2011, 35(10): 1563-1569. doi: 10.3724/SP.J.1231.2011.17228 [26] 周成, 许柳雄, 张新峰, 等. 金枪鱼围网沉降性能影响因子的多元回归分析[J]. 中国水产科学, 2013, 20(3): 672-681. http://www.fishscichina.com/zgsckx/article/abstract/5178 [27] 唐浩, 许柳雄, 周成, 等. 基于GAM模型研究金枪鱼围网沉降性能影响因素[J]. 水产学报, 2013, 37(6): 944-950. doi: 10.3724/SP.J.1231.2013.38336 [28] 唐浩, 许柳雄, 王学昉, 等. 金枪鱼围网模型试验结果与海上实测的比较评估[J]. 中国水产科学, 2013, 20(4): 884- 892. doi: 10.3724/SP.J.1118.2013.00884 [29] 李灵智, 黄洪亮, 陈帅, 等. 基于静水池模型试验的金枪鱼围网沉降性能研究[J]. 中国海洋大学学报, 2015, 45(3): 48-53. doi: 10.16441/j.cnki.hdxb.20130275 [30] 徐国强, 许柳雄, 周成, 等. 基于海上实测的金枪鱼围网下纲沉降及提升性能[J]. 海洋渔业, 2015, 37(2): 171-178. https://www.doc88.com/p-3827777355463.html [31] 袁军亭. 网片的三维力学模型研究及应用[D]. 上海: 上海海洋大学, 2007. https://xueshu.baidu.com/usercenter/paper/show?paperid=455aa289a29ea0e71f45ac9155cdb80b&site=xueshu_se&hitarticle=1 [32] 黄小华, 郭根喜, 胡昱, 等. 圆形网衣在水流作用下的运动变形特性[J]. 中国水产科学, 2010, 17(2): 312-319. https://xueshu.baidu.com/usercenter/paper/show?paperid=69cefe25fc788c59234bc177cb8209cd&site=xueshu_se&hitarticle=1 [33] 赵云鹏. 深水重力式网箱水动力特性数值模拟研究[D]. 大连: 大连理工大学, 2007. 10.7666/d.y1205374 [34] 张俊, 陈作志, 陈国宝, 等. 南海鸢乌贼水声学测量和评估相关技术研究[J]. 南方水产科学, 2014, 10(6): 1-11. doi: 10.3969/j.issn.2095-0780.2014.06.001 -
期刊类型引用(4)
1. 袁太平,廖宇琦,黄小华,胡昱,陶启友,李根,庞国良. 深水网箱网衣清洗装备关键部件设计及试验. 南方水产科学. 2024(01): 25-33 .  本站查看
本站查看
2. 魏盛军,蔡文鸿,丁兰,张哲,张艺,潘柏霖,陈思源,杨章武,魏观渊,林元俊,郑国富. 福建海上养殖设施污损生物处理方式现状调查. 渔业研究. 2023(02): 147-154 . 百度学术
3. 黄小华,庞国良,袁太平,胡昱,王绍敏,郭根喜,陶启友. 我国深远海网箱养殖工程与装备技术研究综述. 渔业科学进展. 2022(06): 121-131 . 百度学术
4. 廖宇琦,袁太平,胡昱,王绍敏,陶启友,黄小华. 转盘流道结构对网箱清洗装备喷嘴流场特性的影响. 渔业现代化. 2021(06): 9-18 . 百度学术
其他类型引用(3)
 下载:
下载:
计量
- 文章访问数: 2979
- HTML全文浏览量: 104
- PDF下载量: 818
- 被引次数: 7
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
