
Acoustic assessment of fishery resources and spatial distribution in Nan'ao Island area
-
摘要: 基于2014年秋季和2015年春季南澳岛海域渔业资源声学与底拖网调查数据,对该海域主要渔获物种类、资源动态与空间分布进行了评估分析。结果表明优势种类存在较大差异,短尾大眼鲷(Priacanthus macracanthus)和鹿斑鲾(Secutor ruconius)分别为秋季和春季主要优势鱼类。秋季和春季声学评估种类平均数量和生物量密度分别为(26 806±39 968) 尾·n mile–2和(403.68±601.9) kg·n mile–2、(34 416±38 497)尾·n mile–2和(124.5±139.26) kg·n mile–2。统计分析结果显示,2014年秋季渔获组成单一和混合2种不同条件下,调查海域渔业资源数量与生物量密度均存在显著性差异(P=0.007<0.05;P=0.004<0.05);而2015年春季不同条件下差异均不显著(P=0.113>0.05;P=0.310>0.05)。该差异可能与不同季节声学评估种类优势集中程度相关。由于鹿斑鲾(Secutor ruconius)等种类季节性集群活动的影响,调查海域渔业资源在水平空间上分布极不均匀。此外,以15 m水深为界,不同季节回波单体目标强度(TS)频率组成及其在垂直方向上的分布存在明显差异。Abstract: Based on the acoustics and bottom trawl survey data of fishery resources in the Nan'ao Island area in autumn 2014 and spring 2015, we analyzed the species composition, dynamic and spatial distribution of fishery resources in that area. The dominant species vaired in different seasons. Priacanthus macracanthus was dominant in autumn and Secutor ruconius was dominant in spring. The average quantity and biomass densities of species assessed by acoustic detection were (26 806±39 968) ind·n mile–2 and (403.68±601.9) kg·n mile–2 in autumn 2014, and (34 416±38 497) ind·n mile–2 and (124.5±139.26) kg·n mile–2 in spring 2015, respectively. Significant difference existed in the quantity and biomass density of fishery resources (P=0.007<0.05; P=0.004<0.05) in single or mixed-fish species captured in autumn 2014. However, the difference in spring 2015 was not significant (P=0.113>0.05; P=0.310>0.05). The difference might be affected by the degree of dominance of the species assessed by acoustics in different seasons. The horizontal distribution of fishery resources was dramatically asymmetrical due to the seasonal schooling behavior of fish species like S.ruconius et.al. Furthermore, variation of TS (target strength) frequency and vertical distribution of single echo detection were obvious in different seasons with the depth of 15 m as the boundary.
-
Keywords:
- biomass density /
- fishery resources /
- acoustic assessment /
- bottom trawl /
- Nan'ao Island
-
中国内陆咸水资源丰富,以盐碱湖泊、地下咸水以及盐碱化沼泽的形式广泛分布在内陆的19个省级行政区,绝大多数处于闲置状态。盐碱水渔业以渔业开发为基础,对盐碱水资源进行有效利用,并在适宜盐碱水养殖的品种筛选及规模化养殖方面初具规模[1-2]。凡纳滨对虾 (Litopenaeus vannamei) 又称南美白对虾,可适应盐度范围为0.5~50,因抗逆性较强、盐度耐受性广等特点,在我国内陆地区有很大的养殖规模及产量,同时也是盐碱水养殖主要的驯养对象之一[3-5]。在凡纳滨对虾的内陆养殖过程中,人们往往采用地下咸水调节盐度或直接利用盐碱水资源开展养殖。盐碱水与海水在化学特性上存在明显差别,其中钾离子 (K+) 浓度较低、离子比例失衡等已被认为是其常见的特性之一,使得利用天然咸水进行对虾养殖的效果不理想[6-7]。即使在初始K+浓度正常的水体中,自然降水、池塘土壤对K+的吸附以及非交换性固定也易造成盐度和K+浓度降低[8-10],因此,在对虾内陆养殖过程中,K+缺乏作为一个常见现象被广泛报道[11]。
因考虑到盐度的不同,学者们多用钠离子与钾离子的浓度比值 (Na+/K+) 作指标,针对凡纳滨对虾在盐碱水条件下的生理代谢、免疫酶活、生长存活等方面开展了一系列实验,以确定凡纳滨对虾生长的最适Na+/K+条件,而K+缺乏对凡纳滨对虾的毒性效应则鲜有报道[12-14]。此外,前期相关研究多在单一盐度环境下进行,多盐度条件下的报道仅见于Pathak等[15]与刘存歧等[16]利用天然盐碱水在盐度15、10、5条件下的凡纳滨对虾生长实验,以及Zhu等[17]利用人工盐碱水在盐度30和15条件下的生长代谢研究。本研究以内陆盐碱水养殖盐度较低、Na+/K+变化大、易波动的现象为背景,通过实验生态学方法,在温度为 (31±0.2) ℃、溶解氧质量浓度为 (5.8±0.1) mg·L−1条件下,对体质量为 (1.35±0.37) g的凡纳滨对虾在盐度2、4、8、12、16梯度下开展了为期72 h的低钾胁迫实验,研究不同盐度下低钾胁迫凡纳滨对虾的半致死时间 (LT50) 以及存活情况,同时结合肌肉和肝胰腺的组织结构变化,分析凡纳滨对虾低钾应激的生理响应过程,为其内陆盐碱水养殖以及环境调控提供依据。
1. 材料与方法
1.1 材料
1.1.1 凡纳滨对虾
实验在中国水产科学研究院南海水产研究所珠海试验基地进行。所用凡纳滨对虾取自试验基地盐度为4的养殖池塘。选取凡纳滨对虾幼虾1 600尾 (平均体质量1.0 g、平均体长4.4 cm) 分别放置于25只体积为450 L的玻璃钢圆桶中暂养,每5桶一组,共5组。暂养期间少量投饵并间利用曝气的自来水以及海水盐每天调节2的盐度,将各组盐度分别调整至2、4、8、12和16。在暂养第8天,即盐度为16组升到相应盐度48 h以上后作为实验用虾,此时幼虾平均体质量为 (1.35±0.37) g、平均体长为 (4.95±0.49) cm。
1.1.2 主要试剂与设备
试剂:分析纯KCl (生工生物)、无钾海水素 (江西海鼎科技有限公司)、海水晶 (广州海神水产科技有限公司)、波恩氏 (Bouin's) 液。设备:ProPlus YSI水质分析仪 (美国维赛)、10 L透明带盖方形塑料桶、E6315-210 g-0.1 mg分析天平 (芯硅谷)、生物光学显微镜 (尼康) 等。
1.2 实验方法
1.2.1 低钾胁迫实验
实验设置5个盐度水平:2、4、8、12、16,设置6个Na+/K+水平:27、50、75、100、125、150,盐度和Na+/K+共30个组合,每个组合分别设置3个重复。实验在充气的10 L透明塑料桶中进行,每桶盛放8 L水,放置10尾对虾。利用人工无钾海水素、充分曝气的自来水 (Na+: 10.9 mg·L−1, K+: 2.49 mg·L−1) 和分析纯KCl配置实验用水,其中Na+/K+ 27为对照组,K+浓度正常,其余为低钾实验组 (表1)。实验期间各组溶解氧控制在 (5.8±0.1) mg·L−1,每3 h观察并记录实验虾的死亡情况,并及时将死亡对虾捞出。为减少因饥饿造成的对虾互相残食,每24 h少量投饵1次并及时清理粪便,实验进行到第36小时半量换水1次。
表 1 实验用人工海水的盐度、钠离子与钾离子质量浓度和钠离子/钾离子Table 1 Salinity, concentration of Na+, K+ and Na+/K+ ratio of experimental artificial seawatermg·L−1 盐度
Salinity钠离子/钾离子 Na+/K+ 27 50 75 100 125 150 2 624.97∶23.22 643.62∶12.85 646.21∶8.62 647.51∶6.48 648.29∶5.2 648.83∶4.32 4 1 241.51∶45.78 1 277.5∶25.55 1 282.65∶17.1 1 285.24∶12.87 1 286.8∶10.29 1 287.84∶8.59 8 2 482.05∶91.94 2 552.95∶51.04 2 563.23∶34.17 2 568.4∶25.69 2 571.51∶20.58 2 573.6∶17.16 12 3 732.64∶138.46 3 838.72∶76.77 3 854.18∶51.38 3 861.95∶38.63 3 866.64∶30.93 3 869.77∶25.8 16 4 993.39∶184.15 5 134.94∶102.72 5 155.63∶68.67 5 166.02∶51.67 5 172.3∶41.37 5 176.47∶34.52 注:表中比号前后数值分别为钠离子、钾离子质量浓度。 Note: Data before and after the sign of ratio are concertations of Na+ and K+, respectively. 1.2.2 组织样品的采集与处理
根据低钾胁迫实验结果,取盐度为4、8、12、16,Na+/K+为27、100、125、150共16个组合,在相同条件下对凡纳滨对虾幼虾进行胁迫实验。在实验进行到第48小时取各组还存活的对虾2尾,取背部肌肉以及完整的肝胰腺组织放入波恩氏液中固定,24 h后用70%乙醇清洗多余的波恩氏液,之后保存于70%乙醇中用于石蜡切片的制作和观察。参照魏杰等[18]的方法,组织经乙醇脱水、二甲苯透明、石蜡包片、切片、HE染色、树胶封片、干燥等步骤后,用生物光学显微镜观察并拍照。
1.3 数据计算与分析
实验数据使用SPSS 19.0和Excel 2016软件进行分析,参照Ramirez等[19]与Honkanen等[20]的方法,使用SPSS 19.0软件中的probit回归模型计算低钾胁迫下凡纳滨对虾的LT50,通过Kaplan-Meier生存分析模块绘制生存函数。对成活率作双因素方差分析,如果差异显著 (P<0.05),则进一步对同一盐度或Na+/K+下的各处理组进行Duncan's多重范围检验。不满足方差齐性时,对数据进行自然对数或平方根转换。通过Excel 2016软件进行盐度与对虾第72小时成活率回归分析。
2. 结果
2.1 不同盐度和Na+/K+下凡纳滨对虾的累计生存情况及LT50
Na+/K+为27的对照组以及Na+/K+为50的实验组个体表现正常,截止到第72小时实验结束,各组成活率均高于75%。Na+/K+为75以上各实验组凡纳滨对虾的死亡情况总体表现为随水体盐度的上升而增加。Na+/K+越高,不同盐度下对虾的死亡率差异随胁迫时间延长越明显。其中Na+/K+为150时,盐度16组的受试对虾在60 h内全部死亡,而盐度2、4、8、12组在第60小时的成活率则分别为90%、73.33%、63.33%、46.66%。Na+/K+为75的条件下,盐度2、4、8、12、16组在第60小时的成活率分别为86.66%、80%、83.33%、86.66%、80%;第72小时的成活率分别为83.33%、76.66%、80%、73.33%、66.66% (图1)。
![]() 图 1 不同盐度和钠离子/钾离子下凡纳滨对虾幼虾的成活率Fig. 1 Survival rate of L. vannamei in artificial seawater at different Na+/K+ ratios and salinities
图 1 不同盐度和钠离子/钾离子下凡纳滨对虾幼虾的成活率Fig. 1 Survival rate of L. vannamei in artificial seawater at different Na+/K+ ratios and salinitiesProbit回归模型预测结果显示,凡纳滨对虾的LT50随着Na+/K+和盐度的升高而降低 (表2)。实验观察结果显示,在Na+/K+为100时,盐度12和16条件下凡纳滨对虾的LT50均为63 h;Na+/K+为125时,盐度12和16条件下凡纳滨对虾的LT50分别为60和45 h;Na+/K+为150时,盐度12和16条件下凡纳滨对虾的LT50分别为60和39 h。在Na+/K+为100、125、150条件下凡纳滨对虾第72小时的成活率与盐度的回归方程分别为lny=0.120 8x+10.099 (R2=0.877 5)、lny=0.127x+11.705 (R2=0.982 2)、lny=0.119 5x+16.235 (R2=0.979 7)。
表 2 不同钠离子/钾离子下凡纳滨对虾幼虾半致死时间Table 2 Regression equation between half lethal time and salinity of juvenile L. vannamei钠离子/钾离子
Na+/K+盐度
Salinity半致死时间
Half lethal
time/h实测半致死
时间
Actual half
lethal time/h75 4 — — 8 — — 12 — — 16 81.83 — 100 4 — — 8 — — 12 69.78 63 16 60.15 63 125 4 — — 8 76.23 — 12 62.61 60 16 49.10 45 150 4 87.24 — 8 68.65 — 12 59.40 60 16 39.95 39 2.2 不同盐度和Na+/K+对凡纳滨对虾幼虾72 h成活率的影响
不同盐度与Na+/K+下凡纳滨对虾的成活率见表3。在盐度2条件下,不同Na+/K+组成活率均高于80%且彼此差异不显著 (P>0.05)。在盐度4条件下,Na+/K+为27的正常对照组成活率最高 (93.33%),与Na+/K+为150成活率最低组差异显著 (P<0.05),与其他组差异不显著 (P>0.05)。在盐度8条件下,Na+/K+为150实验组成活率最低并显著低于除Na+/K+为125外的其他各组 (P<0.05),正常组的成活率最高且显著高于Na+/K+为125的实验组 (P<0.05)。在盐度12和16条件下,凡纳滨对虾幼虾成活率随着Na+/K+升高出现明显降低的趋势,Na+/K+为27、50和75组的成活率显著高于其他各组。双因子方差分析结果显示,盐度、Na+/K+及交互对凡纳滨对虾的成活率影响极显著 (P<0.001)。在Na+/K+为27、50、75条件下,盐度2~16各组之间成活率无显著差异 (P>0.05)。当Na+/K+大于100时,整体上凡纳滨对虾的成活率呈现随盐度升高而下降的趋势,Na+/K+为100时,盐度2~8组的成活率高于盐度12和16组且差异显著 (P<0.05);Na+/K+为125时,盐度2和4组的成活率最高且差异不显著 (P>0.05),其他各组之间差异显著 (P<0.05);Na+/K+为150时,盐度4组与盐度2、8组成活率差异不显著 (P>0.05),其他各组之间差异显著 (P<0.05,表3)。
表 3 不同盐度、钠离子/钾离子下凡纳滨对虾成活率Table 3 Survival rate of L. vannamei at different salinities and Na+/K+% 钠离子/钾离子
Na+/K+盐度 Salinity 2 4 8 12 16 27 90±0 93.33±5.77a 83.33±5.77a 86.67±5.77a 80±0a 50 86.67±5.77 76.67±15.28ab 76.67±5.77ab 83.33±11.55a 76.67±15.28a 75 83.33±11.55 76.7±15.28ab 80±10ab 73.33±5.77a 66.67±5.78a 100 83.33±5.77A 86.67±5.77abA 80±10abA 43.33±15.28bB 33.33±5.78bB 125 83.33±5.77A 83.33±11.55abA 66.67±5.77bcB 43.33±5.78bC 13.33±5.78cD 150 80±10A 70±10bAB 60±10cB 20±10cC 0 双因子方差分析结果 Two-way analysis of variance 盐度 Salinity S (<0.001) 钠离子/钾离子 Na+/K+ S (<0.001) 交互 Salinity×Na+/K+ S (<0.001) 注:同列不同小写字母上标表示差异显著 (P<0.05);同行不同大写字母上标表示差异显著(P<0.05)。 Note: Values with different lowercase superscripts within the same column indicate significant difference (P<0.05); values with different uppercase superscripts within the same row indicate significant difference (P<0.05). 2.3 盐度和Na+/K+对凡纳滨对虾肝胰腺及肌肉组织结构的影响
对虾的肝胰腺由发达的多级分支的小管组成,肝胰腺小管间具有脂肪组织与结缔组织,并伴有血管分布。肝胰腺小管外壁由单层柱状上皮细胞构成,管腔的横切面多呈多角星或卵圆形 (图2-a)。盐度4、Na+/K+为150的实验组对虾肝胰腺小管壁组织完整,小管排列比较整齐且边界清晰,与对照组无明显差异 (图2-b)。盐度8、Na+/K+为150的实验组对虾肝胰腺小管排列比较整齐,但可见部分肝胰腺的小管壁出现破损 (图2-c)。盐度12、Na+/K+为125的条件下,对虾肝胰腺小管管腔缩小,同时小管形状与排列不规则 (图2-e)。盐度12、Na+/K+为150,盐度16、Na+/K+为125的实验组对虾肝胰腺小管排列混乱,部分肝胰腺小管间的边界模糊,管腔几乎不可见,可见细胞核和组织残片散落在肝小管间隙 (图2-d、2-f)。
![]() 图 2 不同盐度和钠离子/钾离子对凡纳滨对虾肝胰腺显微结构的影响 (HE 染色,200×)a. 盐度4,钠离子/钾离子 27,示基膜 (BM) 和管腔 (L);b. 盐度4,钠离子/钾离子 150;c. 盐度8,钠离子/钾离子 150,示肝胰腺小管基膜破损;d. 盐度12,钠离子/钾离子 150,示肝胰腺小管管腔变形;e. 盐度12,钠离子/钾离子 125;f. 盐度16,钠离子/钾离子 125,示肝胰腺小管界限模糊。Fig. 2 Effects of salinity and Na+/K+ on hepatopancreas microstructure of L. vannamei (HE staining, 200×)a. Salinity 4, Na+/K+ 27, showing basement membranes (BM) and lumen (L); b. Salinity 4, Na+/K+ 150; c. Salinity 8, Na+/K+ 150, showing basement membranes of hepatopancreas tubule were ruptured; d. Salinity 12, Na+/K+ 150, showing lumen variant; e. Salinity 12, Na+/K+ 125; f. Salinity 16, Na+/K+ 125, showing the demarcation line of hepatopancreas tubule was blurred.
图 2 不同盐度和钠离子/钾离子对凡纳滨对虾肝胰腺显微结构的影响 (HE 染色,200×)a. 盐度4,钠离子/钾离子 27,示基膜 (BM) 和管腔 (L);b. 盐度4,钠离子/钾离子 150;c. 盐度8,钠离子/钾离子 150,示肝胰腺小管基膜破损;d. 盐度12,钠离子/钾离子 150,示肝胰腺小管管腔变形;e. 盐度12,钠离子/钾离子 125;f. 盐度16,钠离子/钾离子 125,示肝胰腺小管界限模糊。Fig. 2 Effects of salinity and Na+/K+ on hepatopancreas microstructure of L. vannamei (HE staining, 200×)a. Salinity 4, Na+/K+ 27, showing basement membranes (BM) and lumen (L); b. Salinity 4, Na+/K+ 150; c. Salinity 8, Na+/K+ 150, showing basement membranes of hepatopancreas tubule were ruptured; d. Salinity 12, Na+/K+ 150, showing lumen variant; e. Salinity 12, Na+/K+ 125; f. Salinity 16, Na+/K+ 125, showing the demarcation line of hepatopancreas tubule was blurred.实验开始12 h后,盐度12与16,Na+/K+为100~150的各组对虾活动迟缓,停止摄食。死亡率随着胁迫时间的延长而上升,同时肉眼可见对虾背部肌肉发白。不同盐度和Na+/K+下凡纳滨对虾幼虾肌肉组织显微结构见图3。盐度4各实验组对虾肌肉纤维排列整齐、紧密,肌肉束间隙较小 (图3-a、3-b);盐度8、Na+/K+为150的实验组对虾肌纤维排列整齐,但肌纤维间隙较对照组有所增大 (图3-c);盐度12、Na+/K+为125的实验组对虾肌纤维间隙宽度增加,可见肌细胞细胞核散落在组织间隙中。随着盐度或Na+/K+的进一步升高,对虾肌纤维发生自溶或者断裂,肌肉组织结构模糊不清,其中盐度16、Na+/K+为125的实验组对虾肌肉组织病变的情况较盐度12、Na+/K+为150的实验组更为严重 (图3-d、3-f)。
![]() 图 3 不同盐度和钠离子/钾离子对凡纳滨对虾肌肉显微结构的影响 (HE 染色,200×)a. 盐度4,钠离子/钾离子 27;b. 盐度4,钠离子/钾离子 150;c. 盐度8,钠离子/钾离子 150;d. 盐度12,钠离子/钾离子 150;e. 盐度12,钠离子/钾离子 125;f. 盐度16,钠离子/钾离子 125。Fig. 3 Effects of salinity and Na+/K+ on muscle microstructure of L. vannamei (HE staining, 200×)a. Salinity 4, Na+/K+ 27; b. Salinity 4, Na+/K+ 150; c. Salinity 8, Na+/K+ 150; d. Salinity 12, Na+/K+ 150; e. Salinity 12, Na+/K+ 125; f. Salinity 16, Na+/K+ 125.
图 3 不同盐度和钠离子/钾离子对凡纳滨对虾肌肉显微结构的影响 (HE 染色,200×)a. 盐度4,钠离子/钾离子 27;b. 盐度4,钠离子/钾离子 150;c. 盐度8,钠离子/钾离子 150;d. 盐度12,钠离子/钾离子 150;e. 盐度12,钠离子/钾离子 125;f. 盐度16,钠离子/钾离子 125。Fig. 3 Effects of salinity and Na+/K+ on muscle microstructure of L. vannamei (HE staining, 200×)a. Salinity 4, Na+/K+ 27; b. Salinity 4, Na+/K+ 150; c. Salinity 8, Na+/K+ 150; d. Salinity 12, Na+/K+ 150; e. Salinity 12, Na+/K+ 125; f. Salinity 16, Na+/K+ 125.3. 讨论
3.1 低钾胁迫对凡纳滨对虾的毒理效应
作为水生动物必需矿物元素之一,钾对于保持体液的渗透压和酸碱平衡、维持动物肌肉和神经的正常效能起着非常重要的作用[21]。Pillard等[22]利用人工配置海水,在盐度31的环境下测试K+浓度对糠虾 (Mysidopsis bahia) 48 h内存活的影响,结果显示不仅高钾,低钾也会引起糠虾死亡。研究表明,在低钾情况下,眼斑拟石首鱼 (Sciaenops ocellatus)、日本鳗鲡 (Anguilla japonica) 的成活率均随Na+/K+增加和胁迫时间的延长而降低[21,23]。本研究中凡纳滨对虾在相同盐度下的死亡率随Na+/K+的增加呈上升趋势,与上述实验结果相似。Prangnell和Fotedar[24]研究了不同K+浓度盐碱水和添加淡水对宽沟对虾 (Penaeus latisulcatus) 生理指标的影响,结果显示Na+/K+为107.9的高Na+/K+组宽沟对虾在第24小时的渗透压调节能力、血钾、血镁显著低于Na+/K+为31.9和25.3的低Na+/K+组,肌肉含水量则显著高于后两组。低Na+/K+组宽沟对虾的血清渗透压、钠浓度和渗透压调节能力在盐度胁迫24 h后停止下降并保持稳定,而高Na+/K+组则在实验期间持续下降,表明严重缺钾不仅会增加对虾的能量消耗,降低血钾,还对宽沟对虾的渗透调节功能造成破坏。Zhu等[12]在盐度30的环境下开展了7个Na+/K+梯度的凡纳滨对虾养殖实验,结果显示高Na+/K+降低了对虾的湿质量、增重率、特定生长率和食物转化率,Na+/K+为110.5组的对虾在2周内全部死亡。本研究中,在盐度为12和16、Na+/K+为125和150的环境下,凡纳滨对虾行动迟缓并停止摄食、LT50明显降低,表现出高Na+/K+对凡纳滨对虾的急性毒性效应,可能是由于高Na+/K+对凡纳滨对虾的渗透压稳态造成严重破坏,引发虾血钾浓度快速降低等急性刺激所造成的,具体原因有待于进一步研究证实。
3.2 盐度对凡纳滨对虾低钾耐受性影响
本实验结果显示,盐度能够对Na+/K+胁迫下的凡纳滨对虾的存活情况产生明显影响,即低盐度下虾对异常Na+/K+的适应能力高于高盐度下,对高Na+/K+的耐受范围更广。中长期对虾养殖实验结果也表明,高Na+/K+条件下,低盐度环境往往会取得更好的养殖效果。Valenzuela-Madrigal等[14]在盐度1.2±0.5的条件下开展了4周的凡纳滨对虾养殖实验,结果显示Na+/K+为162.5的实验塘中对虾成活率达到76.4%,而在盐度4、Na+/K+为120的条件下,4~6周的成活率仅为10%~21.25%[25-26]。Zhu等[15]在盐度30和15的条件下,分析了不同Na+/K+对凡纳滨对虾幼虾生长、物质和能量吸收及食物转化率的影响,结果表明盐度30下,对虾在Na+/K+ (mg/mg) 为20.1~27.9范围内能获得良好生长,在盐度约15的环境下将水体Na+/K+降低到44.8,即可达到较好的养殖效果。
在氨氮 (NH4-N)、亚硝酸盐对凡纳滨对虾和墨吉对虾 (Fenneropenaeus merguiensis) 的毒性试验中,氨氮、亚硝酸盐的毒性随盐度的降低而升高,即低渗环境可以降低对虾对氨氮、亚硝酸盐的耐受性[27-29]。而在低K+环境下,低渗环境提高了凡纳滨对虾对低K+条件的耐受。Partridge和Lymbery[30]研究显示,高渗条件下尖吻鲈 (Lates calcarifer) 对K+的需求更高,在盐度为45的高渗条件下,尖吻鲈在Na+/K+为108的环境下无法存活,在Na+/K+为54的环境下表现为负生长,而在盐度为15和5的低渗环境下的生长与正常环境组无明显差异。Tantylo和Fotedar[31]在盐度分别为5、15、25、35的环境下,对斑节对虾 (P. monodon) 开展Na+/K+为120的胁迫实验,结果显示盐度15、25、35下斑节对虾在第6、第7、第10天全部死亡,仅盐度5下的斑节对虾有较强的血淋巴渗透压和Na+、K+、Ca2+、Mg2+调节能力,实验结束后获得88.9%以上的成活率。在一定的盐度范围内,盐度降低,高Na+/K+的毒性随之降低,这可能与盐度影响对虾的渗透压和钾代谢有关。因此,养殖生产者在考虑水体中K+浓度时,还须将盐度作为一个重要考虑因素。
3.3 低钾胁迫对凡纳滨对虾肌肉和肝胰腺组织结构的影响
人体中细胞内和胞外钾离子浓度梯度对维持细胞静息电位、神经肌肉兴奋性和心肌自律性十分重要,低血钾可导致心律失常、肌无力、骨骼肌痉挛、横纹肌溶解等症状[32]。Knochel等[33]认为,骨骼肌在运动过程中需要释放K+来维持小动脉的扩张,进而保证肌肉的正常供血,缺钾会导致肌肉因供血不足而发生自溶。低血钾兔心肌切片显示心肌细胞肿胀、排列紊乱,胞质内可见大小不等的空泡。陆敏等[34]认为低血钾降低了Na+-K+-ATP酶活性,影响了心肌细胞的代谢并导致其坏死。Tantylo和Fotedar[31]研究指出高Na+/K+会引起斑节对虾血清钾浓度降低,盐度越高,斑节对虾对血钾的维持能力越弱。在缺钾条件下,高渗和等渗环境下尖吻鲈通过骨骼肌中的K+来维持血钾的稳定[30]。对虾的肌肉属于横纹肌,本研究中对虾在高Na+/K+条件下肌肉自溶或断裂的现象与缺钾的症状相似,且随着盐度的升高该现象更加明显,表明高盐度会加剧因高Na+/K+引起的对虾肌肉钾缺乏症状。
本研究发现,高Na+/K+及盐度会造成凡纳滨对虾肝胰腺组织的损伤,在高Na+/K+下,随着盐度升高,对虾首先出现肝胰腺小管壁破损现象,继而出现比较明显的形态结构变化。肝胰腺是对虾能量物质代谢的主要场所,在日本沼虾 (Macrobrachium nipponense) 的低氧胁迫和克氏原螯虾 (Procambarus clarkii) 的低pH胁迫试验中,虾的肝胰腺小管中B细胞转运泡体积首先出现变化,后随着时间延长小管形态出现挤压变形等变化,未见细胞坏死等描述[35-36]。而在塔玛亚历山大藻毒素对中国明对虾 (F. chinensis) 急性毒性实验中,虾肝小管排列较整齐,边界较清晰,但肝小管出现变形,可见坏死的细胞核以及残余组织散落在小管间隙[37]。Pathak等[15]研究显示缺钾型内陆盐碱水养殖的凡纳滨对虾肝胰腺组织出现了管腔变形、血细胞浸润、小管基膜坏死、组织残片和细胞崩解等现象。与pH、溶解氧等环境因子胁迫相比,高Na+/K+主要引起了肝胰腺小管细胞坏死,对小管细胞的分泌功能和分泌活动的影响不明显,组织观察效果与外源毒素对虾类肝胰腺破坏效果更类似。
综上所述,本研究表明Na+/K+和盐度对凡纳滨对虾幼虾存活存在交互作用。盐度能够对Na+/K+胁迫下的凡纳滨对虾存活产生明显影响。在较低盐度下,凡纳滨对虾对高Na+/K+耐受性更强,耐受范围更广。在Na+/K+大于75、盐度大于8的条件下,随着盐度或Na+/K+升高,凡纳滨对虾LT50和72 h成活率明显降低。水中的K+缺乏会对凡纳滨对虾肌肉和肝胰腺组织结构造成损伤,盐度越高毒性效果越明显。因此,在开发利用缺钾型盐碱水进行凡纳滨对虾养殖时,应综合盐度因素考虑Na+/K+对凡纳滨对虾的影响。在低盐条件下,缺钾对凡纳滨对虾短期存活的影响往往并不显著,但在长期养殖过程中,仍需关注钾缺乏对其生长的影响。
-
![]()
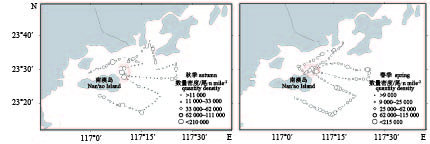
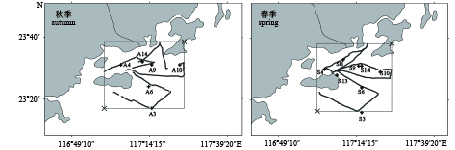
图 1 南澳岛海域声学调查区域与拖网站位分布图
A3~A14. 秋季拖网位点;S3~S14. 春季拖网位点
Figure 1. Survey area of acoustic assessment and distribution of trawl sampling sites in Nan'ao Island area
A3–A14. trawl sampling sites in autumn; S3–S14. trawl sampling sites in spring
![]()
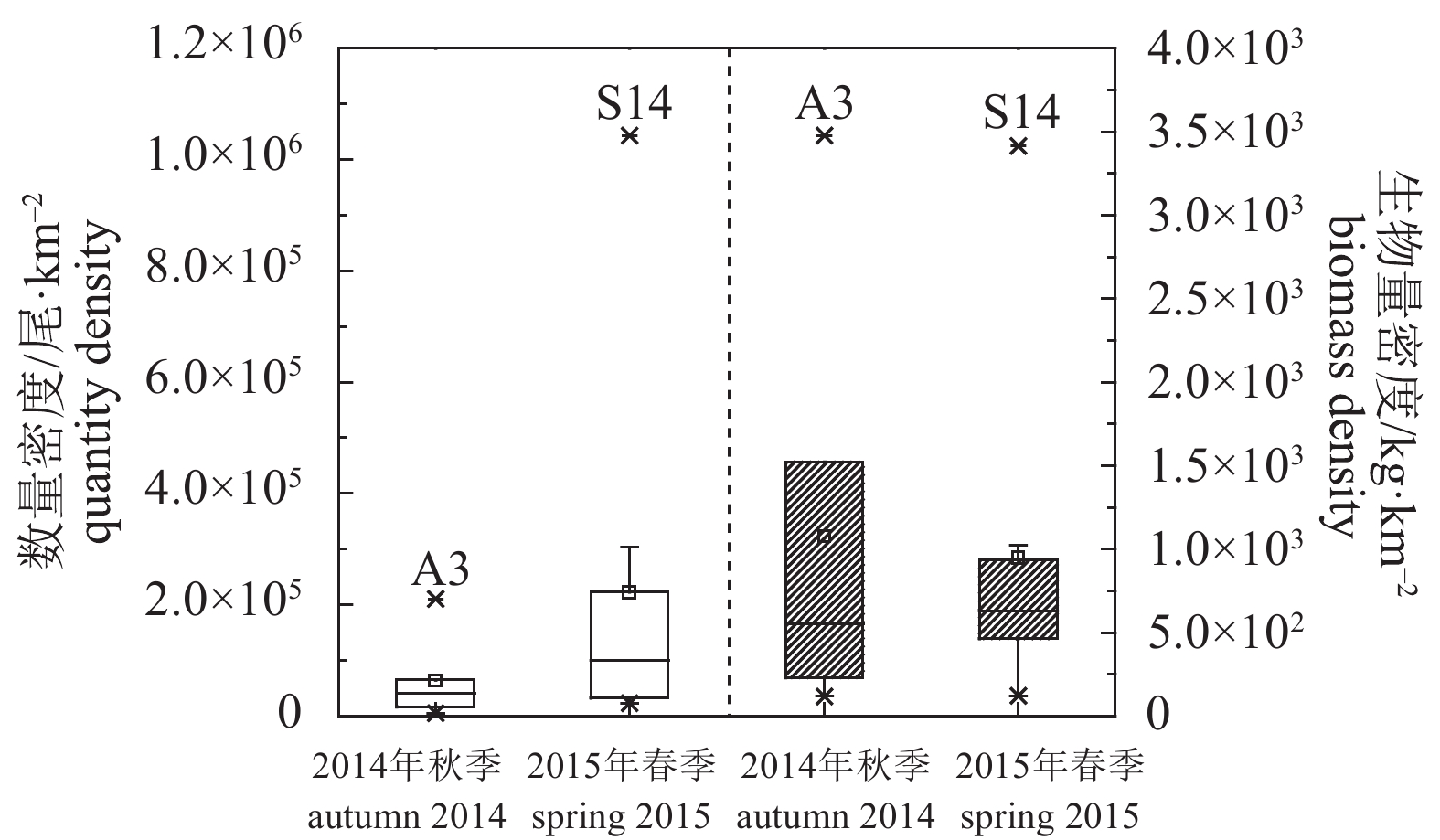
图 3 调查海域各拖网位点参与声学评估种类数量与生物量密度
Figure 3. Quantity and biomass density of acoustic assessed species in each trawling site of survey area
![]()
图 4 种类单一和混合2种不同条件下调查海域渔业资源声学评估
误差线. 地理空间差异;**. α=0.05水平下差异显著
Figure 4. Acoustic assessment of fishery resources in survey area in single or multi-species assumptions
Error bar. geographical spatial difference; **. significant difference at α=0.05 level
![]()
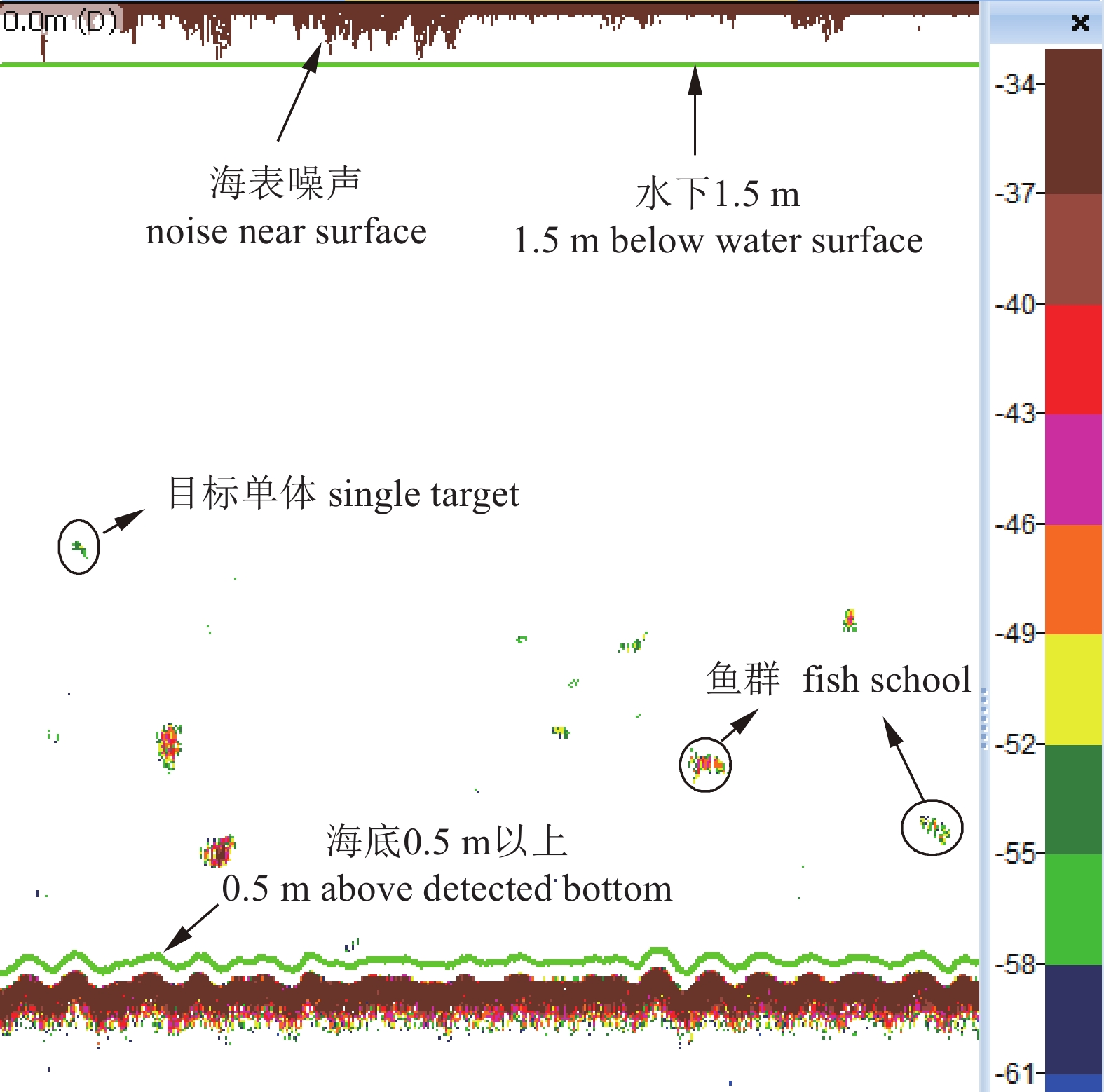
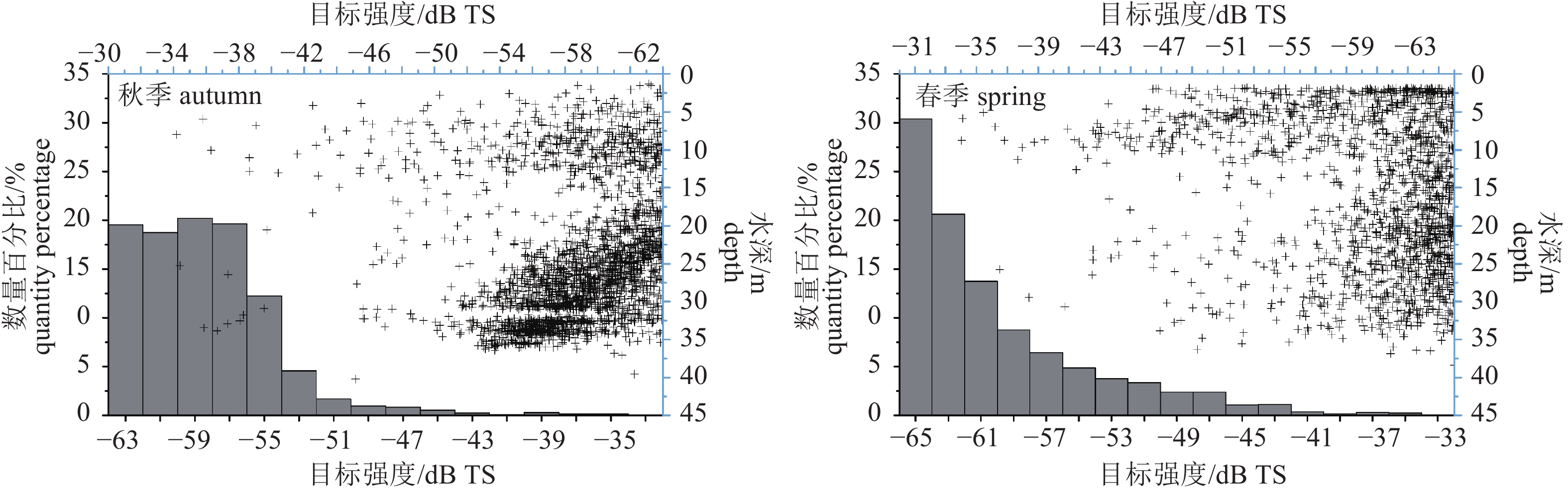
图 6 调查海域回波单体频率组成与垂直分布
Figure 6. Frequency and vertical distribution of single echo detection in survey area
表 1 EY60科学鱼探仪主要技术参数设定
Table 1 Setting of main technical parameters of scientific echosounder EY60
技术参数
technical parameter2014年秋季
autumn 20142015年春季
spring 2015换能器频率/kHz transducer frequency 120 70 发射功率/W transmitting power 200 300 脉冲宽度/μs pulse duration 128 512 等效波束角/dB equivalent beam angle –21 –21 换能器增益/dB transducer gain 26.8 25.69 横向波束宽度/(°) athwardship 3 dB beam width 7 6.53 纵向波束宽度/(°) alongship 3 dB beam width 7 6.43 吸收系数/dB·km–1 absorption coefficient 45.3 18.99 声速/m·s–1 sound speed 1 531.82 1 536.75  下载: 导出CSV
下载: 导出CSV
表 2 主要生物学取样设置
Table 2 Main setting of biological sampling
主要参数
main parameter2014年秋季
autumn 20142015年春季
spring 2015采样类型 sample type 底拖网 底拖网 发动机功率/kW engine power 280 280 平均拖网速度/km·h–1 average trawling speed 5.56 4.93 平均采样时长/h average trawling time 0.55 0.46 网口宽度/m net width 6 4 网口高度/m net height 2 2 网目/cm net size 3×3 3×3
下载: 导出CSV
表 3 声学评估优势种类生物学信息
Table 3 Biological information of dominant species in acoustic assessment
季节(年份)
season (year)物种
species数量百分比/%
quantity percentage生物量百分比/%
biomass percentage相对重要性指数
IRI体长范围/mm
body length range体质量范围/g
body mass range秋季 (2014)
autumn短尾大眼鲷
Priacanthus macracanthus45.3 34.8 6 678 72~88 8~21 丝背细鳞鲀
Stephanolepis cirrhifer12.0 20.8 2 187 68~107 6~55 丽叶鲹
Caranx kalla9.3 6.5 1 322 60~135 4~85 杜氏枪乌贼
Loligo duvaucelii4.9 6.5 1 134 50~123* 6~58 鹿斑鲾
Secutor ruconius7.4 2.3 647 27~70 2~12 二长棘鲷
Parargyrops edita3.5 9.0 624 95~108 32~56 斑鳍白姑鱼
Argyrosomus pawak6.2 2.2 280 54~105 5.3~30 黄斑篮子鱼
Siganus canaliculatus0.6 0.9 129 74~126 5~74 春季 (2015)
spring鹿斑鲾
Secutor ruconius86.9 68.0 11 613 36~63 1.6~10 二长棘鲷
Parargyrops edita5.9 5.4 845 35~62 1.7~58 杜氏枪乌贼
Loligo duvaucelii1.0 6.9 689 42~182* 4.9~140 竹䇲鱼
Trachurus japonicus3.2 4.1 458 60~90 2.4~11 刺鲳
Psenopsis anomala0.5 2.6 229 65~110 8.5~45 短吻鲾
Leiognathus brevirostris1.1 3.6 179 58~94 4.2~24 皮氏叫姑鱼
Johnius belangerii0.3 2.6 145 92~160 14~75 沙带鱼
Lepturacanthus savala0.5 1.8 114 66~150 3.6~45 注:*. 胴长
Note: *. mantle length
下载: 导出CSV
表 4 调查海域声学评估种类丰度与生物量密度
Table 4 Abundance and biomass density of acoustic assessed species in survey area
季节(年份)
season (year)种类
speciesb20/dB 丰度密度/尾·n mile–2
abundance density生物量密度/kg·n mile–2
biomass density秋季 (2014)
autumn短尾大眼鲷 Priacanthus macracanthus –68.0 12 142.92 150.22 丝背细鳞鲀 Stephanolepis cirrhifer –76.0 3 216.67 87.92 丽叶鲹 Caranx kalla –72.5 2 492.92 27.80 鹿斑鲾 Secutor ruconius –72.5 1 983.61 9.67 斑鳍白姑鱼 Argyrosomus pawak –68.0 1 661.95 9.46 杜氏枪乌贼 Loligo duvaucelii –80.0 1 313.47 28.32 二长棘鲷 Parargyrops edita –68.0 938.19 39.55 黄斑篮子鱼 Siganus canaliculatus –72.9 160.83 3.50 春季 (2015)
spring鹿斑鲾 Secutor ruconius –72.5 27 791.47 87.42 二长棘鲷 Parargyrops edita –68.0 1 886.88 6.94 竹䇲鱼 Trachurus japonicus –72.5 1 023.39 5.37 杜氏枪乌贼 Loligo duvaucelii –80.0 319.81 9.62 短吻鲾 Leiognathus brevirostris –72.5 351.79 4.58 刺鲳 Psenopsis anomala –80.0 159.90 3.53 沙带鱼 Lepturacanthus savala –66.1 159.90 2.55 皮氏叫姑鱼 Johnius belangerii –68.0 95.94 3.66 注:b20值来自文献记载
Note: The b20 values come from documentary records.
下载: 导出CSV
-
[1] 陈丕茂, 袁华荣, 贾晓平, 等. 大亚湾杨梅坑人工鱼礁区渔业资源变动初步研究[J]. 南方水产科学, 2013, 9(5): 100-108. [2] 张俊, 陈丕茂, 房立晨, 等. 南海柘林湾-南澳岛海洋牧场渔业资源本底声学评估[J]. 水产学报, 2015, 39(8): 1187-1198. [3] TESSIER A, PASTOR J, FRANCOUR P, et al. Video transects as a complement to underwater visual census to study reserve effect on fish assemblages[J]. Aquat Biol, 2013, 18(3): 229-241.
[4] DEMARTINI E E, ROBERTS D. An empirical test of biases in the rapid visual technique for species-time censuses of reef fish assemblages[J]. Mar Biol, 1982, 70(2): 129-134.
[5] SIMMONDS J, MACLENAN D. Fisheries acoustics: theory and practice[M]. 2nd ed. Oxford: Blackwell Science, 2005: 1-379.
[6] KOSLOW J A. The role of acoustics in ecosystem-based fishery management[J]. ICES J Mar Sci, 2009, 66(6): 966-973.
[7] ACOLAS M L, ANRAS M B, VERON V, et al. An assessment of the upstream migration and reproductive behaviour of allis shad (Alosa alosa L.) using acoustic tracking[J]. ICES J Mar Sci, 2004, 61(8): 1291-1304.
[8] KLOSER R J, RYAN T, SAKOV P, et al. Species identification in deep water using multiple acoustic frequencies[J]. Can J Fish Aquat Sci, 2011, 59(6): 1065-1077.
[9] 赵宪勇, 金显仕, 唐启升. 渔业声学及其相关技术的应用现状和发展前景[M]. 北京: 海洋出版社, 2000: 55-62. [10] 陈国宝, 李永振, 赵宪勇, 等. 南海北部海域重要经济鱼类资源声学评估[J]. 中国水产科学, 2005, 12(4): 445-451. [11] 李娜娜, 陈国宝, 于杰, 等. 大亚湾杨梅坑人工鱼礁水域生物资源量声学评估[J]. 水产学报, 2011, 35(11): 1640-1649. [12] 张俊, 陈国宝, 陈作志, 等. 南沙南部陆架海域渔业资源声学评估[J]. 南方水产科学, 2015, 11(5): 1-10. [13] ZENG L, ZHOU L, FU D H, et al. Diel distribution, migration and abundance assessment of Toxabramis houdermeri in Baise Reservoir, China[J]. Folia Zoologica, 2016, 65(1): 1-9.
[14] 赵宪勇, 陈毓桢, 李显森, 等. 多种类海洋渔业资源声学评估技术和方法探讨[J]. 海洋学报, 2003, 25(增刊1): 192-202. [15] SIMRAD. Simrad ER60 scientific echo sounder software reference manual[Z]. Norway: Simrad Maritime as Kongsberg, 2008: 19-31.
[16] AGLEN A. Random errors of acoustic fish abundance estimates in relation to the survey grid density applied[R]. Rome: FAO Fisheries Report, 1983: 293-298.
[17] HIGGINBOTTOM I, WOON S, SCHNEIDER P. Hydroacoustic data processing for standard stock assessment using Echoview: technical manual[Z]. Hobart, Australia: Myriax Software Pty Ltd Publication, 2008: 70-85.
[18] PARKER STETTER S L, RUDSTAM L G, SULLIVAN P J, et al. Standard operating procedures for fisheries acoustic surveys in the Great Lakes[M]. Ann Arbor: Great Lakes Fish Comm Spec Pub, 2009: 103-146.
[19] 袁梦, 汤勇, 徐姗楠, 等. 珠江口南沙海域秋季渔业资源群落结构特征[J]. 南方水产科学, 2017, 13(2): 18-25. [20] 连玉喜, 黄耿, MALGORZATA, 等. 基于水声学探测的香溪河鱼类资源时空分布特征评估[J]. 水生生物学报, 2015, 39(5): 920-929. [21] ELTON C S. Animal ecology[M]. New York: The MacMillan Company, 1930: 187-188.
[22] DICKIE L M, KERR S R, BOUDREAU P R. Size-dependent processes underlying regularities in ecosystem structure[J]. Ecol Monogr, 1987, 57(3): 233-250.
[23] KERR S R, DICKIE L M. The Biomass Spectrum: a predator-prey theory of aquatic production[M]. New York, USA: Columbia University Press, 2001: 83-84
[24] JENNINGS S, WARR K J, MACKINSON S J. Use of size-based production and stable isotope analyses to predict trophic transfer efficiencies and predator-prey body mass ratios in food webs[J]. Mar Ecol Prog Ser, 2002, 240: 11-20.
[25] KUBECKA J, DUNCAN A. Diurnal changes of fish behaviour in a lowland river monitored by a dual-beam echosounder[J]. Fish Res, 1998, 35(1/2): 55-63.
[26] COMTE L, BUISSON L, DAUFRESNE M, et al. Climate-induced changes in the distribution of freshwater fish: observed and predicted trends[J]. Freshw Biol, 2013, 58(4): 625-639.
[27] BOHL E. Diel pattern of pelagic distribution and feeding in planktivorous fish[J]. Oecologia, 1979, 44(3): 368-375.
[28] FILIPA FILIPE A, MARKOVIC D, PLETTERBAUER F A, et al. Forecasting fish distribution along stream networks: brown trout (Salmo trutta) in Europe[J]. Divers Distrib, 2013, 19(8): 1059-1071.
[29] SCALABRIN C, MARFIA C, BOUCHER J. How much fish is hidden in the surface and bottom acoustic blind zones?[J]. ICES J Mar Sci, 2009, 66(6): 1355-1363.
[30] MELLO L S, ROSE G A. The acoustic dead zone: theoretical vs. empirical estimates, and its effect on density measurements of semi-demersal fish[J]. ICES J Mar Sci, 2009, 66(6): 1364-1369.
[31] DRASTIK V, KUBECKA J. Fish avoidance of acoustic survey boat in shallow waters[J]. Fish Res, 2005, 72(2/3): 219-228.
[32] WHEELAND L J, ROSE G A. Quantifying fish avoidance of small acoustic survey vessels in boreal lakes and reservoirs[J]. Ecol Freshw Fish, 2015, 24(1): 67-76.
[33] TORESEN R. Absorption of acoustic energy in dense herring schools studied by the attenuation in the bottom echo signal[J]. Fish Res, 1991, 10: 317-327.
[34] MOWBRAY F K. Changes in the vertical distribution of capelin (Mallotus villosus) off Newfoundland[J]. ICES J Mar Sci, 2002, 59(59): 942-949.
[35] PARAMO J R. Acoustic-geostatistical assessment and habitat–quantity relations of small pelagic fish from the Colombian Caribbean[J]. Fish Res, 2003, 60(2/3): 309-319.
[36] 陈勇, 于长青, 张国胜, 等. 人工鱼礁的环境功能和集鱼效果[J]. 大连水产学院学报, 2002, 17(1): 64-69. [37] NEILSON J D, CLARK D, MELVIN G D, et al. The diel-vertical distribution and characteristics of pre-spawning aggregations of pollock (Pollachius virens) as inferred from hydroacoustic observations: the implications for survey design[J]. ICES J Mar Sci, 2003, 60(4): 860-871.
[38] ERZINI K, GONCALVES J M, BENTES L, et al. Size selectivity of trammel nets in southern European small-scale fisheries[J]. Fish Res, 2006, 79(1/2): 183-201.
[39] RIHA M, JUZA T, PRCHALOVA M, et al. The size selectivity of the main body of a sampling pelagic pair trawl in freshwater reservoirs during the night[J]. Fish Res, 2012, 127(9): 56-60.
[40] GUILLARD J, ALBARET J J, SIMIER M, et al. Spatio-temporal variability of fish assemblages in the Gambia Estuary (West Africa) observed by two vertical hydroacoustic methods: moored and mobile sampling[J]. Aquat Living Resour, 2004, 17(1): 47-55.
[41] ZENG L, ZHOU L, GUO D L, et al. Ecological effects of dams, alien fish, and physiochemical environmental factors on homogeneity/heterogeneity of fish community in four tributaries of the Pearl River in China[J]. Ecol Evol, 2017, 7(11): 3904-3915.
-
期刊类型引用(3)
1. 李彬涛,谭鹤群,张义仁,夏成醒. 进水结构对圆形循环水养殖池流场的影响. 华中农业大学学报. 2024(02): 22-29 .  百度学术
百度学术
2. 李瑞鹏,田云臣,李青飞,丛雪琪,秦海晶. 进水流速对圆形循环水养殖池流场特性影响的数值模拟. 渔业科学进展. 2024(03): 55-65 . 百度学术
3. 史宪莹,李猛,任效忠,冯德军,刘航飞,周寅鑫,刘海波,赵晨旭. 长宽比对双进水管结构矩形圆弧角养殖池排污特性的影响. 大连海洋大学学报. 2023(04): 707-716 . 百度学术
其他类型引用(2)






计量
- 文章访问数: 3686
- HTML全文浏览量: 1243
- PDF下载量: 374
- 被引次数: 5
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
