
Effects of increasing dietary vitamin C on serum lysozyme activity and antioxidant ability of tissues in Pampus argenteus
-
摘要:
研究了增加饲料中维生素C(VC)质量分数对银鲳(Pampus argenteus)血清溶菌酶活性及组织抗氧化能力的影响。以鱼粉、虾粉为蛋白源,鱼油为脂肪源配制VC质量分数分别为104.21 mg·kg-1、455.33 mg·kg-1和800.54 mg·kg-1的3组饲料,依次编号为C1(对照组)、C2、C3。每组饲料设3个重复,投喂平均体质量为(6.18±0.15)g的银鲳幼鱼9周。结果显示,随着饲料中VC质量分数的增加,血清溶菌酶活性逐渐升高,C3组较C1、C2组溶菌酶活性显著性升高(P < 0.05)。增加VC质量分数可显著提高组织中的超氧化物歧化酶(SOD)活性及总抗氧化能力(T-AOC)(P < 0.05),但VC对不同组织中SOD活性的影响程度并不相同。随着饲料中VC质量分数的增加,肝脏与肌肉中VC水平显著性升高(P < 0.05)。同时,高水平VC也显著降低了组织中丙二醛(MDA)的质量摩尔浓度。分析表明,增加饲料中VC质量分数(大于455.33 mg·kg-1)可提高银鲳血清溶菌酶活性与组织抗氧化能力,降低组织中的脂质过氧化反应。
Abstract:This study investigated the effects of increasing dietary vitamin C on serum lysozyme activity and antioxidant ability of tissues in silver pomfret (Pampus argenteus). The basal practical diet was formulated with fish meal and shrimp meal as protein sources, and fish oil as lipid source. There were three experimental diets containing ascorbic acid 104.21 mg·kg-1 (C1), 455.33 mg·kg-1 (C2) and 800.54 mg·kg-1 (C3) respectively, and triplicate feeding by every diet group to the juvenile with initial average weight (6.18±0.15) g for 9 weeks. The results show that serum lysozyme activity increases with the increase of dietary vitamin C, and lysozyme activity of C3 diet group is significantly higher than those of C1 and C2 diet groups (P < 0.05). The superoxide dismutase (SOD) activities significantly increase with the increasing vitamin C supplementation (P < 0.05), but the degree of influence is different in various tissues. High dietary vitamin C also significantly promotes the total antioxidant ability (T-AOC) of tissues (P < 0.05).Vitamin C concentrations in liver and muscle significantly improve with increasing vitamin C supplementation, while the malonaldehyde (MDA) contents of tissues significantly reduce with the increase of dietary vitamin C. In conclusion, increasing dietary vitamin C (>455.33 mg·kg-1) can significantly improve the serum lysozyme activity and tissue antioxidant ability, and reduce the tissue lipid peroxidation of silver pomfret.
-
Keywords:
- silver pomfret /
- vitamin C /
- lysozyme /
- antioxidant
-
盐度是影响河口、过河口及海水水生生物生长和发育的重要环境因子,与水生生物的性腺发育关系密切。研究表明,成体金钱鱼 (Scatophagus argus)[1]、印度囊鳃鲶 (Heteropneustes fossilis)[2]、红眼沙鲈(Psammoperca waigiensis)[3-4]、梭鱼 (Liza haematocheila)[5]、凡纳滨对虾 (Litopenaeus vannamei)[6]、中华绒螯蟹(Eriocheir sinensis)[7]等的性腺需要在一定盐度刺激下才可正常发育。
刀鲚 (Coilia nasus) 是长江口重要的洄游性经济鱼类[8],主要分布于中国、韩国沿海以及日本的有明海[9-12],因味道鲜美、肉质细嫩成为重要的经济鱼类[13]。近年来由于水体污染、水利兴修、资源过度开发等因素,进入产卵场的刀鲚种群数量显著降低,以致不能形成常规的渔汛[14-15]。盐度是刀鲚在洄游过程中最直观的影响因素[16],研究显示,进行生殖洄游的鱼类须到达特定盐度才能完成卵巢的正常发育,才会发生产卵行为[17],如鲥 (Tenualosa reevesii)、暗纹东方鲀 (Takifugu obscurus)、大麻哈鱼 (Oncorhynchus keta) 的性腺发育和成熟仅在盐度低于0.5的淡水环境中进行;而日本鳗鲡 (Anguilla japonica)、松江鲈 (Trachidermus fasciatus)的性腺发育与繁殖需要盐度较高的海水环境[17]。近年,Xu等[18]通过代谢组学对刀鲚卵巢发育不同步的问题进行了研究,Yin等[19]通过代谢组学研究了刀鲚在产卵迁移过程中能量的消耗和储存,但未明确指出海、淡水对卵巢发育的影响。为了更好地保护并合理利用刀鲚资源,需要通过人工养殖来维持其资源量,因此,对刀鲚在人工海、淡水环境下卵巢发育机制的研究十分重要。
代谢组学技术作为近年新兴的一种高通量测序技术,在各研究领域尤其是环境对物种的影响中得到广泛应用[20-22],代谢组代表生物细胞、组织、器官或生物体中的完整代谢产物,这些代谢产物可以通过代谢组学的方法进行测定[23]。关于刀鲚卵巢发育的代谢组学研究较少,本研究主要通过液相色谱-质谱联用 (Liquid chromatograph-mass spectrometer, LC-MS) 的代谢组学技术分析筛选养殖在淡水和海水2种环境下刀鲚卵巢组织的差异代谢产物,并对差异代谢产物进行代谢通路富集分析,为进一步研究刀鲚在生殖洄游过程中卵巢组织的发育及其机制以及人工养殖提供参考依据。
1. 材料与方法
1.1 实验材料
实验于2018年12月—2019年2月在上海市水产研究所奉贤基地进行,实验材料选用该基地提供的野生型F3代,Ⅱ龄刀鲚。实验前将刀鲚放入淡水池中 (长5 m×宽3 m×高1 m) 中暂养2周,其后随机分到淡水组 (实验组) 和海水组 (对照组,盐度为25) 水泥池中养殖,每组100尾。海水从河口抽取,盐度约为9,用日晒盐将盐度提高到25。每天08:00、15:00投喂从池塘捞取的活饵 (桡足类、糠虾等),每14 d换水1次。
1.2 实验方法
1.2.1 采样方法
养殖实验结束后,停食1 d。将刀鲚置于30 mg·L−1丁香酚溶液中麻醉5 min后,用干毛巾将鱼体表面擦干,取其性腺,一半装于2 mL的冻存管,迅速置于液氮罐中冷冻,之后于−80 ℃保存以备刀鲚卵巢代谢组学分析;另一半置于Bouin's液中固定24 h,通过石蜡切片和HE染色确定性别后,将海、淡水刀鲚的卵巢分别每3个样混合,每组6个平行,平均体长为 (21.00±2.67) cm,平均体质量为 (25.80±9.05) g。
1.2.2 样本处理
称取刀鲚卵巢样本20 mg ,加入400 μL提取液 [V(甲醇)∶V(水)=4∶1],利用高通量组织破碎仪破碎 (−20 ℃, 50 Hz, 6 min);涡旋 (30 s) 混匀后,低温超声萃取30 min (5 ℃, 40 kHz);将其置于−20 ℃ 30 min;离心(13 000×g, 4 ℃, 15 min),去掉上清液,并加入100 μL复溶液 [V(乙腈)∶V(水)=1∶1] 复溶,同时将所有样本的提取液等体积混匀制备质控样本 (Quality control, QC),每个QC的体积与样本体积相同。将所测样本和QC转移至LC-MS 进样瓶进行上机分析。本次LC-MS 分析的仪器平台为AB SCIEX公司的超高效液相色谱串联飞行时间质谱UPLC-TripleTOF系统。
1.2.3 色谱条件
实验采用美国Milford公司BEH C18色谱柱 (100 mm × 2.1 mm,1.7 μm)。配置流动相A 为水 (含0.1%甲酸),流动相B 为V(乙腈)∶V(异丙醇)=1∶1 (含0.1%甲酸),进行梯度洗脱,流速为0.40 mL·min−1,进样量为20 μL,柱温为40 ℃。
1.2.4 质谱条件
样品质谱信号采集分别采用正负离子扫描模式、离子喷雾电压。质谱检测范围为50~1 000 m.z−1。ESI离子源参数设置雾化气压 (GS1) 50 psi;辅助气压 (GS2) 50 psi;气帘气压:30 psi;温度:500 ℃;喷雾电压:5 000 V (POS模式)或−4 000 V (NEG模式)。
1.3 数据处理与分析
首先采用代谢组学Progenesis QI处理软件 (Waters Corporation, Milford, USA)过滤质谱数据的低质量峰,填充缺失值,归一化,评估QC样本RSD,数据log转换预处理。将原始数据用Progenesis QI进行搜库鉴定,将质谱信息与代谢数据库HMDB和METLIN进行匹配。
采用R软件包Ropls进行多元统计分析:首先将所有实验样品与QC样品比较后进行主成分分析 (Principal Component Analysis, PCA)和正交偏最小二乘判别分析(Orthogonal Partial Least Squares Discrimination Analysis, OPLS-DA),再经200次置换检验以判断模型是否过拟合。
采用t检验并结合OPLS-DA分析中的VIP值,筛选出组间的差异代谢物,筛选条件为差异倍数(Fold-change FC>1)、P(<0.05)和变量投影重要度(Variable importance in the projection, VIP>1),三者取交集。代谢通路的注释分析基于KEGG数据库。
将得到的差异代谢产物在KEGG Compound和HMDB数据库进行化合物分类情况统计。
2. 结果
2.1 多元统计分析结果
首先对淡水组 (N=6) 和海水组 (N=6) 的样品进行PCA和OPLS-DA分析以评估两组间的统计学差异,发现所有样品均处于样本置信区间。十字架表示的质控样本比较集中,说明仪器的稳定性好,检测的数据可靠,可用于PCA和OPLS-DA分析。PCA得分结果显示,淡水组和海水组样品点在空间上具有明显的分离,表明两组间的代谢模式存在差异 (图1-a);OPLS-DA分析可看出两组数据的样品点分别聚为一簇,在空间上呈现分离状态,表明淡水组和海水组间产生了显著差异 (图1-b),模型的评价指标R2=0.971,Q2=0.562,说明所构建的模型良好。为进一步验证模型是否过拟合,采用200次置换检验 (图2),可知Q2于y轴的截距为负,且置换检验得到的R2=0.873 5和Q2=−0.146 4均小于原始值,由此可见模型没有过拟合,可以用于后续差异代谢物筛选的分析。
![]() 图 1 海水组和淡水组的主成分分析 (a) 和正交偏最小二乘判别分析 (b) 得分图Figure 1. PCA (a) and OPLS-DA (b) scores of seawater and freshwater
图 1 海水组和淡水组的主成分分析 (a) 和正交偏最小二乘判别分析 (b) 得分图Figure 1. PCA (a) and OPLS-DA (b) scores of seawater and freshwater2.2 总代谢产物Venn图
对在阳离子和阴离子2种模式下获得的海、淡水养殖刀鲚的卵巢代谢产物数据分别构建Venn图(图3)。在阳离子分离模式下,2组卵巢组织中共有代谢产物310个,其中淡水组中特有代谢产物9个,海水组12个;在阴离子分离模式下,2组共有代谢产物392个,其中淡水组中特有代谢产物也为9个,海水组则降到10个。
2.3 差异代谢产物注释
海水和淡水养殖刀鲚卵巢组织样本中的差异代谢产物的火山图见图4,差异代谢产物具体信息详见表1。与海水组相比,淡水组刀鲚卵巢组织有47个代谢产物表现出显著差异性,其中11个代谢产物有上调表达,36个代谢产物有下调表达,表达差异程度最显著的为碳环血氧烷A2 (Carbocyclic thromboxane A2)、半乳糖神经酰胺 (Galactosyl ceramide),差异倍数分别为10.40、2.78倍。根据表1,淡水组的皮质醇含量比海水组高1.61倍,且呈显著性差异 (P<0.05)。
表 1 差异代谢物信息表Table 1. Differential metabolites information sheet差异代谢物
Differential metabolite分子式
Molecular formula变量投影重要度
VIPFC
Fold changeP 变化趋势
Variation trend磷脂酰肌醇 PI [20:4(5Z,8Z,11Z,14Z)/0:0] C29H49O12P 1.58 0.93 0.017 下降 谷氨酰胺色氨酸 Glutaminyltryptophan C16H20N4O4 1.05 0.96 0.018 下降 L-异亮氨酸 L-Isoleucine C6H13NO2 1.33 0.96 0.005 下降 木麻黄6-α-D-葡萄糖苷 Casuarine 6-alpha-D-glucoside C14H25NO10 1.13 0.94 0.043 下降 甘油一脂 MG(10:0/0:0/0:0) C13H26O4 3.59 0.71 0.025 下降 皮质醇 Cortisol C21H30O5 3.28 1.61 0.008 上升 碳环血氧烷A2 Carbocyclic thromboxane A2 C22H36O3 2.88 10.40 0.007 上升 (±)9-十八碳二烯酸 (±)9-HPODE C18H32O4 2.90 0.58 0.020 下降 N-棕榈酰蛋氨酸 N-palmitoyl methionine C21H41NO3S 1.86 0.75 0.022 下降 13-羟基十八酸 13-hydroxyoctadecanoic acid C18H36O3 1.95 0.86 0.005 下降 咖啡酰环戊醇 Caffeoylcycloartenol C39H56O4 2.07 0.81 0.036 下降 11,13-二十碳二烯酸 15-OxoEDE C20H34O3 2.51 0.70 0.032 下降 2,3-二氢苯并呋喃 2,3-dihydrobenzofuran C8H8O 1.19 1.08 0.033 上升 4-甲酰基吲哚 4-formyl indole C9H7NO 1.34 0.83 0.040 下降 红花素C Safflomin C C30H30O14 2.04 0.85 0.003 下降 6-[(2-羧基乙酰基)氧]-3,4,5-三羟基氧烷-2-羧酸6-[(2-carboxyacetyl)oxy]-3,4,5-trihydroxyoxane-2-carboxylic acid C9H12O10 2.07 1.12 0.001 上升 异戊二烯 Isoputreanine C7H16N2O2 2.33 0.84 0.001 下降 脯氨酸 L-Proline C5H9NO2 1.80 0.91 0.003 下降 羟脯氨酸 Hydroxyprolyl-hydroxyproline C10H16N2O5 3.13 0.72 0.007 下降 5-O-阿魏酰黑精 5-O-Feruloylnigrumin C21H25NO10 1.0 0.95 0.037 下降 壬二酸 Azelaic acid C9H16O4 1.12 0.96 0.012 下降 尿苷 Uridine C9H12N2O6 1.35 0.95 0.001 下降 视黄酯 Retinyl ester C20H30O2 1.24 0.96 0.004 下降 顺-9,10-环氧硬脂酸 cis-9,10-Epoxystearic acid C18H34O3 1.66 0.92 0.025 下降 异柠檬酸盐 Isocitrate C6H8O7 1.55 1.12 0.047 上升 戊二酸 Glutaric acid C5H8O4 2.53 1.38 0.015 上升 6-脱氧噬菌体胺 (6-脱氧花青) 6-Deoxyfagomine C6H13NO2 1.36 0.94 0.012 下降 乙酰-L-酪氨酸 Acetyl-L-tyrosine C11H13NO4 1.38 0.87 0.010 下降 γ-谷氨酰鸟氨酸 Gamma glutamyl ornithine C10H19N3O5 1.87 1.58 0.023 上升 对茴香酸异戊酯 (异戊基异茴香酸酯) Isoamyl p-anisate C13H18O3 1.76 0.67 0.041 下降 3-氧十二酸 3-Oxododecanoic acid C12H22O3 1.01 0.92 0.043 下降 C-2神经酰胺 C-2 Ceramide C20H39NO3 2.21 0.75 0.028 下降 磷脂酰乙醇胺 PE(15:0/16:1(9Z)) C36H70NO8P 1.42 1.10 0.030 上升 半乳糖神经酰胺 Galactosyl ceramide (d18:1/14:0) C38H73NO8 1.97 2.78 0.045 上升 N-棕榈酰甘氨酸 N-Palmitoyl glycine C18H35NO3 1.97 0.86 0.000 下降 二十碳五烯酸 Eicosapentaenoic acid C20H30O2 1.01 0.98 0.004 下降 溶血磷脂酰乙醇胺 LysoPE(0:0/20:2(11Z,14Z)) C25H48NO7P 1.33 1.12 0.037 上升 9-羟基癸酸 9-Hydroxydecanoic acid C10H20O3 1.21 0.92 0.028 下降 甘油一脂 MG(a-13:0/0:0/0:0)[rac] C16H32O4 1.92 0.79 0.040 下降 反式-2-十二碳烯二酸 Traumatic acid C12H20O4 1.37 0.86 0.025 下降 9-氧壬酸 9-Oxo-nonanoic acid C9H16O3 1.15 0.93 0.019 下降 8-羟基-5,6-辛二烯酸 8-Hydroxy-5,6-octadienoic acid C8H12O3 1.04 0.94 0.016 下降 (S)-3-磺酸盐 (S)-3-Sulfonatolactate C3H6O6S 2.57 3.05 0.046 上升 γ-谷氨酰缬氨酸 gamma-Glutamylvaline C10H18N2O5 1.80 0.87 0.008 下降 穗花牡荆苷 Agnuside C22H26O11 1.41 0.91 0.005 下降 假尿苷 Pseudouridine C9H12N2O6 1.19 0.93 0.003 下降 雌三醇7-(6-反式-对-香豆酰基葡萄糖苷)
Eriodictyol 7-(6-trans-p-coumaroylglucoside)C30H28O13 1.44 0.92 0.020 下降 注:FC表示某差异代谢物在淡水组相对于海水组的表达倍数变化,FC>1表示该代谢物上调,FC<1表示该代谢物下调;P<0.05表示差异显著,P<0.01表示差异极显著。 Note: FC value indicates the change of expression multiple of a differential metabolite in FOV group compared with SOV group. FC>1 indicates that the metabolite is upregulated; FC<1 indicates that the metabolite is down-regulated. P<0.05 indicates significant difference, and P<0.01 indicates extremely significant difference. 将上述47个差异表达代谢产物比对到KEGG 数据库进行富集分析,共有8个代谢产物显著富集到12条KEGG通路上 (图5和表2)。显著差异代谢产物的富集通路主要涉及氨酰-tRNA生物合成 (Aminoacyl-tRNA biosynthesis)、嘧啶代谢 (Pyrimidine metabolism)、鞘磷脂代谢 (Sphingolipid metabolism)和三羧酸循环 (TCA cycle)。上调表达的2个代谢产物L-脯氨酸 (L-Proline)和L-异亮氨酸 (L-Isoleucine)参与到氨酰tRNA生物合成通路上。下调表达的2个代谢产物尿苷和假尿苷参与到嘧啶代谢通路中。在实验组卵巢组织中上调表达的半乳糖神经酰胺和异柠檬酸 (Isocitrate) 代谢物分别参与到鞘磷脂代谢和三羧酸循环通路中。
表 2 差异代谢产物的KEGG通路富集表Table 2. KEGG pathway enrichment table of differential metabolites富集的差异代谢产物名称
Enriched differential metabolite name富集个数
Numble富集通路
ID Pathway通路描述
Pathway desciptionP 半乳糖神经酰胺 Galactosyl ceramide (d18:1/14:0) 1 map00600 鞘脂代谢 0.059 脯氨酸 L-Proline 1 map00330 精氨酸和脯氨酸代谢 0.176 异柠檬酸盐 Isocitrate 1 map00020 柠檬酸循环 0.057 戊二酸 Glutaric acid 1 map00310 赖氨酸降解 0.130 皮质醇 Cortisol 1 map00140 类固醇激素生物合成 0.207 尿苷 Uridine;假尿苷 Pseudo uridine 2 map00240 嘧啶代谢 0.011 异柠檬酸盐 Isocitrate 1 map00630 乙醛酸和二羧酸代谢 0.137 L-异亮氨酸 L-Isoleucine;脯氨酸 L-Proline 2 map00970 氨酰tRNA生物合成 0.008 L-异亮氨酸 L-Isoleucine 1 map00290 缬氨酸、亮氨酸和异亮氨酸生物合成 0.064 二十碳五烯酸 Eicosapentaenoic acid 1 map01040 不饱和脂肪酸生物合成 0.097 L-异亮氨酸 L-Isoleucine 1 map00280 缬氨酸、亮氨酸和异亮氨酸降解 0.107 (S)-3-磺酸盐 (S)-3-Sulfonatolactate 1 map00270 半膀氨酸和蛋氨酸代谢 0.141 将得到的差异代谢产物比对到HMDB数据库,统计发现占比最多的是脂质和类脂分子,高达41.18%;其次为有机酸及其衍生物,占比35.29%。
将代谢差异物比对到KEGG Compound数据库,发现聚类最多的是氨基酸和羧酸;比对到KEGG Pathway数据库,发现富集程度最高的分别是脂质代谢通路 (Lipid metabolism)、氨基酸代谢通路和消化系统。
3. 讨论
通过非靶向代谢组学得到刀鲚卵巢在海、淡水环境下47种差异代谢物,发现代谢物主要包括脂质和类脂、有机酸及衍生物、氨基酸及羧酸,这与Xu等[18]对刀鲚卵巢Ⅲ—Ⅴ期发育过程的研究结果相似,差异代谢物主要涉及糖酵解和糖异生、组氨酸代谢、赖氨酸代谢以及脂肪酸的代谢相关通路。由此可见,海、淡水环境对人工养殖刀鲚的卵巢发育具有重要作用。
碳环血氧烷A2是由活化的血小板在止血过程中产生的,具有凝血之前的特性,也是一种已知的血管收缩剂[24-26],本实验淡水组碳环血氧烷A2高表达可能是刀鲚机体组织为应对进入到淡水中盐度降低作出的适应性调节。半乳糖神经酰胺是一种强免疫兴奋剂,在许多体内实验模型中显示出强大的抗肿瘤活性[27],虹鳟 (Oncorhynchus mykiss) 在洄游产卵期会遇到海水中不同的病毒、细菌等进而增强体内免疫系统[28],本研究与此类似,刀鲚为应对环境改变所带来的免疫胁迫压力,也会通过增强特殊的免疫通路来完成洄游繁殖进程。
皮质醇是一类类固醇激素,属于糖皮质激素类,其通过糖异生作用提高血糖水平,也具有抑制免疫系统的功能,有助于脂肪、蛋白质和碳水化合物的代谢[29]。研究表明,在产卵季节,大麻哈鱼的血浆皮质醇水平会有升高现象[30]。皮质醇在薄唇鲻鱼 (Liza ramada) 卵巢卵黄形成早期呈下降趋势,在卵黄中期呈上升趋势,在卵黄后期下降,在产卵前期上升到较高水平,皮质醇的这种周期性变化支持了卵巢的发育和产卵[31]。类固醇代谢在卵巢发育中发挥着重要作用,其主要由内分泌腺如精巢、卵巢和肾上腺等单位合成之后释放到血液循环中,而所有的类固醇激素均由胆固醇合成,Xu等[18]研究指出,胆固醇合成途径在刀鲚卵巢发育过程中 (Ⅲ—Ⅴ期) 逐渐增强。本实验中皮质醇在淡水环境下比海水环境高1.61倍,每年2月初次性成熟的刀鲚开始从沿海区域进入长江等近海淡水区进行生殖洄游,在此过程中卵巢逐渐发育成熟,说明皮质醇在刀鲚洄游过程中参与了卵巢的发育过程,可能进一步促进了卵巢发育。此外,鱼体内的皮质醇具调节渗透压作用,能够刺激鱼类鳃上皮氯细胞密度增加,增强鳃内Na+/K+-ATPase活性,使过多的盐分排除体外[32-34],推测皮质醇在刀鲚的洄游过程中参与了渗透压的调节。
氨酰-tRNA合成酶在翻译中的作用是通过准确地将同源tRNA与相应的氨基酸配对来定义遗传密码,增强生物体在蛋白质翻译水平的准确性[35-36],本实验中下调的差异代谢物L-脯氨酸和L-异亮氨酸被富集到氨酰-tRNA生物合成代谢通路上,表明刀鲚卵巢组织为适应低盐度的环境可能相应地降低了蛋白质的翻译。同样下调表达的尿苷和假尿苷2个差异代谢产物富集到嘧啶代谢通路中,该通路的产物β-氨基丁酸 (β-aminobutyric acid) 可以参与到TCA循环中[37] ,表明淡水养殖环境下刀鲚卵巢组织内嘧啶类代谢受到部分抑制,甚至可影响能量代谢。刀鲚卵巢组织中鞘磷脂代谢在淡水环境中得到提升,被富集到该通路的半乳糖神经酰胺可稳固细胞膜脂质双分子层的机械稳定和化学抗性[38-39],由此可见,在刀鲚洄游过程中神经酰胺类代谢产物在信号转导和细胞识别中发挥重要作用。
4. 结论
本研究使用LC-MS非靶向代谢组学分析方法,研究了养殖在海、淡水中刀鲚的卵巢组织代谢产物的变化,共筛选得到47种差异代谢产物,推测碳环血氧烷A2和半乳糖神经酰胺的高表达,分别是为了更好地适应环境变化作出的适应性调节和增强免疫;淡水环境下皮质醇大量积累,推测皮质醇在刀鲚的洄游过程中参与了渗透压调节,同时可能与卵巢的发育相关;对差异代谢产物进行KEGG通路富集分析,筛选出氨酰-tRNA生物合成、嘧啶代谢和鞘磷脂代谢3条最显著的代谢途径,推测盐度的变化对刀鲚卵巢组织中的氨基酸、嘧啶、鞘磷脂的合成代谢有较大影响。
-
![]()
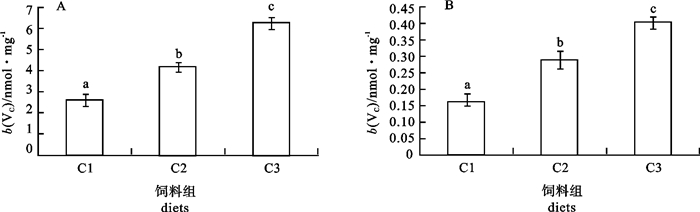
图 1 增加饲料中VC质量分数对银鲳血清溶菌酶活性的影响
各处理组间不同标示字母表示差异显著(P < 0.05),后图同此
Figure 1. Effects of increasing dietary VC on the serum lysozyme activity in silver pomfret
Columns with different superscripts are significantly different (P < 0.05). The same case in the following figures.
![]()
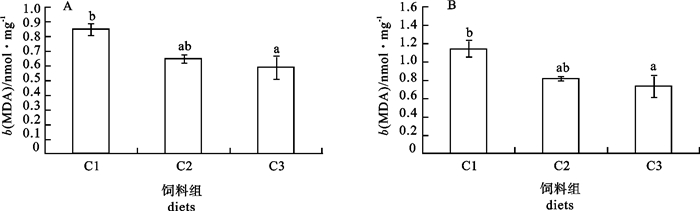
图 2 增加饲料中VC质量分数对银鲳肝脏(A)和肌肉(B)中VC质量摩尔浓度的影响
Figure 2. Effects of increasing dietary VC on the liver (A) and muscle (B) VC contents in silver pomfret
![]()
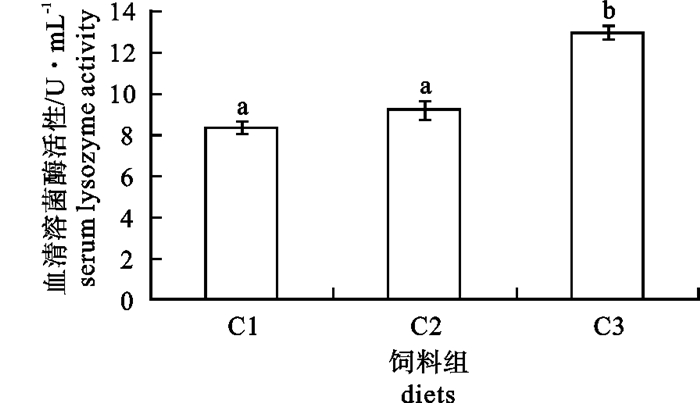
图 3 增加饲料中VC质量分数对银鲳肝脏(A)和肌肉(B)丙二醛质量摩尔浓度的影响
Figure 3. Effects of increasing dietary VC on the liver (A) and muscle (B) MDA contents in silver pomfret
表 1 对照组(C1组)基础饲料的组成
Table 1 Composition of the control diet
饲料组成
diet composition质量分数/g·kg-1
content饲料组成
diet composition质量分数
content鱼粉fish meala 690 营养组成(干质量) proximate composi-tion (dry weight) 虾粉shrimp mealb 60 粗蛋白/% crude protein 46.53 鱼油fish oil 60 粗脂肪/% crude fat 12.37 羧甲基纤维素carboxymethyl cellulosec 15 灰分/% ash 11.26 α-淀粉α-starch 135 维生素C/mg·kg-1 ascorbic acid 104.21 复合维生素vitamin premixd 20 复合矿物质mineral premixe 20 注:a. 鱼粉含蛋白62%,脂肪9%,灰分13%;b. 虾粉含蛋白44%,脂肪8%,灰分7%;c. 饲料粘合剂;d. 复合维生素(mg·kg-1)为肌醇400,烟酸150,泛酸钙44,核黄素20,维生素B6 12,维生素K 10,维生素B1 10,维生素A 7.3,叶酸5,生物素1,维生素D3 0.06,维生素B12 0.02,维生素C 100,维生素E 35;e. 复合矿物质(mg或mg·kg-1)为磷酸二氢钾(KH2PO4)22 g,硫酸铁(FeSO4·7H2O)1.0 g,硫酸锌(ZnSO4·7H2O)0.13 g,硫酸锰(MnSO4·4H2O)52.8 mg,硫酸铜(CuSO4·5H2O)12 mg,硫酸钴(CoSO4 ·7H2O)2 mg,碘化钾(KI)2 mg
Note:a. fish meal contain protein 62%,fat 9%,ash 13%;b. shrimp meal contain protein 44%,fat 8%,ash 7%;c. diet adhesive;d. vitamin premix (mg·kg-1) contain inositol 400,nicotinic acid 150,calcium pantothenate 44,lactochrome 20,vitamin B6 12,vitamin K 10,vitamin B1 10,vitamin A 7.3,folic acid 5,biotin 1,vitamin D3 0.06,vitamin B12 0.02,vitamin C 100,vitamin E 35;e. mineral premix (mg or mg·kg-1) contain KH2PO4 22 g,FeSO4·7H2O 1.0 g,ZnSO4·7H2O 0.13 g,MnSO4·4H2O 52.8 mg,CuSO4·5H2O 12 mg,CoSO4·7H2O 2 mg,KI 2 mg. 下载: 导出CSV
下载: 导出CSV
表 2 增加饲料中VC质量分数对组织中超氧化物歧化酶活性和总抗氧化能力的影响(X ±SE)
Table 2 Effects of increasing dietary VC on the tissue SOD activities and T-AOC in silver pomfret
饲料
diets超氧化物酶活性SOD activity 总抗氧化能力T-AOC 血清/U·mL-1
serum肌肉/U·mg-1prot
muscle肝脏/U·mg-1prot
liver血清/U·mL-1
serum肌肉/U·mg-1prot
muscle肝脏/U·mg-1prot
liverC1 78.25±6.41a 35.04±0.21a 371.24±18.47a 5.61±0.20a 0.32±0.03a 5.86±0.19a C2 109.83±4.70b 38.37±2.12ab 434.76±12.93a 8.08±0.65b 0.89±0.03b 8.58±0.18b C3 127.69±4.47b 41.30±0.68b 547.04±22.48b 8.55±0.40b 0.95±0.03b 9.15±0.23b 注:同一列不同标示字母表示差异显著(P < 0.05)
Note:Different superscript letters within each column represent significant difference (P < 0.05).
下载: 导出CSV
-
[1] FRACALOSSI D M, ALLEN M E, YUYAMA L K, et al. Ascorbic acid biosynthesis in Amazonian fishes[J]. Aquaculture, 2001, 192(2/3/4): 321-332. doi: 10.1016/S0044-8486(00)00455-5
[2] AI Q H, MAI K S, ZHANG C X, et al. Effects of dietary vitamin C on growth and immune response of Japanese seabass, Lateolabrax japonicus[J]. Aquaculture, 2004, 242(1/2/3/4): 489-500. doi: 10.1016/j.aquaculture.2004.08.016
[3] AI Q H, MAI K S, TAN B P, et al. Effects of dietary vitamin C on survival, growth, and immunity of large yellow croaker, Pseudosciaena crocea[J]. Aquaculture, 2006, 261(1): 327-336. doi: 10.1016/j.aquaculture.2006.07.027
[4] LIM C, KLESIUS P H, LI M H, et al. Interaction between dietary levels of iron and vitamin C on growth, hematology, immune response and resistance of channel catfish (Ictalurus punctatus) to Edwardsiella ictaluri challenge[J]. Aquaculture, 2000, 185(3/4): 313-327. doi: 10.1016/S0044-8486(99)00352-X
[5] ORTUNO J, ESTEBAN M A, CUESTA A, et al. Effect of oral administration of high vitamin C and E dosages on the gilthead seabream (Sparus aurata L. ) innate immune system[J]. Veter Immunol Immunopathol, 2001, 79(3/4): 167-180. doi: 10.1016/s0165-2427(01)00264-1
[6] SATO M, KONDO T, YASHINAKA R, et al. Effect of dietary ascorbic acid levels on collagen formation in rainbow trout[J]. Bull Jap Soc Sci Fish, 1982, 48(4): 553-556. doi: 10.2331/suisan.48.553
[7] CHEN R G, LOCHMANN R, GOODWIN A, et al. Effects of dietary vitamins C and E on alternative complement activity, hematology, tissue composition, vitamin concentrations and response to heat stress in juvenile golden shiner (Notemigonus crysoleucas)[J]. Aquaculture, 2004, 242(1/2/3/4): 553-569. doi: 10.1016/j.aquaculture.2004.09.012
[8] SOBHANA K S, MOHAN C V, SHANKAR K M. Effect of dietary vitamin C on the disease susceptibility and inflammatory response of mrigal, Cirrhinus mrigala (Hamilton) to experimental infection of Aeromonas hydrophila[J]. Aquaculture, 2002, 207(3/4): 225-238. doi: 10.1016/S0044-8486(01)00793-1
[9] 冯伟, 李健, 李吉涛, 等. Vc对中国对虾非特异免疫因子及TLR/NF-κB表达量的影响[J]. 水产学报, 2011, 35(2): 200-207. doi: 10.3724/SP.J.1231.2011.17118 [10] 徐维娜, 刘文斌, 邵仙萍, 等. 维生素C对异育银鲫原代肝脏细胞活性及抗敌百虫氧化胁迫的影响[J]. 水产学报, 2011, 35(12): 1849-1856. doi: 10.3724/SP.J.1231.2011.17577 [11] 赵红霞, 曹俊明, 谭永刚, 等. 军曹鱼幼鱼维生素C需要量的研究[J]. 动物营养学报, 2008, 20(4): 435-441. doi: 10.3969/j.issn.1006-267X.2008.04.010 [12] 明建华, 谢骏, 徐跑, 等. 大黄素、维生素C及其配伍对团头鲂生长、生理生化指标、抗病原感染以及两种HSP70s mRNA表达的影响[J]. 水产学报, 2010, 34(9): 1447-1459. doi: 10.3724/SP.J.1231.2010.06891 [13] 刘海燕, 雷武, 朱晓鸣, 等. 饲料中不同维生素C含量对长吻 
[14] 胡斌, 李小勤, 冷向军, 等. 饲料VC对草鱼生长、肌肉品质及非特异性免疫的影响[J]. 中国水产科学, 2008, 15(5): 794-800. doi: 10.3321/j.issn:1005-8737.2008.05.011 [15] ORTUNO J, ESTEBAN M A, MESEGUER J. The effect of dietary intake of vitamins C and E on the stress response of gilthead seabream (Sparus aurata L. )[J]. Fish & Shellfish Immunol, 2003, 14(2): 145-156. doi: 10.3969/j.issn.1000-9957.2003.02.014
[16] LIN M F, SHIAU S Y. Dietary l-ascorbic acid affects growth, nonspecific immune responses and disease resistance in juvenile grouper, Epinephelus malabaricus[J]. Aquaculture, 2005, 244(1/2/3/4): 215-221. doi: 10.1016/j.aquaculture.2004.10.026
[17] 彭士明, 施兆鸿, 孙鹏, 等. 养殖密度对银鲳幼鱼生长及组织生化指标的影响[J]. 生态学杂志, 2010, 29(7): 1371-1376. doi: 10.13292/j.1000-4890.2010.0228 [18] 彭士明, 尹飞, 孙鹏, 等. 不同饲料对银鲳幼鱼增重率、肝脏脂酶及抗氧化酶活性的影响[J]. 水产学报, 2010, 34(6): 769-774. doi: 10.3724/SP.J.1231.2010.06823 [19] 尹飞, 孙鹏, 彭士明, 等. 低盐度胁迫对银鲳幼鱼肝脏抗氧化酶、鳃和肾脏ATP酶活力的影响[J]. 应用生态学报, 2011, 22(4): 1059-1066. doi: 10.13287/j.1001-9332.2011.0107 [20] 彭士明, 施兆鸿, 李杰, 等. 运输胁迫对银鲳血清皮质醇、血糖、组织中糖元及乳酸含量的影响[J]. 水产学报, 2011, 35(6): 831-837. doi: 10.3724/SP.J.1231.2011.17242 [21] HOSSAIN M A, ALMATAR S M, JAMES C. Optimum dietary protein level for juvenile silver pomfret, Pampus argenteus (Euphrasen)[J]. J World Aquac Soc, 2010, 41(5): 710-720. doi: 10.1111/j.1749-7345.2010.00413.x
[22] 钟国防, 钱曦, 华雪铭, 等. 玉米蛋白粉替代鱼粉对暗纹东方鲀溶菌酶活性及c型溶菌酶mRNA表达的影响[J]. 水产学报, 2010, 34(7): 1121-1128. doi: 10.3724/SP.J.1231.2010.06711 [23] 苏慧, 区又君, 李加儿, 等. 饥饿对卵形鲳鲹幼鱼不同组织抗氧化能力、Na+/K+-ATP酶活力和鱼体生化组成的影响[J]. 南方水产科学, 2012, 8(6): 28-36. doi: 10.3969/j.issn.2095-0780.2012.06.005 [24] 葛颖华, 钟晓明. 维生素C和维生素E抗氧化机制及其应用的研究进展[J]. 吉林医学, 2007, 28(5): 707-708. doi: 10.3969/j.issn.1004-0412.2007.05.066 [25] WANG X J, KIM K W, BAI S C, et al. Effects of the different levels of dietary vitamin C on growth and tissue ascorbic acid changes in parrot fish (Oplegnathus fasciatus)[J]. Aquaculture, 2003, 215(1): 203-211. doi: 10.1016/S0044-8486(02)00042-X
[26] PENG S M, CHEN L Q, QIN J G, et al. Effects of dietary vitamin E supplementation on growth performance, lipid peroxidation and tissue fatty acid composition of black seabream (Acanthopagrus schlegeli) fed oxidized fish oil[J]. Aquac Nutr, 2009, 15(3): 329-337. doi: 10.1111/j.1365-2095.2009.00657.x
[27] 秦洁芳, 陈海刚, 蔡文贵, 等. 邻苯二甲酸二丁酯对汉氏棱鳀生化指标的影响[J]. 南方水产科学, 2011, 7(2): 29-34. doi: 10.3969/j.issn.2095-0780.2011.02.005 [28] 黄志斐, 马胜伟, 张喆, 等. BDE3胁迫对翡翠贻贝(Perna viridis)SOD、MDA和GSH的影响[J]. 南方水产科学, 2012, 8(5): 25-30. doi: 10.3969/j.issn.2095-0780.2012.05.004 -
期刊类型引用(6)
1. 王子涵,曾聪,姜子禺,曹玲. 东海及其邻近海域受胁鱼类保护空缺分析. 热带海洋学报. 2023(01): 66-86 .  百度学术
百度学术
2. 王新月,陈生熬,王程欣,訾方泽,常德胜,许豪,李大鹏. 叶尔羌高原鳅耳石形态探究及群体判别分析. 渔业科学进展. 2023(04): 201-211 . 百度学术
3. 谢玉,玄文丹,周永东,朱凯,朱文斌,王忠明,徐开达,王嘉浩. 东海海域带鱼矢耳石形态特征与生长研究. 浙江海洋大学学报(自然科学版). 2023(04): 294-302 . 百度学术
4. 潘梦珍,王海山,叶乐,陈治,杨超杰. 崖州湾附近海域金线鱼形态学指标的相关性. 海南热带海洋学院学报. 2020(02): 11-17 . 百度学术
5. 蔡研聪,黄梓荣,李佳俊,许友伟,孙铭帅,陈作志,刘维达. 南海北部近海新记录种——苏门答腊金线鱼资源分布特征. 南方水产科学. 2020(06): 1-11 . 本站查看
6. 杨超杰,唐婷婷,吴东霞,杜良玉,方馨正. 崖州湾附近海域金线鱼繁殖生物学研究. 海南热带海洋学院学报. 2019(05): 6-11 . 百度学术
其他类型引用(5)





计量
- 文章访问数: 1163
- HTML全文浏览量: 110
- PDF下载量: 2061
- 被引次数: 11
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
