
Characteristic analysis of fishing condition of saury in Northwest Pacific under different climate regimes
-
摘要: 根据1985—2013年日本秋刀鱼(Cololabis saira)生产数据,结合海表温度(sea surface temperature,SST)、海表温度距平值(sea surface temperature anomaly,SSTA)及亲潮、黑潮环境数据,探究太平洋年代际振荡(Pacific decadal oscillation,PDO)冷、暖2种气候模态下秋刀鱼产卵场、索饵场海况特征及资源丰度差异。结果表明,PDO暖期内单位捕捞量努力量(catch per unit effect,CPUE)略高于冷期,另CPUE与PDO指数存在滞后关系,且前者滞后于后者3年时相关系数最大(– 0.318);PDO冷、暖模态转换对索饵场SST影响较产卵场大(P<0.05),产卵场及索饵场SSTA与PDO指数间均呈负相关性,滞后时间分别为 –1和0年时相关系数最大;PDO暖期内黑潮经向流量年间变化率低于冷期,冷、暖2种模态下亲潮春季占有面积存在显著差异(P<0.01),且冷、暖模态转换对亲潮影响程度大于黑潮。研究表明,2种气候模态通过影响秋刀鱼产卵场及索饵场的海表温、海况环境,进而对西北太平洋秋刀鱼资源量产生年间波动影响。Abstract: We studied the sea current characteristics and the difference of resource abundance in spawning and feeding ground under the effects of two climate regimes of Pacific Decadal Oscillation (PDO) based on the catch data of Pacific saury (Cololabis saira) in Japan from 1985 to 2013, and the environmental data including sea surface temperature (SST), sea surface temperature anomaly (SSTA), Oyashio and Kuroshio data. The results show that the catch per unit effect (CPUE) in warm period was slightly higher than that in cold period, and the maximum correlation coefficient was – 0.318 between CPUE and PDO index when the former lagged the latter three years. The effect of change of PDO index on SST in feeding ground was higher than that in spawning ground (P<0.05) and SSTA was negatively correlated with PDO index in spawning and feeding grounds, and the maximum correlation coefficient was obtained when the lag time was –1 and 0 year. The interannual variability of longitude flow of Kuroshio was lower in warm regime of PDO, while the average occupied area in spring of Oyashio had significant difference under two climate regimes (P<0.01), and the impact degree caused by the change of cold and warm regimes on Oyashio was greater than that on Kuroshio. The results show that two different climate regimes have annual fluctuation influence on saury resource in the Northwest Pacific by affecting SST and sea state condition of spawning and feeding grounds.
-
金枪鱼延绳钓是捕捞栖息于深水层的大型金枪鱼的主要渔具渔法,也是中国远洋金枪鱼渔业最主要的作业方式之一[1]。南海中南部深水区面积广阔,是大洋性金枪鱼类的重要洄游通道之一,蕴藏着丰富的大型金枪鱼资源,目前主要由越南和中国台湾钓船捕捞[2]。2012年越南钓船在南海的大型金枪鱼产量达16 232 t,其中黄鳍金枪鱼(Thunnus albacaes) 12 456 t,大眼金枪鱼(T.obesus) 3 761 t,长鳍金枪鱼(T.alalunga)15 t[3]。
中国大陆1954年~2006年间曾多次组织延绳钓渔船在南海生产,因经济效益不佳未能坚持下来。南海金枪鱼资源密度偏低、渔港离中心渔场太远、专业钓船无法全年生产,以及未能准确把握渔场渔期等是造成试捕亏损的主要原因[2]。
张鹏等[2]2010年分析了南海金枪鱼渔业和资源情况,认为兼捕作业或季节性生产是中国大陆开发南海金枪鱼资源的最好选择。由于黄鳍金枪鱼和鸢乌贼(Sthenoteuthis oualaniensis)之间存在着捕食关系,两者的渔场在时间和空间上基本一致,利用捕捞鸢乌贼的灯光罩网船进行延绳钓兼作,也许是开发南海金枪鱼资源较为可行的模式。
2010年以来,广东海洋大学与多家渔业公司合作,相继开展了8个航次的南海深水延绳钓调查和探捕。与南海以往的生产和调查结果相比,新一轮探捕的金枪鱼渔获率较低,未达到商业生产的要求[4];建议延绳钓船与灯光罩网船联合作业,船队捕获的金枪鱼由延绳钓船进行加工保鲜处理,认为由功能不同的渔船组队生产,更能取得规模效益[5]。其2010年7月航次的调查是利用加装了延绳钓渔具的灯光罩网渔船进行,证明罩网渔船开展延绳钓作业在技术上可行。但其延绳钓作业占用了灯光罩网的正常作业时间,且因为错过了鸢乌贼的生产旺季,探捕海域又偏离罩网中心渔场,调查船的渔获率较低[5]。
该研究是在灯光罩网生产旺季利用罩网渔船开展金枪鱼延绳钓兼作试验,在不影响灯光罩网正常作业的前提下,延绳钓作业也取得了相对较高的渔获率。文章介绍了该次捕捞试验情况,并对相关问题进行了分析,旨在探索出一种经济可行的金枪鱼捕捞新模式,推动南海大型金枪鱼资源的开发,提高渔船捕捞效益,维护国家海洋权益。
1. 材料与方法
1.1 试验船、作业时间和海域
试验船为广东电白县博贺渔港的“粤电渔42212”灯光罩网渔船,钢质,2012年2月建成,船长44.42 m,型宽7.80 m,型深4.30 m,设计排水量816.9 t。渔船主机2台,总功率318 kW,配500盏金属卤化物集鱼灯(×1 kW),发电机4台,总功率720 kW;包括船长在内船员11人。渔船配有速冻舱和保温舱,渔获冷冻保存,最低温度可到-18 ℃。
航次时间为2015年3月4日至4月30日,海上历时58 d,作业地点在10°36′N~12°52′N、114°11′E~114°43′E南沙群岛北部海域。
1.2 灯光罩网作业
罩网主尺度为303.00 m×87.66 m,沉子纲配重1 515 kg,网衣最大网目35 mm,网囊最小网目20 mm。渔船到达渔场后打开撑杆漂流,夜晚开灯诱鱼,视鱼群诱集情况放网。作业时首先通过4根穿过撑杆顶端的网角绳将网具拉开铺设在渔船下方,然后逐步调暗灯光将鱼群诱集至近表层,再同时松开4根网角绳放网罩扣捕捞。渔船作业一网耗时约30 min,每次放网后即开灯诱鱼等待下一网作业。渔船每晚的作业网次不一致,这主要取决于该晚的渔情和海况,通常每晚作业1~2网后,如果渔获率较低,渔船就会关灯休息,反之则会一直作业下去,直至天明。
1.3 金枪鱼延绳钓作业
延绳钓试验以不影响罩网的正常作业为前提。采用小型延绳钓渔具,主要由干线、支线、钓钩、浮子和无线电浮标等组成。干线采用台湾生产的尼龙编织绳,分段长度60 m;支线由硬质聚丙烯绳、锦纶单丝、细钢丝等3段通过转环连接而成,长度26 m;钓钩为环形钩;浮子绳长度18 m。两个浮子间装钩5枚,彼此的间距为60 m,每装配50支钓钩的干线为一筐。每次放钓时使用DST centi微型温深仪测量钓钩作业水层。渔船傍晚放钓,以船尾上甲板为操作平台,甲板人员4人,船速5.5 kn左右,所用饵料为罩网捕获的鸢乌贼;第二天早晨开始起钓,以渔船右前甲板为操作平台,甲板人员6人,船速3.1 kn左右,采用油压式卷扬机绞收干线,干线和支线人工整理后盘绕在筐内。
1.4 数据处理
数据统计处理采用Excel 2003软件。灯光罩网的渔获率按渔获质量计算,根据每天的渔获质量和开灯时长计算出单位光诱时间渔获量(kg · h-1)。延绳钓的渔获率按渔获质量和渔获尾数分别计算,参考李永振等[6]的定义,把按质量计算的渔获率称为CPUE(catch per unit effort,kg · 千钩-1);把按渔获尾数计算的渔获率称为上钩率(尾· 千钩-1)。
2. 结果与分析
2.1 灯光罩网作业及渔获组成
灯光罩网作业概况见表 1。渔船航次58 d,作业40晚,合计放网297次,开灯357.2 h,总渔获量129.635 t,总渔获率362.9 kg · h-1。作业日期分为2段,一是3月9日至3月29日(农历一月十九至二月初十),共计21晚,放网157次;二是4月6日至4月25日(农历二月十八至三月初七,4月14日晚因风浪大未作业),共计19晚,放网140次。因为月相对灯光罩网的渔获率有显著影响,满月前后罩网一般停止作业[7]。
表 1 灯光罩网作业概况Table 1. General information of light falling-net fishing夜晚
No.日期
date作业地点 position 网次
fishing times光诱时间
lighting time渔获量/kg
catch渔获率/kg·h-1
catch rateLat.(N) Long.(E) 1 0309 11°41′ 114°36′ 8 19:11~04:13 2 848 315.3 2 0310 11°42′ 114°37′ 8 19:07~05:13 1 702 168.5 3 0311 11°16′ 114°32′ 9 18:56~05:21 5 223 501.4 4 0312 11°16′ 114°32′ 8 19:04~05:33 3 236 308.7 5 0313 11°24′ 114°33′ 8 18:57~05:22 4 977 477.8 6 0314 11°24′ 114°33′ 9 18:56~05:36 5 609 525.8 7 0315 11°25′ 114°33′ 7 19:01~03:27 4 546 539.1 8 0316 11°32′ 114°37′ 6 19:00~03:17 1 416 170.9 9 0317 11°26′ 114°39′ 6 19:20~02:17 2 819 405.7 10 0318 11°41′ 114°36′ 8 19:00~05:36 2 206 208.1 11 0319 11°20′ 114°33′ 12 19:03~05:39 7 687 725.2 12 0320 11°20′ 114°33′ 10 19:00~05:29 6 504 620.4 13 0321 11°20′ 114°33′ 11 18:56~05:34 8 984 844.9 14 0322 11°20′ 114°33′ 6 19:02~03:07 1 862 230.3 15 0323 11°20′ 114°33′ 2 18:58~00:31 124 22.3 16 0324 11°44′ 114°36′ 9 19:04~05:35 2 650 251.9 17 0325 11°40′ 114°35′ 3 19:00~23:38 303 65.5 18 0326 11°52′ 114°33′ 2 19:01~22:34 182 51.3 19 0327 10°40′ 114°16′ 8 19:16~05:15 3 254 325.9 20 0328 10°40′ 114°16′ 8 19:06~04:58 1 304 132.2 21 0329 10°45′ 114°11′ 9 19:03~05:05 2 106 209.9 22 0406 11°28′ 114°39′ 1 19:10~20:46 91 56.9 23 0407 11°24′ 114°33′ 7 19:10~04:17 1 439 157.8 24 0408 11°21′ 114°30′ 8 18:58~05:31 5 043 478.0 25 0409 11°21′ 114°30′ 5 19:02~01:27 2 537 395.4 26 0410 11°47′ 114°39′ 8 19:15~05:32 4 590 446.4 27 0411 11°48′ 114°38′ 8 19:03~05:18 3 565 347.8 28 0412 11°47′ 114°38′ 10 18:59~05:10 5 281 518.6 29 0413 11°44′ 114°36′ 8 19:09~04:45 4 545 473.5 30 0415 11°44′ 114°37′ 10 18:59~04:41 3 969 409.2 31 0416 11°43′ 114°36′ 4 19:05~00:20 634 120.7 32 0417 11°22′ 114°31′ 8 19:05~05:05 3 316 331.6 33 0418 11°47′ 114°37′ 10 19:04~05:08 4 787 475.5 34 0419 11°40′ 114°43′ 9 19:08~05:19 3 679 361.3 35 0420 11°47′ 114°37′ 9 19:03~05:10 4 083 403.6 36 0421 11°33′ 114°43′ 8 19:04~04:36 3 539 371.2 37 0422 11°29′ 114°38′ 8 19:02~04:58 3 435 345.8 38 0423 11°36′ 114°40′ 7 19:01~03:31 3 186 374.8 39 0424 11°47′ 114°37′ 5 19:15~02:41 1 119 150.5 40 0425 12°52′ 114°14′ 7 19:17~04:26 1 255 137.2 渔船19点左右开灯诱鱼,每晚放网1~12次,平均7.4次;开灯1.60~10.67 h,平均8.9 h;渔获量0.091~8.984 t,平均3.241 t;渔获率22.3~844.9 kg · h-1,平均336.4 kg · h-1。通常渔获率高的夜晚,渔船作业网次较多,捕捞产量也相应较高。
灯光罩网有记录的渔获达46种,渔获质量居前10位的种类见表 2。鸢乌贼为绝对优势渔获,占总渔获质量的85.31%;扁舵鲣(Auxis thazard)、鲔(Euthynnus affinis)、鲣(Katsuwonus pelamis)、圆舵鲣(A.rochei) 等属小型金枪鱼类,合计占总渔获质量的5.29%。该航次罩网共捕获大型金枪鱼8尾,都为黄鳍金枪鱼,体长范围103~138 cm,平均118.5 cm,体质量范围25~48 kg,平均34.1 kg,总质量273 kg,占总渔获量的0.21%,加上捕获的幼鱼,黄鳍金枪鱼在总渔获质量中居第9位。相比该船前二年同期的生产情况[8],该航次的渔获率略低,渔获组成基本一致。
表 2 灯光罩网居前10位的渔获组成Table 2. Catch composition of falling-net top 10 species排名
No.种类
species质量/kg
weight质量百分比/%
ratio of weight1 鸢乌贼 Sthenoteuthis oualaniensis 110 595.50 85.31 2 细鳞圆鲹 Decapterus macarellus 10 856.55 8.37 3 扁舵鲣 Auxis thazard 3 050.54 2.35 4 鲔 Euthynnus affinis 1 703.35 1.31 5 鲣 Katsuwonus pelamis 1 374.41 1.06 6 圆舵鲣 Auxis rochei 727.99 0.56 7 斑条魣 Sphyraena jello 315.10 0.24 8 黑带鳞鳍梅鲷 Pterocaesio tile 312.09 0.24 9 黄鳍金枪鱼 Thunnus albacares 275.43 0.21 10 鳞首方头鲳 Cubiceps squamiceps 252.61 0.19 2.2 延绳钓作业及渔获组成
延绳钓作业概况见表 3。延绳钓试验是在3月30日至4月7日(农历二月十一至十七的满月期间)进行,渔船傍晚放钓,每百钩耗时36~45 min,平均39 min;第二天早晨起钓,每百钩耗时69~103 min,平均84 min。作业7次,合计放钩2 700枚,总渔获数量69尾,总渔获质量1 281.6 kg,总上钩率为25.56尾· 千钩-1,总CPUE为474.67 kg · 千钩-1。延绳钓不同钓次间的上钩率和CPUE的波动较大,平均上钩率为25.74尾· 千钩-1,平均CPUE为427.48 kg · 千钩-1。
表 3 金枪鱼延绳钓作业概况Table 3. General information of tuna longline fishing钓次
No.放钩数量
number of hooks放钓位置
setting position放钓日期
setting date放钓时间
setting time起钓日期
retrieving date起钓时间
retrieving time尾数
quantity上钩率/尾· 千钩-1
catch rate质量/kg
weightCPUE/kg· 千钩-1 Lat.(N) Long.(E) 1 300 11°20′ 114°31′ 0330 16:56~19:01 0331 06:38~10:31 9 30.00 69.4 231.17 2 300 11°33′ 114°37′ 0331 17:22~19:36 0401 06:40~10:45 13 43.33 98.7 329.00 3 400 11°03′ 114°30′ 0401 16:56~19:21 0402 06:07~12:42 17 42.50 220.7 551.75 4 400 10°57′ 114°25′ 0402 17:03~19:48 0403 06:37~13:30 10 25.00 326.6 816.50 5 500 10°36′ 114°15′ 0403 16:34~19:49 0404 06:08~12:44 8 16.00 336.5 673.00 6 600 10°44′ 114°14′ 0404 17:12~20:46 0405 06:29~14:27 11 18.33 227.4 378.92 7 200 11°24′ 114°33′ 0406 15:59~17:17 0407 06:43~09:00 1 5.00 2.4 12.00 延绳钓有记录的渔获10种(表 4),其中,剑鱼(Xiphias gladius)、异鳞蛇鲭(Lepidocybium flavobrunneum)、大鳞乌鲂(Taractichthys steindachneri)、帆蜥鱼(Alepisaurus ferox)为延绳钓独有渔获,没有在罩网渔获中出现。剑鱼和黄鳍金枪鱼是国际市场上的高档水产品[9],合计占到总渔获质量的84.04%,平均上钩率和CPUE合计达5.67尾· 千钩-1、344.60 kg · 千钩-1。黄鳍金枪鱼渔获体长范围92~124 cm,平均109.8 cm,体质量范围17~40 kg,平均28.6 kg;剑鱼渔获体长范围132~247 cm,平均212.8 cm,体质量范围9~110 kg,平均77.8 kg。其他种类的经济价值相对较低,黑(Dasyatis atratus)、蛇鲭(Gempylus serpens)等直接在海上抛弃。另外,有1尾黄鳍金枪鱼和1尾鲨鱼在起钓过程中逃逸,未纳入统计。
表 4 延绳钓渔获物概况Table 4. Catch composition of longline fishing种类
species质量
weight尾数
quantity上钩率/尾·千钩-1
catch rateCPUE/kg·千钩-1 kg % ind. % mean range mean range 剑鱼 Xiphias gladius 934 72.88 12 17.39 3.76 0~8.00 291.86 0~775.00 黄鳍金枪鱼 Thunnus albacares 143 11.16 5 7.25 1.90 0~5.00 52.74 0~122.50 斑条魣 Sphyraena jello 55 4.29 4 5.80 1.90 0~10.00 26.19 0~133.33 异鳞蛇鲭 Lepidocybium flavobrunneum 53.2 4.15 5 7.25 1.90 0~3.33 23.54 0~66.67 蛇鲭 Gempylus serpens 39.3 3.07 28 40.58 11.19 0~16.67 15.94 0~31.17 鲯鳅 Coryphaena hippurus 19.5 1.52 2 2.90 0.57 0~4.00 5.57 0~39.00 大鳞乌鲂 Taractichthys steindachneri 13 1.01 3 4.35 1.00 0~3.33 4.29 0~13.33 黑魟 Dasyatis atratus 12.2 0.95 4 5.80 1.24 0~5.00 3.63 0~12.50 刺鲅 Acanthocybium solandi 7 0.55 1 1.45 0.24 0~1.67 1.67 0~11.67 帆蜥鱼 Alepisaurus ferox 5.5 0.43 5 7.25 2.02 0~7.50 2.06 0~11.25 3. 讨论
3.1 南海金枪鱼延绳钓渔业
南海历史上曾是日本和中国台湾延绳钓船的主要作业渔场之一。日本钓船20世纪30年代以前就到南沙海域生产,60~70年代达到鼎盛期,1967年金枪鱼产量达6 568 t[10],目前已经没有钓船在南海作业;台湾钓船1979年金枪鱼和剑旗鱼产量达28 919 t (金枪鱼24 834 t),2012年仅为14 601 t (金枪鱼8 175 t) [11],渔船主要在秋冬季节到南海生产[12]。越南延绳钓渔业1991年起步后发展较为迅速[2],2012年金枪鱼和剑旗鱼产量已达16 632 t [3],但越南渔船很多只是季节性从事延绳钓作业[13],且很多延绳钓船目前已经改为灯光手钓作业。这是越南近年来兴起的一种新型作业方式,渔船装有几个至十几个集鱼灯,伸出4根木杆,杆上各挂1根钓线,系2个金枪鱼钩,配3~4 kg重沉,傍晚放钓,使用鱿鱼饵料;渔船夜晚开灯诱鱼,船员一边从事鸢乌贼手钓,一边等待金枪鱼上钩。相比延绳钓渔业,灯光手钓金枪鱼产量更高,且几乎可以全年生产,但其满月期间无法作业[3, 14]。越南平定省的507艘延绳钓船2011年底开始全部转为灯光手钓作业,2012年灯光手钓渔船已达1 060艘[14]。因此,相比20世纪60~70年代而言,南海的金枪鱼延绳钓渔业总体上是呈萎缩态势的。
中国大陆2010年~2013年8个航次的深水延绳钓探捕也显示,南海大型金枪鱼的资源密度较低,在南海劳动力和生产成本日益高企的今天,延绳钓作业难以取得经济效益,其平均渔获率和CPUE分别为20.95尾· 千钩-1和177.17 kg · 千钩-1 [4]。该研究中延绳钓的平均上钩率和CPUE相对较高,分别为25.74尾· 千钩-1和427.48 kg · 千钩-1,其中金枪鱼和剑旗鱼的平均上钩率和CPUE分别为5.67尾· 千钩-1和344.60 kg · 千钩-1,略高于越南平定省延绳钓渔业300 kg · 千钩-1的CPUE平均水平[14],但远低于20世纪50~60年代日本钓船在相同海域33.1尾· 千钩-1的平均上钩率[10],说明南海目前的大型金枪鱼资源远没有历史上丰富。越南金枪鱼钓业的快速发展,与其距离渔场近、生产成本低有关[13-14]。资料显示,越南延绳钓和手钓渔船,一个航次20~30 d,生产成本(3.3~4.2)万元,捕捞产值(4.5~7.5)万元,船员收入1 500~2 100元[14]。
因为金枪鱼类高度洄游的特性,南海延绳钓渔业的中心渔场存在明显季节变化,不同年度和月份的上钩率也有较大波动[12, 14]。专业延绳钓渔船投资和运营成本高,捕捞效益无法保证,且面临着生产淡季渔船的去向问题。利用其他作业类型渔船兼捕,或者组织远洋钓船季节性到南海生产,是中国大陆发展南海金枪鱼渔业较理性的选择。
3.2 罩网渔船兼作延绳钓模式
鸢乌贼灯光罩网渔业是近年来南海外海渔业发展的热点。渔船早期在离港较近的西中沙海域生产[2, 15],2011年作业渔场拓展至南沙海域[16],2013年仅广西就有65艘灯光罩网渔船获准前往南沙生产[17]。相比其他作业类型渔船而言,利用罩网渔船开展延绳钓兼作优势明显。
1) 金枪鱼延绳钓和灯光罩网的渔场分布恰好一致。因为食物链关系,罩网渔船夜晚捕捞鸢乌贼时,经常罩捕到前来摄食的大型金枪鱼类,西中沙和南沙海域罩网渔获中,大型金枪鱼都占有一定比例[8, 15]。该研究中灯光罩网捕获大型黄鳍金枪鱼8尾(273 kg),延绳钓没有耗时另找渔场,而是在罩网渔场作业,也取得相对较高的上钩率,证明两者的渔场在时空分布上基本一致。
2) 金枪鱼延绳钓和灯光罩网的作业时间没有冲突。灯光罩网是夜晚作业,白天休息,月暗夜作业,满月时休息(表 1)。该研究中,延绳钓试验是在农历二月十一至十七的满月期间进行,这段时间灯光罩网正好无法作业,而渔民普遍认为农历十五前后月光较强,下钓生产可以求得较好的产量[18]。另外,从作业时段来看,该研究中延绳钓是傍晚放钓,第二天早晨起钓,没有占用罩网作业时段,只要人员搭配合理,渔船可每天开展2种作业。4月6日渔船就是天黑前完成放钓,夜晚从事罩网作业,第二天早晨起钓。
新建钢质渔船目前已经成为南海外海罩网渔业生产的主力[8],其一个航次将近2个月,可以利用满月和白天空闲时间开展延绳钓作业。罩网渔船完成加装钓具的改造后,只要渔获物能够抵消起放钓时的油耗和人工成本,延绳钓作业就有利可图。相比发展专业钓船而言,罩网渔船兼作延绳钓模式具有投资金额少、生产成本低的优势,渔船可以根据海域资源和渔获价格变动情况,灵活安排2种作业的时间和比重,有助于提高渔船的捕捞效益和抗风险能力。
3.3 存在问题和发展建议
该研究为了降低渔船改造成本,延绳钓采用传统小型钓具,只在起钓时使用卷扬机,干线和支线必须用人工整理盘绕在筐内,放钩数量少,甲板工作量大。远洋钓船目前已经普遍采用滚筒式钓机,每天放钩3 000枚左右,效率高且省人工[19]。南海新建罩网渔船的长度普遍超过40 m,有足够的空间加装滚筒式钓机,以大幅提高延绳钓的作业效率。因为罩网自身就捕获不少金枪鱼,加上渔船可兼作延绳钓,目前制约南海金枪鱼渔业发展的瓶颈问题已经不在捕捞技术,而在于以下2个方面。
1) 金枪鱼的海上保鲜问题。南海外海罩网渔业目前仍以个体经营为主,渔船主要依靠自身解决后勤补给,没有外海渔业基地或冷藏运输船配套。因航次时间较长,装备相对落后,导致渔船回港后,金枪鱼普遍无法达到生食标准,只能当作普通渔获物出售[20]。以该研究为例,合计捕获大型黄鳍金枪鱼416 kg、剑鱼934 kg,回港后售价分别为14元· kg-1和8元· kg-1,而同期远洋深冷黄鳍金枪鱼和剑鱼的收购价分别为10美元· kg-1和4.5美元· kg-1,价格相差数倍。
2) 外海渔期相对短暂问题。南海外海罩网渔业目前仍属季节性作业,渔船主要在2月~6月间到南海中南部深水区作业,其他时间则在南海北部陆架区生产[8],渔船只有在外海作业时才能捕到大型金枪鱼,目前每年渔期只有4个月左右。因渔期相对较短,投资成本较高,导致渔民没有积极性改进金枪鱼保鲜技术和装备。目前,南海区仅海南省东方市的一艘新建罩网渔船投资160多万元安装了超低温速冻和冷藏舱。
台湾钓船主要在秋冬季节到南海生产,秋季渔船主要在120°E~121°E、19°N~21°N海域作业,冬季则散布于115°E~119°E、13°N~20°N海域[12]。越南钓船每年11月~4月主要在14°00′N~16°30′N、112°00′E~115°00′E海域作业,此时金枪鱼的产量高于其他月份,尤其以12月~2月的产量最高[14]。中国大陆2012年9月~10月的罩网探捕也发现,秋季南海外海依然有鱼可捕,中心渔场位于12°30′N~15°30′N、110°30′E~115°00′E海域,此时除了鸢乌贼和大型金枪鱼类以外,小型金枪鱼类资源也较丰富[21]。上述资料表明,秋冬季节才是南海金枪鱼渔业的作业旺季,且中心渔场位于离中国港口较近的西中沙海域,而11月~2月恰好是南海北部陆架海域灯光罩网渔业的生产淡季[15],这非常有助于罩网渔船兼作金枪鱼延绳钓模式的推广。
建议加强南海金枪鱼船载保鲜与钓捕技术装备的集成研究,加快南海外海渔业基地与冷链物流体系的建设,加强南海深水区渔业资源的调查和探捕,解决金枪鱼保鲜问题,不断延长外海渔期,有力推动南海外海金枪鱼和灯光罩网渔业的发展。
-
![]()
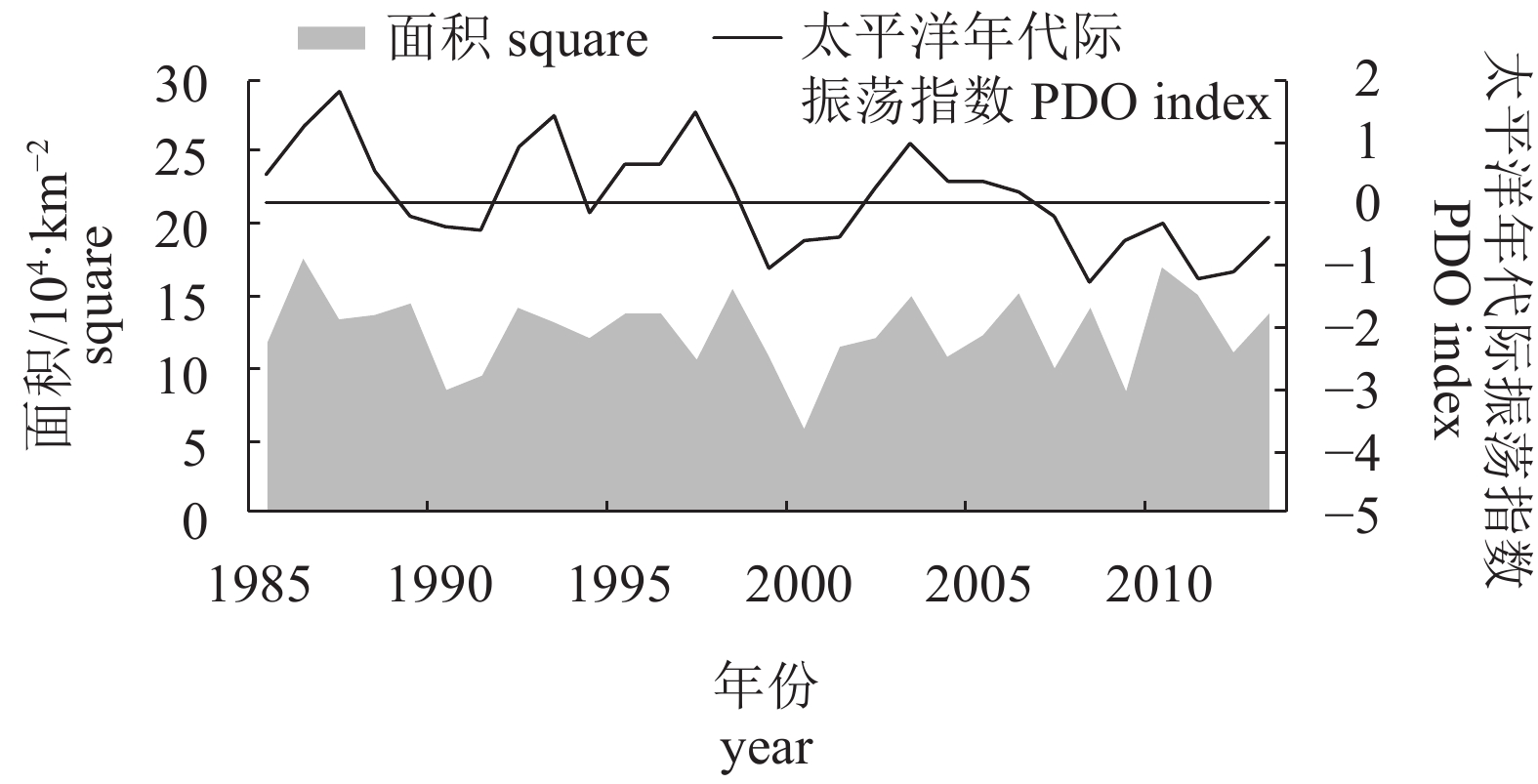
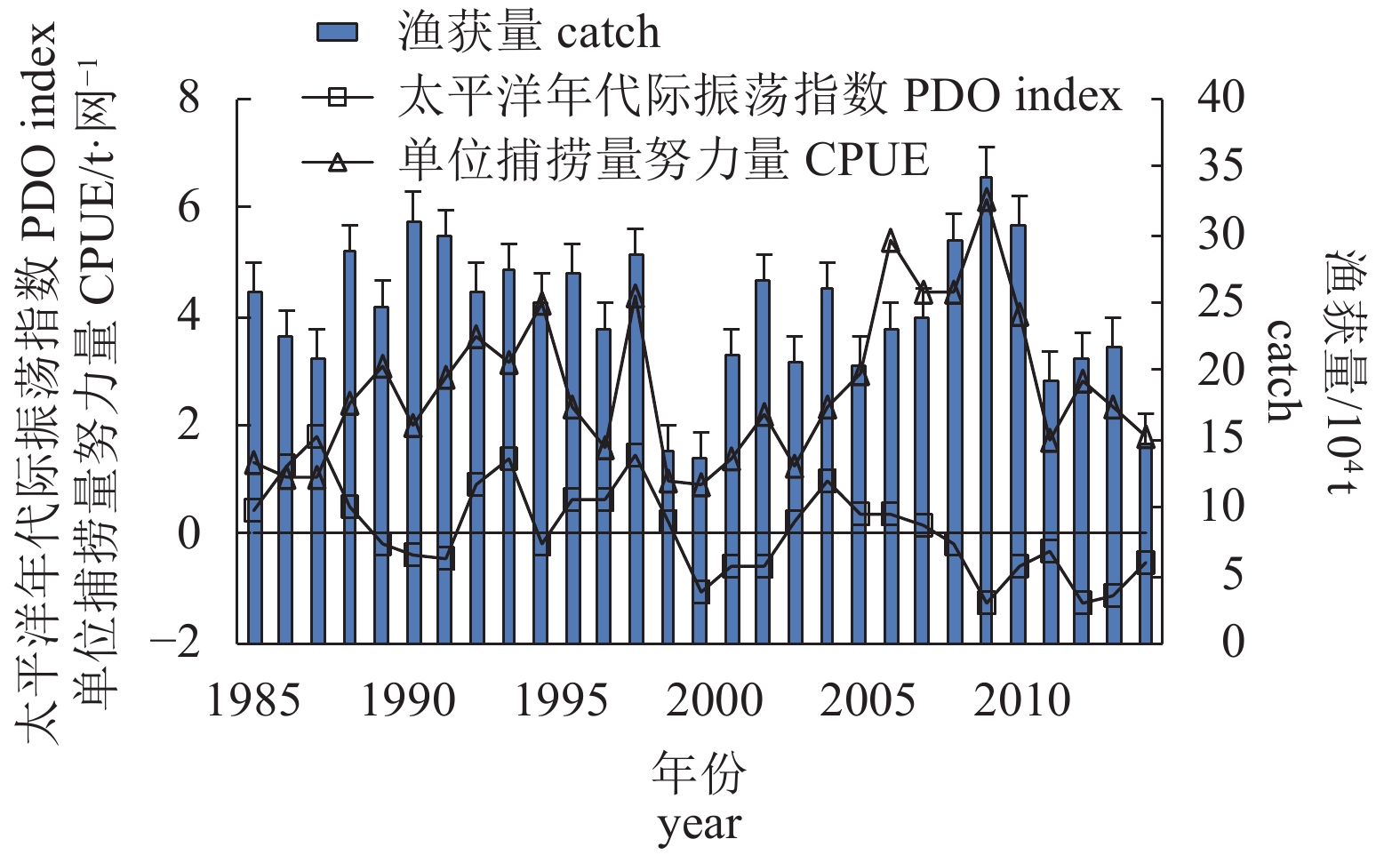
图 1 日本1985—2013年秋刀鱼渔获量、太平洋年代际振荡指数及单位捕捞量努力量变化
PDO为正数代表气候模态处于暖期,为负数代表处于冷期
Figure 1. Variation of saury catch, PDO index and CPUE in Japan from 1985 to 2013
Positive PDO index indicates that the climate is in warm period, while negative PDO index indicates that the climate is in cold period.
![]()
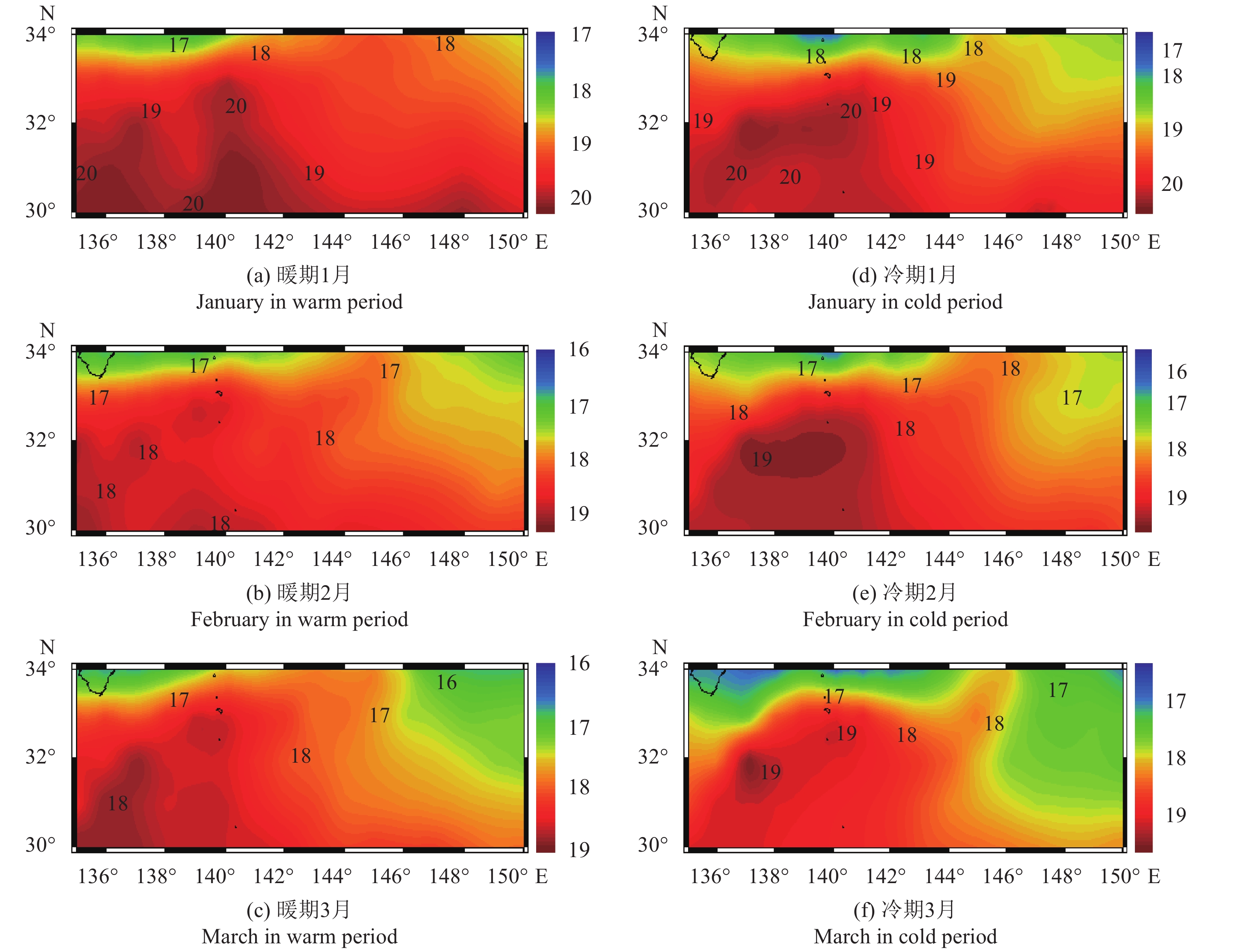
图 2 产卵场内太平洋年代际振荡冷、暖位相下月平均海表温度分布
Figure 2. Monthly mean SST distribution in cold and warm periods of PDO in spawning ground
![]()
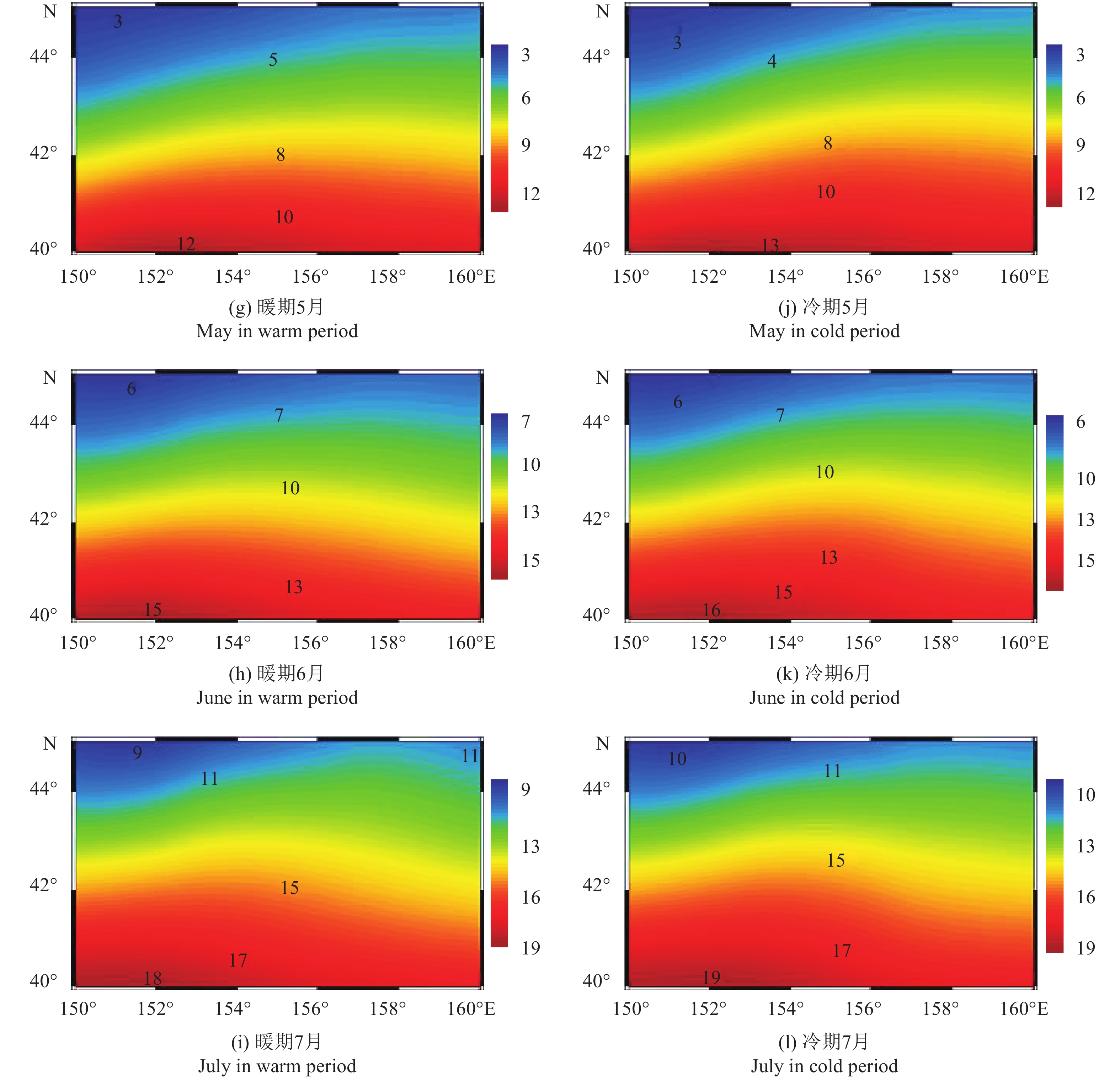
图 3 索饵场内太平洋年代际振荡冷、暖位相下月平均海表温度分布
Figure 3. Monthly mean SST distribution in cold and warm periods of PDO in feeding ground
![]()
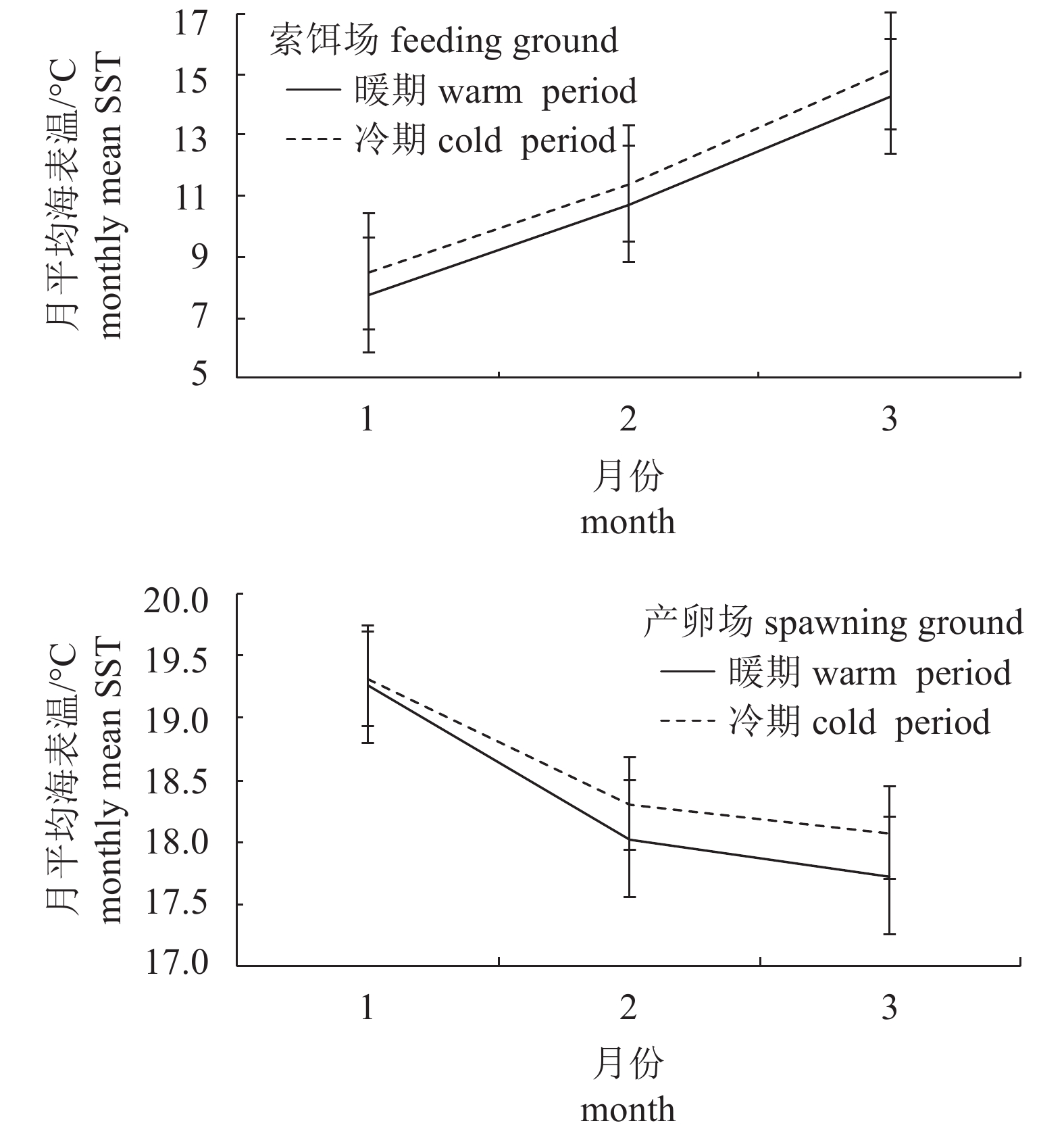
图 4 索饵场、产卵场月平均海表温度随时间变化趋势
Figure 4. Variation trends of monthly mean SST in feeding and spawning ground along with time
![]()
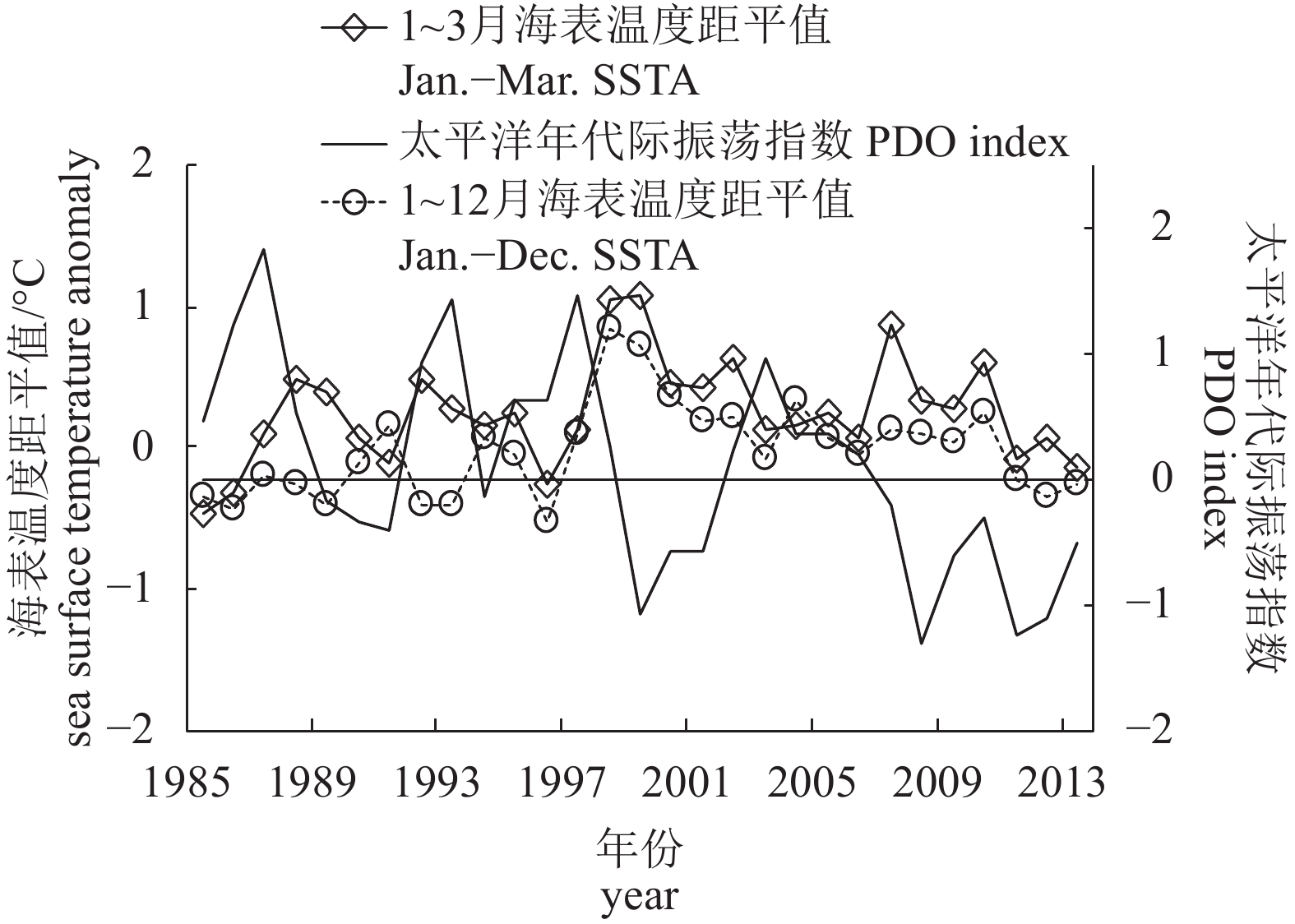
图 5 1985—2013年产卵场平均海表温度距平值与太平洋年代际振荡指数关系
Figure 5. Relationship between mean SSTA and PDO index of spawning ground from 1985 to 2013
![]()
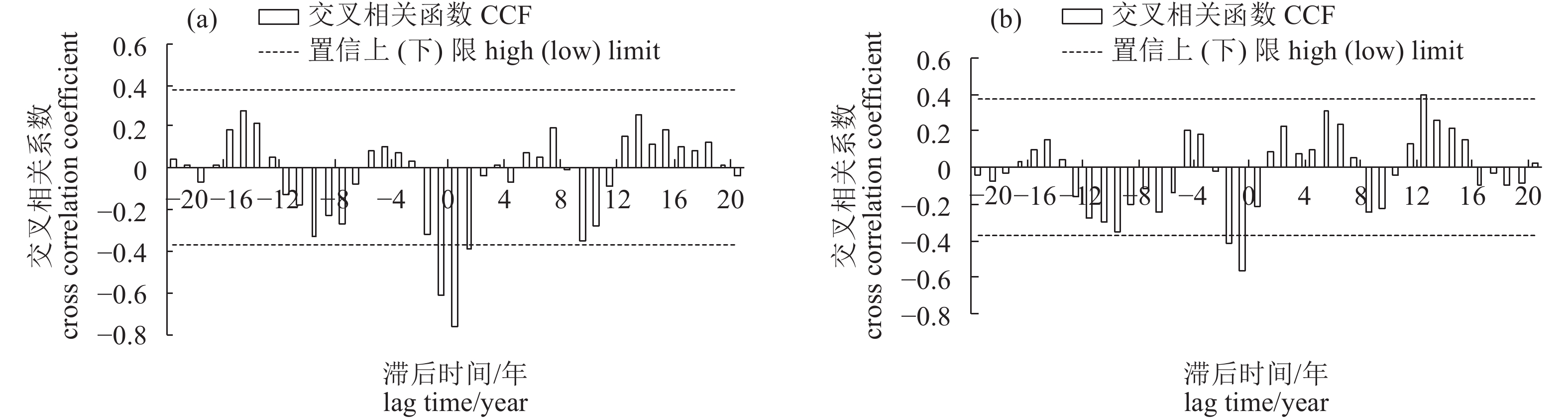
图 6 太平洋年代际振荡指数与1—12月 (a)、1—3月 (b) 海表温度距平值交相关系数
Figure 6. Cross correlation coefficients between PDO index and SSTA from January to December (a) and January to March (b)
![]()
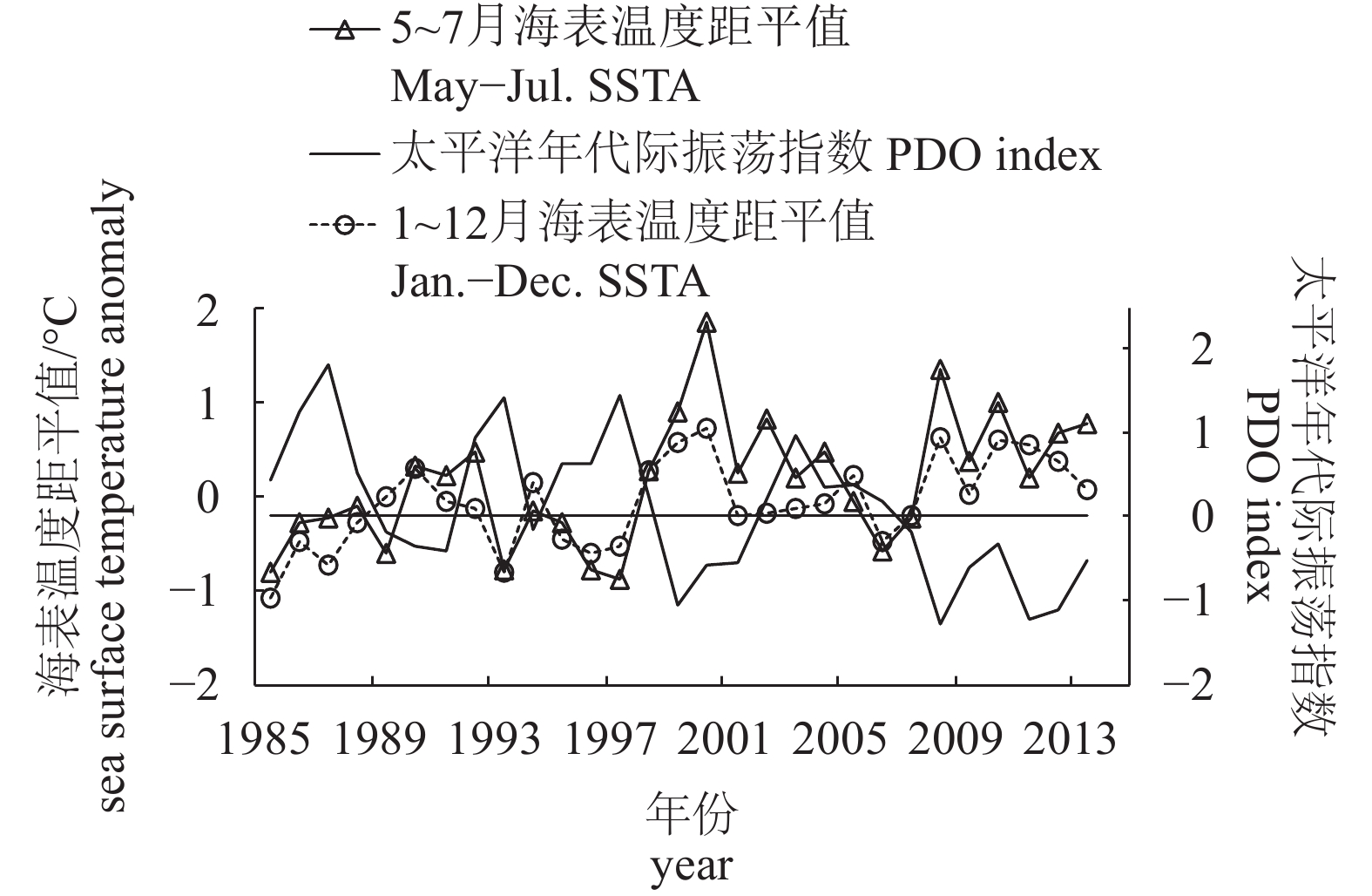
图 7 1985—2013年索饵场平均海表温度距平值与太平洋年代际振荡指数关系
Figure 7. Relationship between mean SSTA and PDO index in feeding ground from 1985 to 2013
![]()
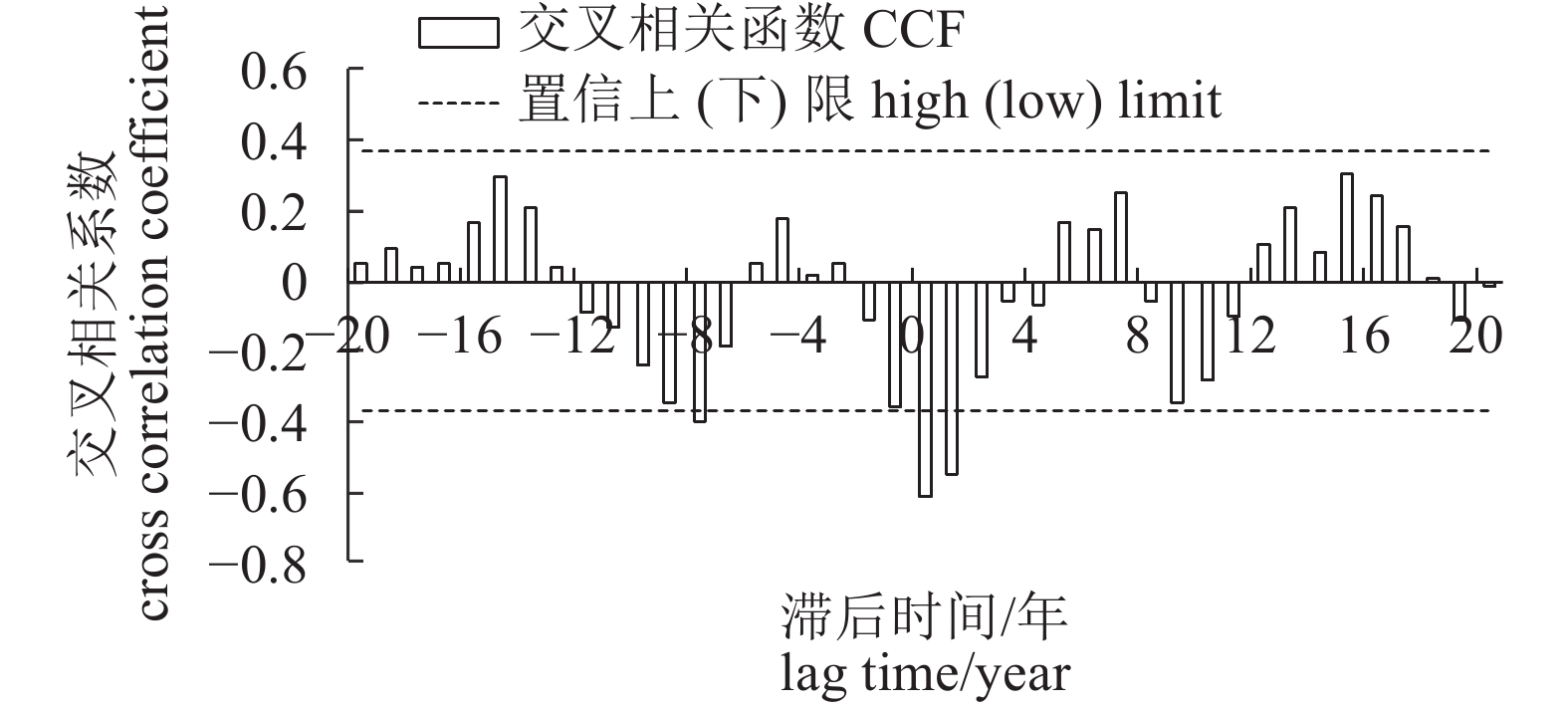
图 8 太平洋年代际振荡指数与5—7月海表温度距平值交叉相关系数
Figure 8. Cross correlation coefficients between PDO index and SSTA from May to July
![]()
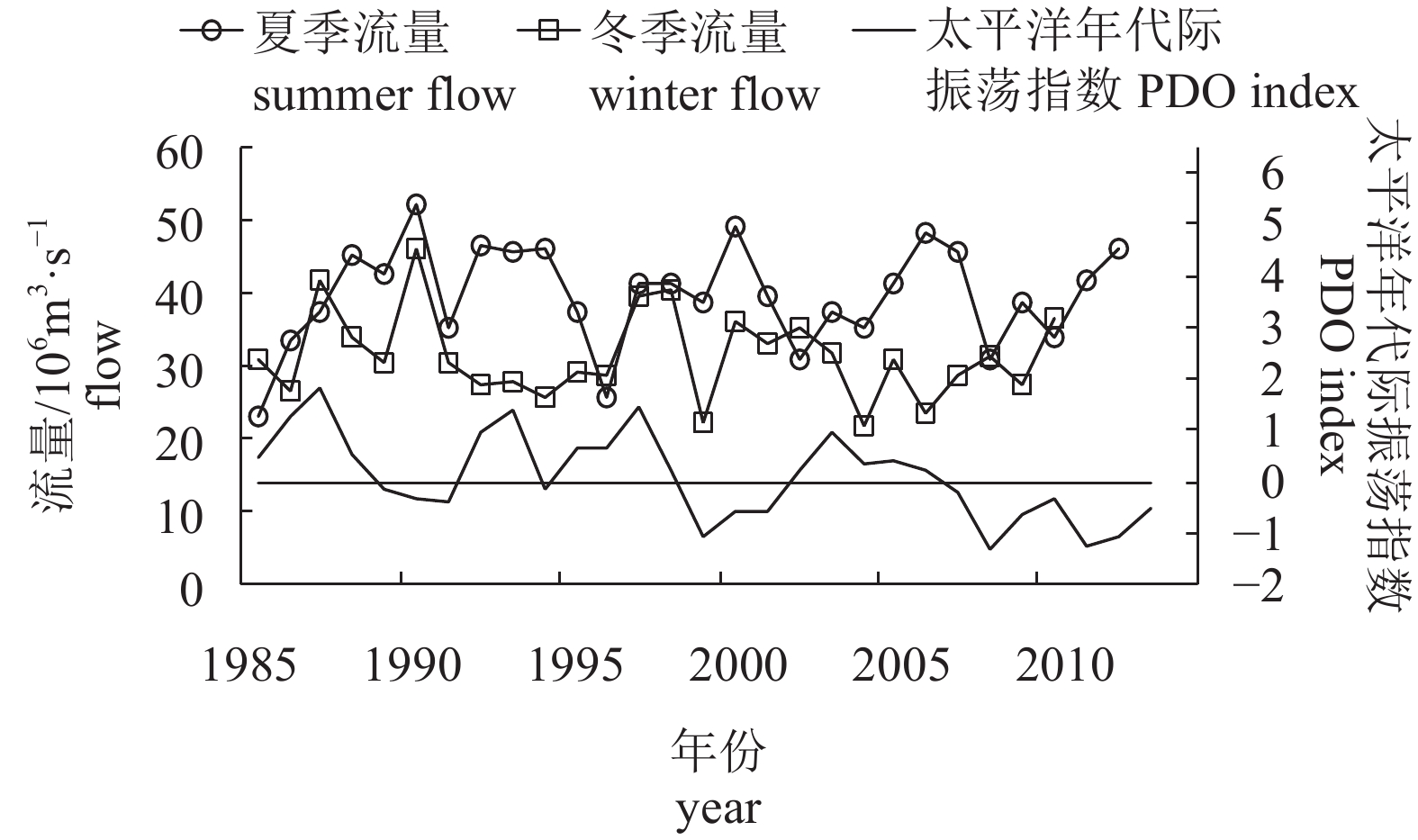
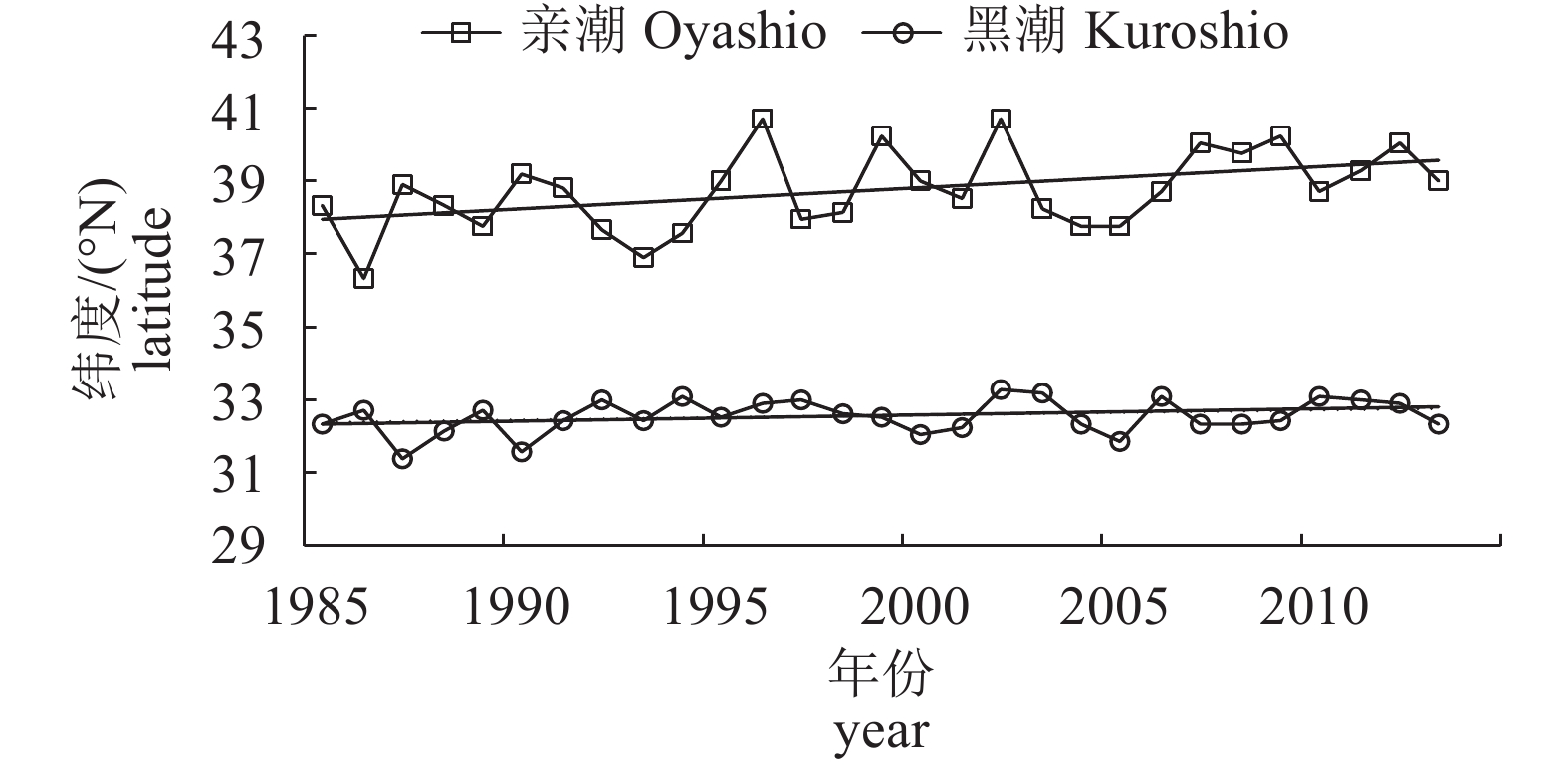
图 9 1985—2013年黑潮夏、冬两季流量与太平洋年代际振荡指数关系
Figure 9. Relationship between Kuroshio transport and PDO index during summer and winter from 1985 to 2013
![]()
图 10 1985—2013年亲潮春季平均占有面积与太平洋年代际振荡指数关系
Figure 10. Relationship between average occupied area in spring of Oyashio and PDO index from 1985 to 2013
-
[1] 商李磊, 朱清澄, 晏磊, 等. 西北太平洋公海秋刀鱼生物学特性研究[J]. 江苏农业科学, 2012, 40(10): 198-201. [2] SUYAMA S, KURITA Y, UENO Y. Age structure of Pacific saury Cololabos saira based on observations of the hyaline zones in the otolith and length frequency distributions[J]. Fish Sci, 2006, 72: 742-749.
[3] 朱清澄, 高玉珍, 花传祥, 等. 西北太平洋秋刀鱼耳石透明带的初步研究[J]. 海洋渔业, 2016, 38(3): 236-244. [4] 吴越, 黄洪亮, 刘健, 等. 2004-2011年中国台湾秋刀鱼作业渔场年代际变化初步分析[J]. 渔业信息与战略, 2014, 29(4): 263-271. [5] TIAN Y J, UENO Y, SUDA M, et al. Climate-ocean variability and the response of Pacific saury (Cololabis saira) in the northwestern Pacific during the last half century[J]. Fish Sci, 2002, 68(1): 158-161.
[6] 张孝民, 朱清澄, 花传祥. 2013年北太平洋公海秋刀鱼渔场与海洋环境的关系[J]. 上海海洋大学学报, 2015, 24(5): 773-782. [7] MATTSUMIYA Y, TANAKA S. 東北·北海道沖の南下サンマの資源動態についてⅢ[J]. 日本水産學會誌, 1978, 44(5): 451-455. [8] 谷德军, 王东晓, 李春晖. PDO源地与机制的若干争论[J]. 热带气象学报, 2003, 19(z1): 136-144. [9] 方海, 张衡, 刘峰, 等. 气候变化对世界主要渔业资源波动影响的研究进展[J]. 海洋渔业, 2008, 30(4): 363-370. [10] MANTUA N J, HARE S R, ZHANG Y, et al. A Pacific interdecadal climate oscillation with impacts on salmon production[J]. B Am Meteorol Soc, 1997, 78(6): 1069-1079.
[11] CHEN X J, CAO J, CHEN Y, et al. Effect of the Kuroshio on the spatial distribution of the red flying squid Ommastrephes bartramii in the Northwest Pacific Ocean[J]. B Mar Sci, 2012, 88(1): 63-71.
[12] TIAN Y J, UENO Y, SUDA M, et al. Decadal variability in the abundance of Pacific saury and its response to climate/oceanic regime shifts in the northwestern subtropical Pacific during the last half century[J]. J Mar Syst, 2004, 52: 235-257.
[13] 沈建华, 韩士鑫, 樊伟, 等. 西北太平洋秋刀鱼资源及其渔场[J]. 海洋渔业, 2004, 26(1): 61-65. [14] 朱清澄, 花传祥, 许巍, 等. 西北太平洋公海7~9月秋刀鱼渔场分布及其与水温的关系[J]. 海洋渔业, 2006, 28(3): 228-233. [15] TSENG C T, SUN C L, BELKIN I M, et al. Sea surface temperature fronts affect distribution of Pacific saury (Cololabis saira) in the Northwestern Pacific Ocean[J]. Deep-Sea Res Ⅱ, 2014, 107: 15-21.
[16] TIAN Y J, AKAMINE T, SUDA M. Variations in the abundance of Pacific saury (Cololabis saira) from the northwestern Pacific in relation to oceanic-climate changes[J]. Fish Res, 2003, 60(2/3): 439-454.
[17] 孙满昌, 叶旭昌, 张健, 等. 西北太平洋秋刀鱼渔业探析[J]. 海洋渔业, 2003, 25(3): 112-115. [18] WATANABE Y. Growth and surival of Pacific saury Cololabis saira in the Kuroshio-Oyashio transitional waters[J]. J Oceanogr, 2003, 59: 403-414.
[19] BERTRAND A, JOSSE E, BACH P, et al. Hydrological and trophic characteristics of tuna habitat: consequences on tuna distribution and longline catchability[J]. Can J Fish Aquat Sci, 2002, 59(6): 1002-1013.
[20] TAKASUKA A, NISHIKAWA K, KURODA H A, et al. Growth variability of Pacific saury Cololabis saira larvae under contrasting environments across the Kuroshio axis: survival potential of minority versus majority[J]. Fish Oceanogr, 2016, 25(4): 390-406.
[21] 张孝民.西北太平洋公海秋刀鱼渔场研究[D]. 上海:上海海洋大学, 2016:1-4. [22] 田思泉, 陈新军, 冯波, 等. 西北太平洋柔鱼资源丰度与栖息环境的关系及其时空分布[J]. 上海海洋大学报, 2009, 18(5): 586-592. [23] 余为, 陈新军, 易倩. 不同气候模态下西北太平洋柔鱼渔场环境特征分析[J]. 水产学报, 2017, 41(4): 525-534. [24] Tohoku National Fisheries Research Institute. Stock assessment and estimation of the acceptable biological catch in 2015 for the Pacific saury (Cololabis saira) in the Northwestern Pacific[R]. Miyagi: Fisheries Research Agency, 2015: 8-9.
[25] 朱清澄, 杨明树, 高玉珍, 等. 西北太平洋秋刀鱼耳石生长与性成熟度、个体大小的关系[J]. 上海海洋大学学报, 2017, 16(2): 263-270. [26] 田永军. 北西太平洋におけるサンマ資源の長期変動特性と気候変化[J]. 水産海洋研究, 2012, 66(1): 16-25. [27] 谢斌, 汪金涛, 陈新军, 等. 西北太平洋秋刀鱼资源丰度预报模型构建比较[J]. 广东海洋大学学报, 2015, 35(6): 58-63. [28] 王闪闪. 黑潮、厄尔尼诺-南方涛动和太平洋年代际涛动的相互联系及对气候影响的研究[D]. 兰州:兰州大学, 2015:5-10. [29] GONG Y, SUH Y S. Effect of climate-ocean changes on the abundance of Pacific saury[J]. J Environ Biol, 2013, 34: 23-30.
[30] BAITALIUK A A, ORLOV A M, ERMAKOV Y K. Characteristic features of ecology of the Pacific saury Cololabis saira (Scomberesocidae, Beloniforms) in open waters and in the Northeast Pacific ocean[J]. J Ichthyol, 2013, 53(11): 899-913.
[31] OOZEKI Y, WATANABE Y, KITAGAWA D. Environmental factors affecting larval growth of Pacific saury, Cololabis saira, in the northwestern Pacific Ocean[J]. Fish Oceanogr, 2004, 13(1): 44-53.
-
期刊类型引用(5)
1. 王春丽,陈峰,蒋日进,张洪亮,朱文斌,朱凯. 典型气候事件对浙江近海底层鱼类时空分布的影响. 生态学报. 2024(10): 4231-4243 .  百度学术
百度学术
2. 白思琦,邹晓荣,张鹏,丁鹏. 环境因子对东南太平洋智利竹?鱼渔场时空分布异质性影响. 南方水产科学. 2021(01): 17-24 . 本站查看
3. 梁佳伟,陈新军,花传祥,胡贯宇,方舟,赵振方. 秋刀鱼渔业资源与渔场学研究进展——基于Citespace分析. 海洋湖沼通报. 2021(03): 117-128 . 百度学术
4. 王靓,花传祥,朱清澄,李非. 北太平洋小型中上层鱼类资源对气候–海洋变化的响应研究进展. 中国水产科学. 2020(11): 1379-1392 . 百度学术
5. 花传祥,朱清澄,许巍,宋利明,李非. 北太平洋秋刀鱼生活史和资源渔场研究进展. 中国水产科学. 2019(04): 811-821 . 百度学术
其他类型引用(3)

 下载:
下载:
计量
- 文章访问数: 3039
- HTML全文浏览量: 1036
- PDF下载量: 492
- 被引次数: 8
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
