
Establishment and application of a SYBR Green I real-time qPCR detection of Ameson portunus
-
摘要:
梭子蟹肌孢虫(Ameson portunus)是感染三疣梭子蟹(Portunus trituberculatus)的常见病原之一,该研究以其SSU rRNA基因为靶标建立了SYBR Green I实时定量PCR检测方法。结果显示:该方法扩增效率为101.45%,检测灵敏度下限可达2.0×101 拷贝·μL-1,特异性、组内重复性和组间重复性均好。利用该荧光定量PCR方法对一批来自“牙膏病”流行区的养殖三疣梭子蟹样品进行检测,梭子蟹肌孢虫检出率为82.35%。检测结果显示,活蟹的每毫克肌肉平均带虫量大幅低于死蟹,表明该地区梭子蟹的死亡与肌孢虫的感染密切相关,而且,当梭子蟹肌孢虫载量达到1.0×109 拷贝·mg-1时三疣梭子蟹存在暴发疾病甚至死亡的风险。另外,通过对比检测发现,梭子蟹肌孢虫套式PCR方法对这批样品的检出率为64.71%,表明该研究建立的荧光定量PCR方法具有更高的灵敏度。
-
关键词:
- 梭子蟹肌孢虫 /
- 三疣梭子蟹 /
- 实时定量PCR方法 /
- SYBR Green I
Abstract:Ameson portunus is one of the common pathogens of the swimming crab (Portunus trituberculatus). In this study, a SYBR Green I real time qPCR method whose amplification efficiency reached up to 101.45% with a detection limit as low as 2.0×101 copies·μL-1 was established by targeting A.portunus SSU rRNA gene. The method was A.portunus-specific with good repeatability within and between groups. Then the qPCR method was used to detect farmed swimming crabs from a "toothpaste disease" epidemic area, and the detection rate of A.portunus was 82.35%. The results show that the average muscle load per milligram of the live crabs was lower than that of the dead crabs significantly, which indicates that the death of crabs in the region is related with infection of A.portunus closely. When the load of A.portunus was 1.0×109 copies·mg-1 in the muscle tissue, P.trituberculatus had a risk of illness or even death. Besides, the detection rate for the same samples by a previously established nested-PCR method was only 64.71%, showing that the qPCR method in the present study had higher sensitivity and can provide good technical support for the early diagnosis and prevention of "toothpaste disease".
-
Keywords:
- Ameson portunus /
- Portunus trituberculatus /
- real time qPCR method /
- SYBR Green I
-
磷是微藻生长发育的必需元素,主要存在于单细胞藻的原生质和细胞核中,它作为底物或调节物直接参与光合作用的各个环节,包括光能吸收、同化力的形成、卡尔文循环、同化产物的运输以及对一些关键性酶的活性起调节作用等[1]。它参与植物生长发育过程中的各种代谢活动,在细胞膜结构、物质代谢以及信号传导等方面都起着极为重要的作用[2]。
当微藻受到环境胁迫时,光合作用受到抑制,光合效率降低,藻细胞吸收的过剩光能通过热量以及荧光的形式散发出去。因此,叶绿素荧光的变化可以在一定程度上反映环境因子对微藻的影响[3],通过对不同营养盐浓度下叶绿素荧光参数变化的分析,可深入了解磷浓度对微藻光合机构主要是PSII的影响。叶绿素荧光法具有快速、准确、对细胞无损伤、需要样品量少的优点,是检测微藻营养盐限制最有发展前景的方法[4-5]。
小球藻(Chlorella sp.)属于绿藻门,其蛋白质含量高达55%~65%,氨基酸组成均衡,含有大量的叶绿素,特别是含有生物活性物质糖蛋白、多糖体及高达13%的核酸等物质。小球藻中富含小球藻生长因子(Chlorella growth factor,CGF),能迅速恢复机体造成的损伤,是公认的健康食品。它不仅是轮虫(Brachionus plicatilis)[6]等水产动物的天然饵料,同时很多研究表明,在养殖水体接种小球藻可调节和优化浮游生物的群落结构,增加溶解氧,从而改善水体的化学环境条件[7-8]。因此,合理培养和利用小球藻在水产动物的健康养殖中具有重要意义。
迄今为止,国外已有一些有关叶绿素荧光法研究营养盐胁迫对微藻光合作用影响的报道[9-14],国内有关营养盐浓度对小球藻叶绿素荧光的影响尚未见报道。目前,通过叶绿素荧光法检测营养盐限制情况的主要方法有3种:(1)通过Fv/Fm(最大光能转化效率)比值的变化来检测。(2)通过营养添加后各荧光参数的瞬时变化(nutrient induced fluorescnce transients,NIFT反应)来检测。(3)通过在线叶绿素荧光的变化检测。此文结合前2种方法研究了不同磷浓度对小球藻各荧光参数的影响,并对其逆境反应的机理进行了初步研究,为其利用提供科学依据,推进了营养盐检测方法的发展和完善。
1. 材料与方法
1.1 材料
实验所用小球藻藻种取自中国海洋大学微藻种质库,编号为MACC/C95。
1.2 微藻培养
实验在500 mL的三角烧瓶中进行,除磷浓度外,其它营养盐均采用f/2培养基[15]。磷设4个浓度梯度,分别为0(缺磷组)、10(低磷组)、36.3(对照组)和290.4 μM(高磷组)。每个浓度3个平行组,培养温度为20±1℃,盐度为31,光照强度为100 μmol · m-2 · s-1。培养过程中不充气,每日随机调换三角瓶并摇动2~3次。培养时间为7 d,每天定时取样,进行叶绿素荧光参数、细胞密度、叶绿素含量的测定。
1.3 叶绿素荧光参数的测定
用Water-PAM水样叶绿素荧光仪(Walz,Effectnich,Germany)测定叶绿素荧光参数。测量前将微藻样品暗适应15 min,叶绿素荧光参数Fv/Fm、ΦPSII(实际光能转化效率)、ETR(电子传递效率)、qP(光化学淬灭)、qN(非光化学淬灭)和NPQ(非光化学淬灭)可通过叶绿素荧光仪直接读出。
1.4 叶绿素含量的测定
用Water-PAM水样叶绿素荧光仪测定叶绿素含量,原理是由于瞬间荧光产量(F值)与叶绿素含量在一定范围内成正比,通过校正可测出叶绿素含量。为避免校正过程中可能产生的误差,文中的叶绿素含量用相对含量(每天测得的叶绿素含量与接种时叶绿素含量的比值)表示。
1.5 细胞密度的测定
每天定时取样,用血球计数板测定细胞密度。
1.6 NIFT反应
培养基中,磷的添加量为0,其它营养盐采用f/2培养基。实验的第3、第5和第7天定时取3 mL样品于石英杯中,测定光强为150 μmol · m-2 ·s-1,每30 s进行瞬时荧光、ΦPSII、qP、NPQ的测定,样品稳定10 min后,向样品中迅速添加30 μL磷源(PO43--P),再测定10 min。根据ROBERTS[16]的研究,此测定光强以及磷源的添加量足够引起NIFT反应。
1.7 恢复实验
微藻在不同磷浓度条件下培养7 d后,分别向缺磷组、低磷组和对照组中添加磷至f/2培养基的浓度,培养条件同1.2。每2 h取样测定叶绿素荧光参数,测量方法同1.3。
1.8 数据处理
用SigmaPlot 10.0软件作图。用SPSS 11.5软件分别进行单因子方差分析和多重比较。P < 0.05表示差异显著。
2. 结果
2.1 磷浓度对小球藻叶绿素荧光参数的影响
磷浓度对小球藻叶绿素荧光参数的影响见图 1-a~f,随着培养时间的延长,Fv/Fm、ΦPSⅡ、ETR、qP总体上呈下降趋势。缺磷组下降幅度最大,其中Fv/Fm值从接种时的0.643下降到0.35,接种后第2天开始到实验结束qP均处于最低水平。高磷组的Fv/Fm值从0.643下降到0.455,下降的幅度仅次于缺磷组。单因子方差分析以及多重比较的结果表明,一次性培养过程中,磷浓度对小球藻的叶绿素荧光参数均有显著影响(P < 0.05)。接种后第1天起到实验结束,缺磷组的Fv/Fm、ΦPSⅡ、ETR均显著低于对照组(P < 0.05),高磷组的ΦPSⅡ、ETR、qP以及低磷组的ΦPSⅡ、ETR均显著低于对照组(P < 0.05)。接种后第3天到实验结束,高磷组的Fv/Fm均显著低于对照组(P < 0.05)。高磷组的qN、NPQ随着培养时间的延长不断下降,从接种后第2天开始到实验结束始终处于最低水平。
![]() 图 1 磷浓度对小球藻叶绿素荧光参数(a~f)、细胞密度(g)、叶绿素含量(h)的影响Figure 1. Effects of different P concentrations on the chlorophyll fluorescence parameters (a~f), cell density (g)and chlorophyll content (h) of Chlorella sp.
图 1 磷浓度对小球藻叶绿素荧光参数(a~f)、细胞密度(g)、叶绿素含量(h)的影响Figure 1. Effects of different P concentrations on the chlorophyll fluorescence parameters (a~f), cell density (g)and chlorophyll content (h) of Chlorella sp.磷浓度对小球藻细胞密度的影响见图 1-g,随着培养时间的增加,各组细胞密度不断增长,生长速率按大小顺序为对照组>低磷组>高磷组>缺磷组。单因子方差分析及多重比较的结果表明,磷浓度对小球藻的细胞密度有显著影响(P < 0.05)。
磷浓度对叶绿素相对含量的影响见图 1-h,随着培养时间的延长,各组叶绿素相对含量不断增长,增加速率按大小顺序为对照组>高磷组>低磷组>缺磷组。单因子方差分析及多重比较的结果表明,磷浓度对小球藻的叶绿素相对含量有显著影响(P < 0.05)。
2.2 磷添加后各荧光参数的NIFT反应
磷源添加后小球藻各荧光参数的NIFT反应见图 2-a~d,磷添加后实验第3天,荧光的变化不明显,第5和第7天荧光迅速下降随后有一定的恢复但始终低于磷添加前的水平。实验第3天,ΦPSII的变化不显著,第5和第7天ΦPSII迅速上升随后又很快恢复至磷添加前的水平。实验第3和第5天qP的变化不显著,而第7天qP不断大幅度上升。整个实验过程中,NPQ在磷添加后不断上升,实验第7天的上升幅度最大。
![]() 图 2 小球藻磷限制后添加磷源时的瞬时荧光反应(a~d)Figure 2. Transient perturbations to fluorescence parameters from P-starved for 3, 5 and 7 days Chlorella sp. after P additions as PO43-
图 2 小球藻磷限制后添加磷源时的瞬时荧光反应(a~d)Figure 2. Transient perturbations to fluorescence parameters from P-starved for 3, 5 and 7 days Chlorella sp. after P additions as PO43-2.3 磷添加后各荧光参数24 h内的变化情况
磷添加后各荧光参数24 h内的变化情况见图 3-a~f,添加磷后,缺磷组的Fv/Fm、ΦPSⅡ、ETR、qP呈明显上升的趋势,低磷组的ΦPSⅡ、ETR也呈上升的趋势。恢复24 h后,缺磷组的Fv/Fm从0.350上升到0.526,ΦPSⅡ从0.114上升到0.246,ETR从5.7上升到12.4。对照组的Fv/Fm变化不明显,ΦPSⅡ、ETR在磷添加后12 h后呈现下降的趋势,其Fv/Fm、ΦPSⅡ、ETR最终大小分别为0.610、0.233、11.7。
![]() 图 3 小球藻不同磷浓度下培养7 d重新添加磷源后24 h内各荧光参数(a~f)的变化Figure 3. Changes of fluorescence parameters under different P concentrations for 7 days after addition of P in Chlorella sp.for 24 h (a~f)
图 3 小球藻不同磷浓度下培养7 d重新添加磷源后24 h内各荧光参数(a~f)的变化Figure 3. Changes of fluorescence parameters under different P concentrations for 7 days after addition of P in Chlorella sp.for 24 h (a~f)各组的qN、NPQ在磷添加4 h以后呈现上升的趋势,缺磷组的上升速率最快、上升幅度最大,低磷组及对照组的qN、NPQ也有所上升,但是上升幅度不大。缺磷组、低磷组、对照组的最终qN分别为1.048、0.373、0.517,最终NPQ分别为1.230、0.260、0.469。
3. 讨论
叶绿素荧光与光合作用的各个反应过程紧密相关,任何胁迫对光合作用产生的影响都可以通过体内叶绿素荧光诱导动力学变化反映出来。因此,叶绿素荧光参数可作为胁迫条件下微藻抗逆反应的指标,较好地反映微藻的实际光合状况。
胁迫条件下,Fv/Fm、ΦPSⅡ、ETR、qP都明显下降。此实验中,缺磷、低磷、高磷组的Fv/Fm、ΦPSⅡ、ETR、qP都显著低于对照组。培养7 d后缺磷组Fv/Fm降低的幅度最大,从接种时的0.643下降到0.350,高磷组的Fv/Fm值从0.643下降到0.455。LIPPEMEIER等[11]对微小亚历山大藻(Alexandrium minutum)的研究表明,磷限制条件下培养9 d后,Fv/Fm值从0.67下降到0.42,而对照组的则保持在0.67左右,这和此实验的结果相一致。Fv/Fm下降表明,磷限制使小球藻PSⅡ反应中心受损,阻碍了光合电子传递的过程,抑制光合作用的原初反应[17]。ΦPSⅡ的降低,说明磷胁迫阻止藻细胞同化力(NADPH,ATP)的形成,从而影响对碳的固定与同化。qP的下降表明,电子由PSⅡ的氧化侧向PSⅡ反应中心的传递受阻,用于进行光合作用的电子减少,以热或其它形式耗散的光能增加,这与ΦPSⅡ的下降是吻合的。非光化学淬灭处在较低的水平,说明藻细胞的卡尔文循环活跃,能量利用率高。缺磷胁迫下小球藻的qN逐渐增加,表明其卡尔文循环的活性受抑制的程度增大,PSⅡ的潜在热耗散增加,对藻体本身是一种保护作用。高磷组的qN、NPQ不断下降,表明小球藻的正常生理功能受到严重伤害,对热能的耗散能力不断丧失。
磷缺乏会影响到微藻光合作用中的卡尔文循环,从而影响到叶绿素的合成和细胞的分裂,此时微藻对外界胁迫的适应性减小,其生长速率下降,最终导致细胞停止分裂。此实验结果表明,缺磷组培养2 d后,细胞分裂处于停滞状态,叶绿素的合成受到阻碍。低磷组的最终细胞密度以及叶绿素含量也显著低于对照组。LIPPEMEIER等[11]研究表明,磷限制的条件下,微小亚历山大藻培养20 d后的细胞密度仅为15.6×103,而磷充足的条件下,其最终密度为240×103,这和此实验的结果相一致。氮是合成蛋白质的主要物质基础,磷的缺乏同时影响了微藻对氮的吸收利用,从而严重阻碍了叶绿素的合成。高磷胁迫也会影响微藻的生长,此实验表明,高磷浓度下小球藻的细胞密度以及叶绿素的最终含量显著低于对照组。
此实验中,缺磷培养的小球藻在磷添加后各荧光参数会出现相应的NIFT反应,YOUNG和BEARDALL[10]的研究表明,杜氏藻(Dunaliella tertiolecta)在氮限制后添加NO3-,荧光在15 s内迅速下降,随后上升到最大值再下降至稳定水平,NPQ在前20 s内随着荧光的下降而下降,ΦPSⅡ、qP都有小幅度的变化,而添加蒸馏水则无此现象。HOLLAND等[18]的研究也表明颤藻(Oscillatoria sp.)以及浮水小球藻(C.emersonii)在磷限制后添加PO43-,荧光以及ΦPSⅡ都迅速下降至更低的水平并保持稳定。这和此实验的结果相一致,小球藻在添加磷源后,荧光也出现瞬时下降的现象。HOLLAND等的研究还表明,从自然海区取回的样品并无NIFT反应,经过实验室缺磷培养后,才出现NIFT反应。结果表明,微藻在受到营养盐限制的情况下,重新添加的瞬间会出现NIFT反应,这为后人检测水体中营养盐的限制情况提供一定的依据。
实验结果显示,磷添加后24 h内,小球藻缺磷组和低磷组的叶绿素荧光参数值都有一定恢复,而对照组的变化不明显。LIPPEMEIER等[11]的研究表明,微小亚历山大藻在磷限制9 d后,重新添加磷培养3 d后其Fv/Fm从0.42恢复到0.67,而未重新添加磷的组Fv/Fm继续下降。YOUNG和BEARDALL[13]对杜氏藻的研究也表明,限制性营养盐氮重新添加后24 h内,Fv/Fm从0.4恢复到0.7。这是因为营养盐限制的条件下,微藻处于“饥饿”状态,重新添加营养盐后出现“奢侈性吸收”。
过低或过高的磷浓度都会限制微藻的生长和光合作用,影响微藻的基本生理功能,此实验结果表明,小球藻生长和进行光合作用的最适磷浓度为36.3 μM(即对照组的磷浓度)。微藻受到营养盐限制后,叶绿素荧光会发生相应的变化。限制性营养盐重新添加后,荧光参数是否出现NIFT反应、NIFT反应的大小、是否能够恢复以及恢复的程度和微藻的种类以及营养盐限制的程度有着密切的联系。因此,可以根据营养盐限制条件下叶绿素荧光参数的变化情况、限制性营养盐添加瞬间各荧光参数的NIFT反应以及营养盐重新添加后各荧光参数的恢复情况来判断微藻营养盐限制的情况,研究其对营养盐限制条件的适应性。
-
![]()
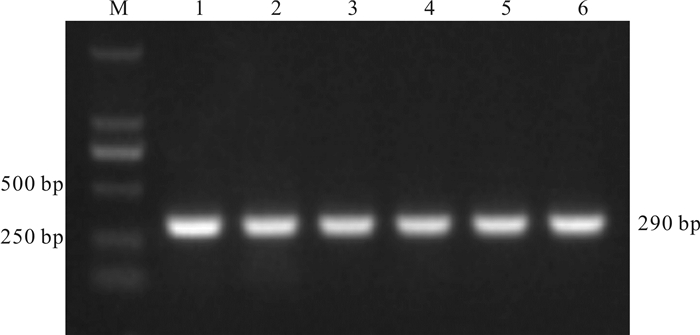
图 1 质粒中目的片段的PCR扩增结果
M. DL2000 marker;1~6.质粒标准品(浓度为2.0×104 拷贝·μL-1)
Figure 1. PCR products of A.portunus gene fragments cloned in the plasmid
1~6.A.portunus standards (2.0×104 copies·μL-1)
![]()
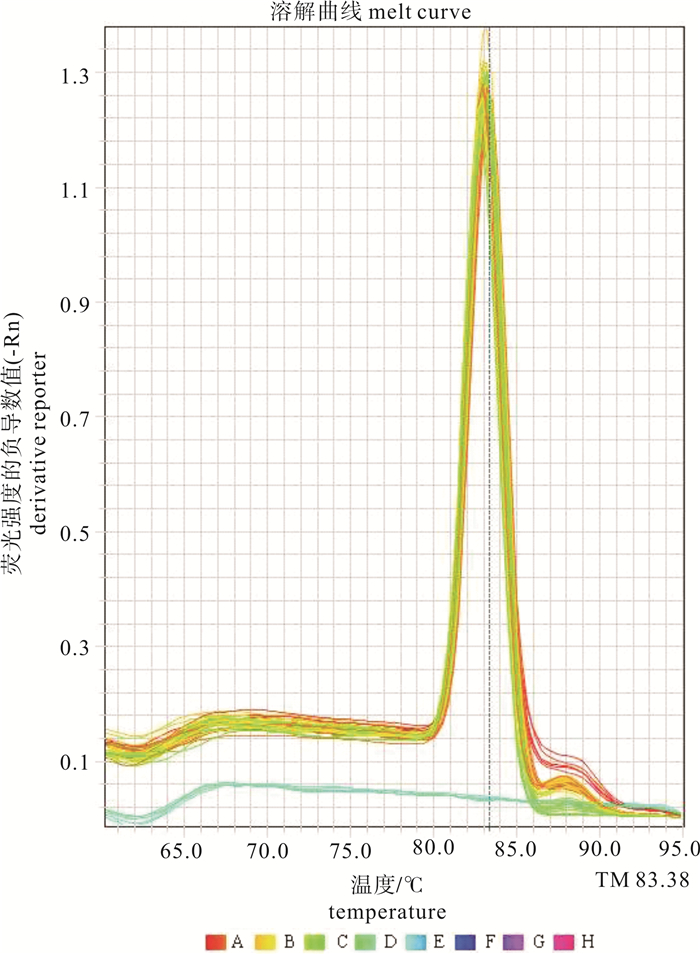
图 2 实时定量PCR扩增产物的熔解曲线
中文注解A~H.不同浓度质粒标准品(从高到低依次为2.0×109~2.0×102拷贝·μL-1);图中蓝色无峰熔解曲线来自阴性对照(无DNA酶水)alt-text>
A~H. A.portunus standards of different concentrations (from 2.0×109 to 2.0×102 copies·μL-1); the blue melting curve without a peak is the product of negative control (DNase-free ddH2O). Figure 2. Melting curves of real time qPCR products
![]()
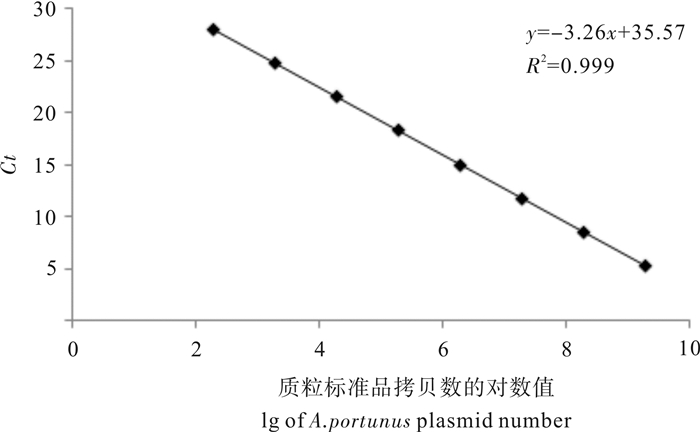
图 3 实时定量PCR方法的标准曲线
Figure 3. Standard curve of real time qPCR amplification method for A.portunus plasmid
![]()
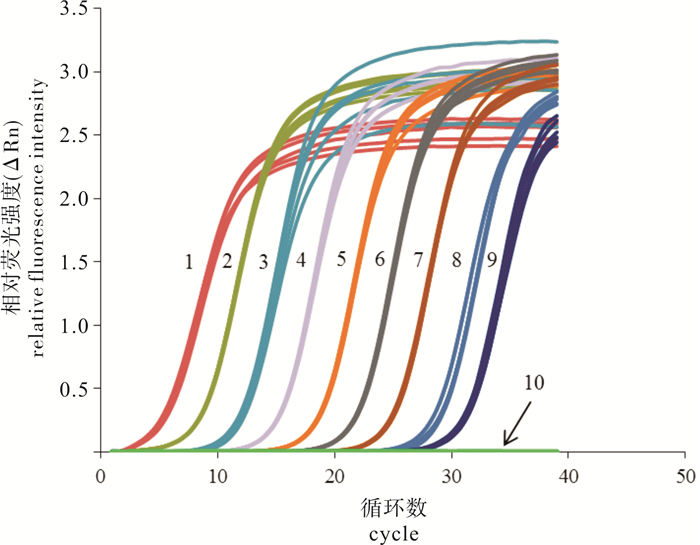
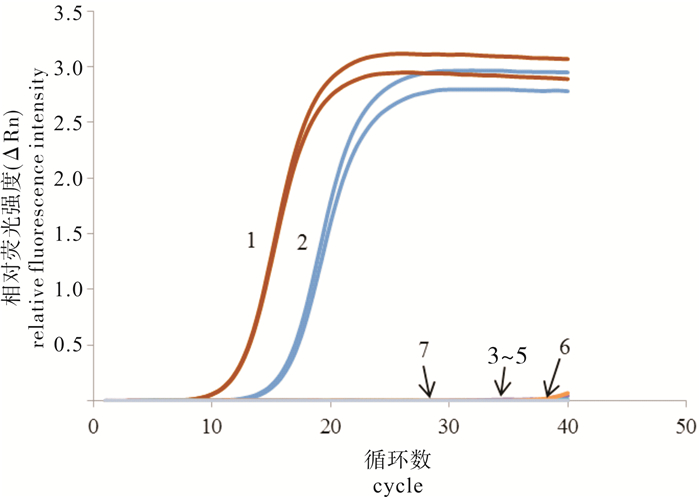
图 4 梭子蟹肌孢虫实时定量PCR方法的灵敏性试验
1~9.不同浓度质粒标准品(从高到低依次为2.0×109~2.0×101拷贝·μL-1);10.无DNA酶水
Figure 4. Sensitivity of real time qPCR method for A.portunus
1~9. A.portunus standards of different concentrations (from 2.0×109 to 2.0×101 copies·μL-1); 10. DNase-free ddH2O
![]()
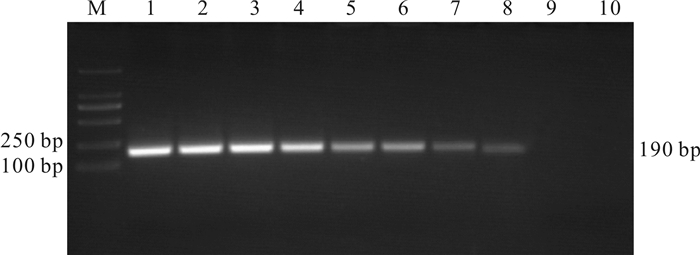
图 5 梭子蟹肌孢虫质粒标准品的套式PCR扩增结果
1~9.不同浓度质粒标准品(从高到低依次为2.0×109~2.0×101 拷贝·μL-1);10.无DNA酶水;M. DL2000 marker
Figure 5. Nested PCR products of A.portunus standards
1~9. A.portunus plasmids of different concentrations (from 2.0×109 to 2.0×101 copies·μL-1); 10. DNase-free ddH2O
![]()
图 6 梭子蟹肌孢虫实时定量PCR方法的特异性试验
1.质粒标准品(2.0×106 拷贝·μL-1);2. “牙膏蟹”肌肉;3. WSSV;4.健康三疣梭子蟹肌肉;5.溶藻弧菌;6.副溶血弧菌;7.无DNA酶水
Figure 6. Specificity of real time qPCR method for A.portunus
1. plasmid (2.0×106 copies·μL-1); 2. muscle of "toothpaste crab"; 3. WSSV; 4. muscle of the healthy P.trituberculatus; 5. V.alginolyticus; 6. V.parahemolyticus; 7. DNase-free ddH2O
表 1 实时定量PCR检测不同浓度质粒标准品的组内重复性
Table 1 Intra-group variability of real time qPCR for A.portunus plasmid
质粒/拷贝·μL-1
plasmid质粒Ct值
Ct values of parallel tests of plasmidsCt平均值
mean Ct of intra-group标准偏差
S.D.变异系数/%C.V. 1 2 3 4 5 2.0×109 5.26 5.16 5.21 5.25 5.24 5.22 0.04 0.77 2.0×108 8.48 8.45 8.52 8.46 8.49 8.48 0.03 0.35 2.0×107 11.69 11.93 11.82 11.67 11.59 11.74 0.13 1.11 2.0×106 15.22 14.93 15.14 14.71 15.09 15.02 0.20 1.33 2.0×105 18.35 18.23 18.18 18.11 18.47 18.27 0.14 0.77 2.0×104 21.42 21.59 21.65 21.49 21.51 21.53 0.09 0.42 2.0×103 24.88 24.85 24.96 24.70 24.56 24.79 0.16 0.65 2.0×102 28.12 28.04 28.03 28.21 27.90 28.06 0.12 0.43  下载: 导出CSV
下载: 导出CSV
表 2 实时定量PCR检测不同浓度质粒标准品的组间重复性
Table 2 Inter-group variability of real time qPCR for A.portunus standards
质粒/拷贝·μL-1
plasmidF P 2.0×109 0.65 0.33 2.0×108 0.09 0.64 2.0×107 0.41 0.13 2.0×106 0.25 0.10 2.0×105 0.16 0.23 2.0×104 0.19 0.53 2.0×103 0.23 0.52 2.0×102 0.12 0.70
下载: 导出CSV
表 3 养殖三疣梭子蟹中梭子蟹肌孢虫的检测结果
Table 3 Detection of A.portunus DNA in cultured P.trituberculatus samples
养殖户
company样品编号
sample No.套式PCR
nested PCRqPCR
拷贝·(ngMuDNA)-1qPCR
拷贝·(mgMu)-1蟹状态
status of crabM 1 + 4.92×108 3.15×1010 D 2 + 4.07×108 4.07×1010 D 3 + 3.89×102 2.47×104 D 4 - 2.69 1.74×102 D 5 + 2.10×104 1.65×106 D 6 + 1.43×108 1.04×1010 D 7 + 3.48×108 2.27×1010 D 8 + 1.16×104 8.48×105 D L 9 + 3.00×102 1.98×104 A 10 - 5.85 3.67×102 A 11 + 1.15×106 8.60×107 A 12 + 3.70×106 2.67×108 A Z 13 - 4.55 3.43×102 A 14 + 5.33×107 4.89×109 D 15 - NA NA A 16 - NA NA A 17 - NA NA A 注:+.阳性;-.阴性;D.死;A.活;Mu.肌肉;NA.无扩增
Note:+. positive;-. negative;D. dead;A. alive;Mu. muscle;NA.no amplification
下载: 导出CSV
-
[1] 王元. 脊尾白虾与三疣梭子蟹微孢子虫病的病原和病理[D]. 上海: 上海海洋大学, 2013: 28-42. http://cdmd.cnki.com.cn/Article/CDMD-10264-1014013589.htm [2] 王浩, 王元, 房文红, 等.微孢子虫感染三疣梭子蟹的肌组织病理及其免疫相关酶活性[J].海洋渔业, 2015, 37(5):457-464. http://www.doc88.com/p-6611231561566.html [3] RYAZANOVA T V, ELISEIKINA M G.Microsporidia of the genera Thelohania (Thelohaniidae) and Ameson (Pereziidae) in two species of lithodid crabs from the Sea of Okhotsk[J].Russ J Mar Biol, 2010, 36(6):435-442. doi: 10.1134/S1063074010060052
[4] VIVARES C P, AZEVEDO C.Ultrastructural observations of the life cycle stages of Ameson atlanticum sp.nov., a microsporidan parasitizing Cancer pagurus L.[J].J Fish Dis, 1988, 11(5):379-387. doi: 10.1111/jfd.1988.11.issue-5
[5] VIDEIRA M, CASAL G, ROCHA S, et al.Potaspora aequidens n. sp. (Microsporidia, Tetramicridae), a parasite infecting the freshwater fish Aequidens plagiozonatus (Teleostei, Cichlidae) from Brazil[J].Parasitol Res, 2015, 114(7):2435-2442. doi: 10.1007/s00436-015-4438-7
[6] STENTIFORD G D, FEIST S W, STONE D M, et al.Microsporidia:diverse, dynamic, and emergent pathogens in aquatic systems[J].Trends Parasitol, 2013, 29(11):567-578. doi: 10.1016/j.pt.2013.08.005
[7] 刘吉平, 曾玲.Calcofluor White M2R荧光染色法识别家蚕微孢子虫[J].昆虫学报, 2007, 50(11):1185-1186. doi: 10.3321/j.issn:0454-6296.2007.11.015 [8] 秦浩然, 李继莲, 和绍禹, 等.Calcofluor White M2R与SytoxGreen双重染色法鉴别蜜蜂微孢子虫[J].应用昆虫学报, 2012, 49(5):1392-1396. doi: 10.7679/j.issn.2095-1353.2012.205 [9] PURIVIROJKUL W, KHIDPRASERT S.First report of microsporidiosis in fairy shrimp Branchinella thailandensis (Sanoamuang, Saengphan and Murugan, 2002)[J].Aquaculture, 2009, 289(1/2):185-190. https://www.cabdirect.org/cabdirect/abstract/20093279266
[10] WANG W, CHEN J.Ultrastructural study on a novel microsporidian, Endoreticulatus eriocheir sp. nov. (Microsporidia, Encephalitozoonidae), parasite of Chinese mitten crab, Eriocheir sinensis (Crustacea, Decapoda)[J].J Invertebr Pathol, 2007, 94(2):77-83. doi: 10.1016/j.jip.2006.09.008
[11] GARCIA L S.Laboratory identification of the microsporidia[J].J Clin Microbiol, 2002, 40(6):1892-1901. doi: 10.1128/JCM.40.6.1892-1901.2002
[12] 刘吉平, 曹阳, SMITH J E, 等.模拟感染家蚕微粒子病的蚕卵、蚕蛾PCR检测的初步研究[J].蚕业科学, 2004, 30(4):367-370. http://www.cqvip.com/QK/93797X/200404/11652322.html [13] TANG K F, PANTOJA C R, REDMAN R M, et al.Development of in situ hybridization and PCR assays for the detection of Enterocytozoon hepatopenaei (EHP), a microsporidian parasite infecting penaeid shrimp[J].J Invertebr Pathol, 2015, 130:37-41. doi: 10.1016/j.jip.2015.06.009
[14] SUEBSING R, PROMBUN P, SRISALA J, et al.Loop-mediated isothermal amplification combined with colorimetric nanogold for detection of the microsporidian Enterocytozoon hepatopenaei in penaeid shrimp[J].J Appl Microbiol, 2013, 114(5):1254-1263. doi: 10.1111/jam.2013.114.issue-5
[15] HAN J E, TANG K F, PANTOJA C R, et al.Detection of a new microsporidium Perezia sp. in shrimps Penaeus monodon and P. indicus by histopathology, in situ hybridization and PCR[J].Dis Aquat Organ, 2016, 120(2):165-171. doi: 10.3354/dao03022
[16] HOPPER J V, HUANG W F, SOLTER L F, et al.Pathogenicity, morphology, and characterization of a Nosema fumiferanae isolate (Microsporidia:Nosematidae) from the light brown apple moth, Epiphyas postvittana (Lepidoptera:Tortricidae) in California[J].J Invertebr Pathol, 2016, 134:38-47. doi: 10.1016/j.jip.2016.01.001
[17] KLEE J, TEK TAY W, PAXTON R J.Specific and sensitive detection of Nosema bombi (Microsporidia:Nosematidae) in bumble bees (Bombus spp.; Hymenoptera:Apidae) by PCR of partial rRNA gene sequences[J].J Invertebr Pathol, 2006, 91(2):98-104. doi: 10.1016/j.jip.2005.10.012
[18] TRIPODI A D, CIBILS-STEWART X, MCCORNACK B P.No-sema bombi (Microsporidia:Nosematidae) and trypanosomatid prevalence in spring bumble bee queens (Hymenoptera:Apidae:Bombus) in Kansas[J].J Kansas Entomol Soc, 2014, 87(2):225-233. doi: 10.2317/JKES130730.1
[19] VOSSBRINCK C R, DEBRUNNER-VOSSBRINCK B A.Molecular phylogeny of the Microsporidia:ecological, ultrastructural and taxonomic considerations[J].Folia Parasitol (Praha), 2005, 52(1/2):131-142.
[20] 刘珍, 张庆利, 万晓媛, 等.虾肝肠胞虫(Enterocytozoon hepatopenaei)实时荧光定量PCR检测方法的建立及对虾样品的检测[J].渔业科学进展, 2016, 37(2):119-126. doi: 10.11758/yykxjz.20150512003 [21] 王瑞, 李莉萍, 黄婷, 等.罗非鱼组织内无乳链球菌实时荧光定量PCR检测方法建立[J].南方水产科学, 2015, 11(3):41-46. http://www.cnki.com.cn/Article/CJFDTOTAL-NFSC201503007.htm [22] 李新苍, 周俊芳, 房文红, 等.实用WSSV定量检测方法的建立及其应用于脊尾白虾病毒感染规律的研究[J].水产学报, 2012, 36(10):1554-1562. http://www.oalib.com/paper/5090013 [23] FU Z, HE X, CAI S, et al.Quantitative PCR for detection of Nosema bombycis in single silkworm eggs and newly hatched larvae[J].J Microbiol Methods, 2016, 120:72-78. doi: 10.1016/j.mimet.2015.12.003
[24] POLLEY S D, BOADI S, WATSON J, et al.Detection and species identification of microsporidial infections using SYBR Green real-time PCR[J].J Med Microbiol, 2011, 60(Pt4):459-466.
[25] DURAND S V, LIGHTNER D V.Quantitative real time PCR for the measurement of white spot syndrome virus in shrimp[J].J Fish Dis, 2002, 25(7):381-389. doi: 10.1046/j.1365-2761.2002.00367.x
[26] PHELPS N B, GOODWIN A E.Validation of a quantitative PCR diagnostic method for detection of the microsporidian Ovipleistophora ovariae in the cyprinid fish Notemigonus crysoleucas[J].Dis Aquat Organ, 2007, 76(3):215-221. https://ar.scribd.com/document/274194487/AQFI-PhD-Proposal
[27] VIVARES C P, SPRAGUE V.The fine structure of Ameson pulvis(Microspora, Microsporida)and its implications regarding classification and chromosome cycle[J].J Invertebr Pathol, 1979, 33(1):40-52. doi: 10.1016/0022-2011(79)90128-9
[28] FINDLEY A M, BLAKENEY E W, WEIDNER E H.Ameson michaelis(Microsporida)in the blue crab, Callinectes sapidus:parasite-induced alterations in the biochemical composition of host tissues[J].Biol Bull, 1981, 161(1):115-125. doi: 10.2307/1541112
[29] SMALL H J, MEYER G R, STENTIFORD G D, et al.Ameson metacarcini sp. nov. (Microsporidia) infecting the muscles of Dungeness crabs Metacarcinus magister from British Columbia, Canada[J].Dis Aquat Organ, 2014, 110(3):213-225. doi: 10.3354/dao02754
[30] PALENZUELA O, REDONDO M J, CALI A, et al.A new intranuclear microsporidium, Enterospora nucleophila n. sp., causing an emaciative syndrome in a piscine host (Sparus aurata), prompts the redescription of the family Enterocytozoonidae[J].Int J Parasitol, 2014, 44(3/4):189-203.
[31] HOCH G, SOLTER L F, SCHOPF A.Hemolymph melanization and alterations in hemocyte numbers in Lymantria disparlarvae following infections with different entomopathogenic microsporidia[J].Entomol Exp Appl, 2004, 113(2):77-86. doi: 10.1111/eea.2004.113.issue-2



计量
- 文章访问数: 3276
- HTML全文浏览量: 1135
- PDF下载量: 429
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
