
A preliminary analysis on biological characteristics of Katsuwonus pelamis in the Nansha Islands sea area in spring
-
摘要:
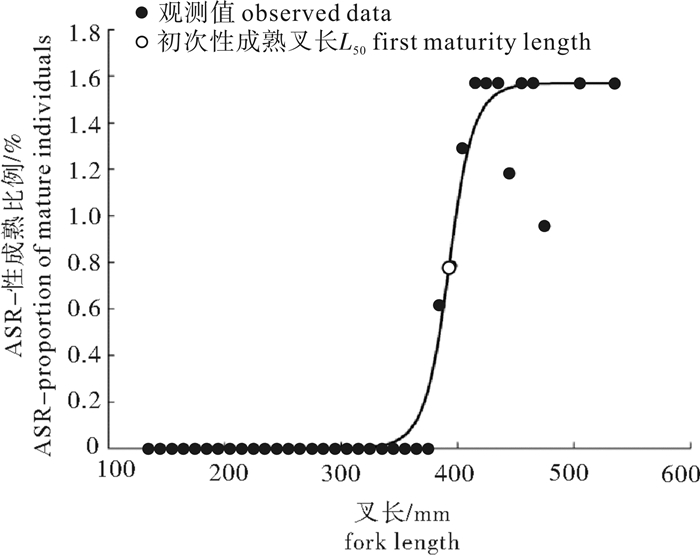
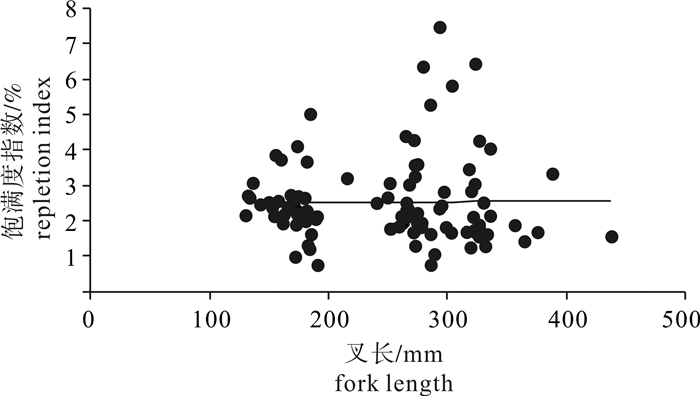
根据2014年5月和2016年4月灯光罩网船在南沙海域生产调查采集的鲣(Katsuwonus pelamis)样本,对其渔业生物学特性进行初步研究。结果显示,渔获物优势叉长为260~280 mm,占14.68%;优势体质量为27~100 g,占18.35%;叉长与体质量关系参数b值为3.403 9。雌、雄性比约为1 : 0.9;性腺成熟度以Ⅰ期(47.71%)和Ⅳ期(27.06%)为主,随着叉长的增大,雄性个体所占比例上升。性腺发育个体性腺长与叉长呈显著性正相关关系(P < 0.01);初次性成熟叉长为392.70 mm;摄食等级以1级和2级为主,分别占36.24%和29.36%,1~4级有逐渐减少的变化趋势;饱满度指数为0.74~7.45,均值为2.53。
Abstract:We investigated the biological characteristics of Katsuwonus pelamis which were collected by light-falling net fishery in the Nansha Islands sea area in May 2014 and April 2016. The results show that the dominant fork length ranged from 260 mm to 280 mm, accounting for 14.68% of the total samples. The dominant body weight ranged from 27 g to 100 g, accounting for 18.35% of the total samples. The somatic growing parameters (b value) between fork length and body weight was 3.403 9. The sex ratio of female to male was 1 : 0.9, and the sexual maturity was higher at Stage Ⅰ (47.71%) and Ⅳ (27.06%), and the proportion of males increased with increasing fork length. Gonad length was positively correlated with fork length significantly (P < 0.01). The fork length of the first gonadal maturity was 392.70 mm. Stage 1 and Stage 2 had the highest feeding intensities, accounting for 36.24% and 29.36% of the total samples, respectively. A descending trend was found from Stage 1 to Stage 4. The plumpness index ranged from 0.74 to 7.45 (averagely 2.53).
-
任何形式的捕捞均有选择性, 其选择性主要取决于渔具选择性,它强烈影响着捕捞群体结构。为了实现渔业资源的可持续利用,开展生态渔业、负责任渔业的研究有重要的意义,渔具选择性和选择性渔具是生态渔业的核心内容之一。而在我国开展的渔具选择性研究尤其是定置渔具选择性能的研究并不多见。
日本沿海主要以小型底拖网、笼、筒捕捞星康吉鳗Conger myriaster(以下简称星鳗)。在我国星鳗也是重要的群众渔业捕捞对象,青岛近海渔民用筒、山东日照渔民利用延绳钓捕捞这种鱼。山东沿海渔民在利用地笼诱捕许氏平鲉、欧氏六线鱼时, 往往同时捕获大量星鳗。本文以东京湾星鳗笼为例,利用套网法研究了其网目选择性,以期为定置渔业的科学管理提供科学依据。
1. 材料和方法
1.1 海上实验
1995~2000年,每年的10~11月在东京湾进行实验。实验笼和生产的相同,不锈钢骨架,展开为640 mm×470 mm×150 mm。每个笼有2个椭圆形开口200 mm×90 mm, 和东京湾渔获对象星鳗(全长190~600 mm, 最大体周长28~104 mm)相比, 开口非常大。因此在研究笼的选择性时, 可以仅考虑网目的选择性。
笼的底部网目尺寸(内径)为10.8 mm,远小于其他部位的网目尺寸(表 1), 星鳗从笼中逃逸的部位是底部以外的其他部分。实验时,笼的底部以外结覆网目内径为10.8 mm的套网, 套网和笼之间有足够大的空间, 且所有套网的规格相同。套网、笼均为锦纶制造,有结节菱形网片。实验时的作业方法和生产相同, 采用延绳钓作业方式。
表 1 实验笼的数量以及网目尺寸(MC)Table 1. The number and mesh size of test net pot(MC)网目尺寸/mm mesh size 1995年 1996年 1997年 1998年 1999年 2000年 21.0±0.68 4 4 4 4 4 4 18.1±0.82 3 3 4 4 4 4 15.5±0.63 3 3 4 4 4 4 13.6±0.44 3 3 4 4 4 4 11.6±0.38 3 3 4 4 4 4 将笼网、套网捕获的星鳗分别装入化纤袋,回实验室测量全长、最大体周长,体重。最大体周长是胸鳍稍后鱼体最粗部位的周长。
1.2 解析方法
如果1尾鱼被捕获的概率是p, 那么Ni尾行为独立有相同特征的鱼遭遇笼,ni尾鱼被捕获的概率$p=C_{N_i}^{n_i} p^{n_i}(1-\$p)^{N_i-n_i}$。
假设1群鱼遭遇渔具, 该鱼群依据全长可以划分为k组,组中值为li,每组鱼Ni,每组被捕的数量mi,那么$\sum_\limits{i=1}^k$尾鱼被捕获的概率$F=\prod_\limits{i=i}^k C_{N_i}^{n_i} p^{n_i}(1-p)^{N_i-n_i}$。
如果套网和笼捕获的鱼的数量为nli, mli(i=1,2,3,…,k), 那么笼捕获的全长li鱼的占该类鱼总渔获量的比例为
$$ \phi_{l_i}=\frac{n_{l_i}}{n_{l_i}+m_{l_i}} $$ (1) 依据概率的统计学定义, F可用$\phi_{l_i}$估计, 而p是渔具的选择率SL[1], 可用Logistic方程表示[2],
$$ S_L=\frac{\exp (s+b L)}{1+\exp (s+b L)} $$ (2) a,b为待估参数,L为鱼类的特征变量(全长l,相对体周长$\frac{G}{M_e}$等)。由F和$\phi_{l_i}$, p和SL的关系, 利用极大似然估计法可以得到待估参数a,b的值。
由不同网目尺寸得到的不同的网目选择性曲线, 利用不同的处理方式可以得到不同的主选择性曲线。为比较这些曲线, 引入AIC(akayike information criterion),AIC=-2 max {ln F}+2K作为判断标准,为待估参数的数目,AIC较小的模型更合适[3]。
2. 结果与讨论
2.1 星鳗的体重、全长,最大体周长间的关系
捕获的星鳗的全长l(cm)与体重w(g)、体周长G(cm)的关系分别为,
w=0.1998l2-7.9106l+93.5290(R2=0.9610, n=2 102),
G=0.1906l-0.6786(R2=0.8488, n=2 102)。
2.2 全长选择性曲线
笼的选择性由2部分组成,网目选择性、出入口的选择性。在东京湾可仅考虑网目的选择性[4]。以全长为方程2的变量,由算式1, 2得到的网目选择性曲线的各个参数(表 2)。
表 2 全长选择性曲线的各参数Table 2. Parameters of whole body length selectivity curve网目尺寸/mm mesh size a b MLL AIC l0.5 S.R. 21.0 -17.26 0.58 -22.01 48.02 29.80 3.80 18.1 -13.83 0.52 -37.65 79.30 26.80 4.30 15.5 -14.44 0.63 -56.77 117.54 23.00 3.50 13.6 -16.11 0.73 -53.71 111.42 22.00 3.00 11.6 -14.65 0.73 -173.71 351.41 20.00 3.00 注: l0.5, 50%选择全长; S.R., 选择域; MLL=Max{lnF}
Notes:l0.5, 50% selective whole body length; S.R., selective range; MLL=Max{lnF}由上述的各参数可知,网目尺寸21.0、18.1、15.5、13.6、11.6 mm笼的全长选择性能,全长0 < l < 25.50 cm、0 < l < 22.50 cm、0 < l < 18.50 cm、0 < l < 18.50 cm、0 < l < 16.50 cm时,全长选择率几乎为0;全长25.50 < l < 35.50 cm、22.50 < l < 32.50 cm、18.50 < l < 27.50 cm、18.50 < l < 26.50 cm、16.50 < l < 24.00 cm时,全长选择率随全长不断增长而逐渐增大;全长在l>35.50 cm、l>32.50 cm、l>27.50 cm、l>26.50 cm、l>24.00 cm时,全长选择率几乎接近于1.00,50%选择全长l0.5分别为29.8、26.8、23.0、22.0、20.0 cm;全长选择域为3.80、4.30、3.50、3.00、3.00 cm。因此,全长选择性曲线沿全长增加方向,随网目增大而向右移,并且网目越小,选择越尖锐。
笼的网目尺寸分别为21.0、18.1、15.5、13.6、11.6 mm,和套网的网目尺寸的比值依次为1.07、1.26、1.44、1.68、1.94。所对应的AIC逐渐减少(表 2)。由于实验中,除笼的网目尺寸有差别外,其他条件尽可能相同,因此造成这种趋势的原因很可能来自套网对实验笼的影响(例如局部流场的改变程度),即可能是在一定范围内,实验网和套网的网目尺寸差别越大,套网对实验网的影响越小;而当两者相当时,套网的影响相当显著。是否是这种原因,有待于更多关于鱼类行为以及渔具和捕捞对象相互作用的实验检验。
2.3 主选择性曲线
鱼能否穿过网目,主要取决于鱼沿体高方向的截面形状以及相对于网目内径的大小[4]。由或为方程1的变量,所得到的方程为主选择性方程,它描绘的曲线为主选择性曲线(master selectivity curve)。有2种方法,each curve model和single curve model可得到主选择性曲线。前者是求得每一网目尺寸所对的主选择性曲线。后者将所有的网目尺寸所对应的相对体周长$\frac{G}{M_c}$或相对全长$\frac{l}{M_c}$值合并,看作一组数值,求得一条反映渔具选择性的曲线。
以$\frac{G}{M_c}$为方程2的变量,由single curve model和each curve model,利用极大似然估计法,得到待估参数(表 3)。因∑ (AICeach curve model)=250.18,而AICsingle curve model=244.32,其差值│244.32-250.18│>1,所以可以认为single curve model与each curve model之间的差异并非偶然,single curve model得到的主选择性曲线可更好地反映网目选择性。
表 3 主选择性曲线的各参数Table 3. Parameters of master selectivity curve模式models a b $\frac{G}{M_{e0.5}}$ S.R. MLL AIC single curve model -4.71 4.25 1.11 0.52 -120.16 244.32 each curve model- 115.09 250.18 21.0 mm -10.47 8.68 1.21 0.25 -16.96 37.92 18.1 mm -8.92 7.70 1.16 2.87 -24.45 52.90 15.5 mm -1.58 2.87 0.55 0.77 -24.54 53.07 13.6 mm -4.87 4.62 1.06 0.48 -16.95 37.91 11.6 mm -0.01×10-2 1.08 9.20×10-5 2.03 -32.19 68.38 50%选择相对体周长1.11,相对体周长的选择域为0.52。$0<\frac{G}{M_c}<0.55$,相对体周长选择率几乎为0;$0.55< \frac{G}{M_c}<1.60$,选择率随相对体周长增加而增加;$\frac{G}{M_c}>1.60$,选择率趋向于1.00。一般认为当$\frac{G}{M_c}=1$时,选择率就应当趋向于1.0,而本实验当$\frac{G}{M_c}>1.60$,选择率才趋向1.00。这种结果和星鳗底拖网的网目选择性中的类似[5]。其最可能的原因是星鳗体表鳞片退化,凸起减少,且分泌大量粘液,使之容易穿过网目。另外,网线对星鳗的刺激,也有促进星鳗积极逃逸的可能。
3. 小结
比较显示,套网和实验网的网目尺寸差别较大时,套网的影响较小。而在两者接近时,套网的影响极为显著。
此外,本文中给出了利用套网法得到的星鳗笼的网目选择性能, 但套网对入笼的星鳗行为存在影响, 这种影响到底有多大, 以及入网的星鳗如何从笼中逃逸, 逃逸后的残存率有多大, 都是将来要研究的重点所在。
-
![]()
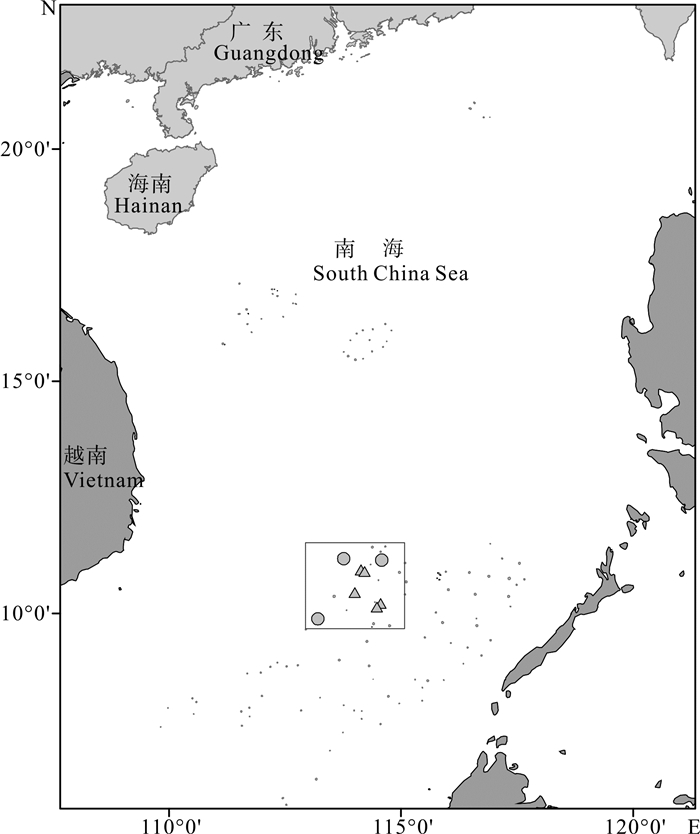
图 1 样本采集海域三角形和圆点分别代表2014年和2016年样品采集站位。
Figure 1. Sampling sea area Triangles and dots indicate sampling sites in 2014 and 2016, respectively.
![]()
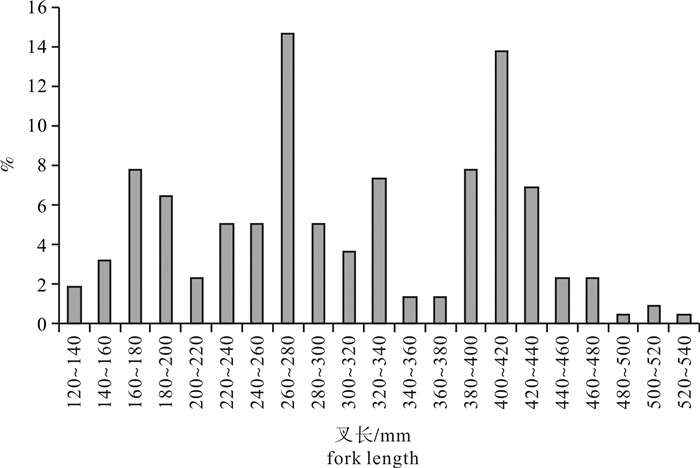
图 2 春季南沙海域鲣叉长分布
Figure 2. Distribution of fork length of K.pelamis in the Nansha Islands sea area in spring
![]()
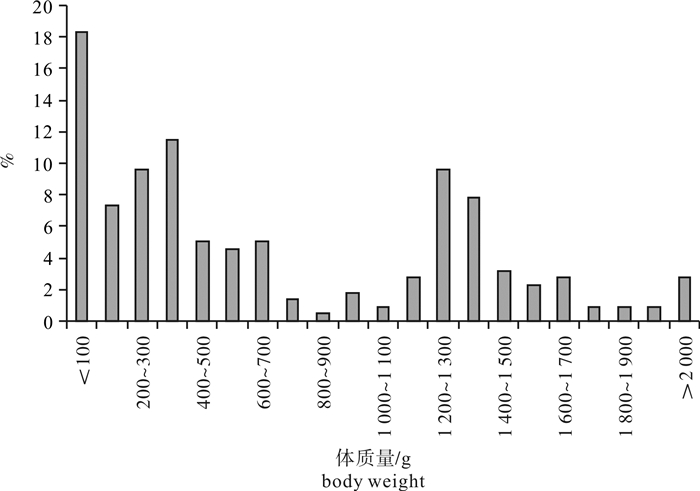
图 3 春季南沙海域鲣体质量分布
Figure 3. Distribution of body weight of K.pelamis in the Nansha Islands sea area in spring
![]()
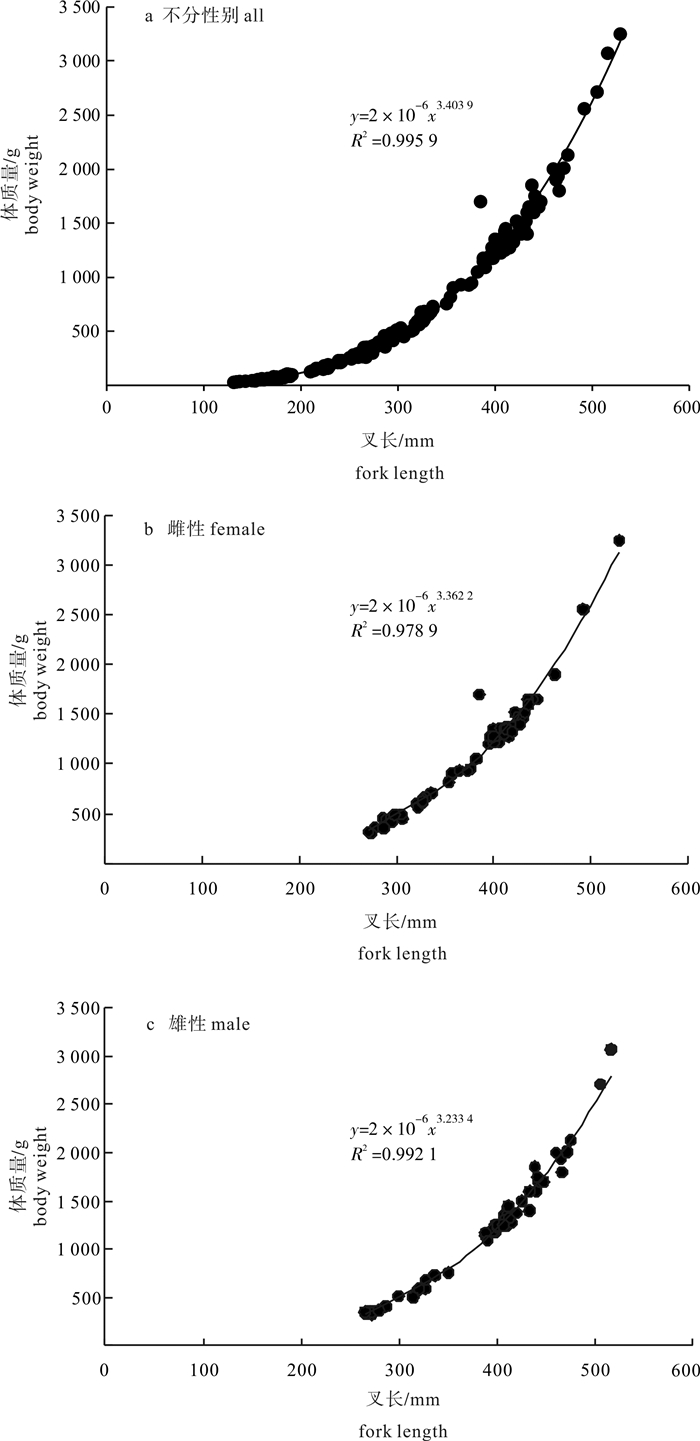
图 4 春季南沙海域鲣叉长与体质量关系
Figure 4. Relationship between fork length and bodyweight of K.pelamis in the Nansha Islands sea area in spring
![]()
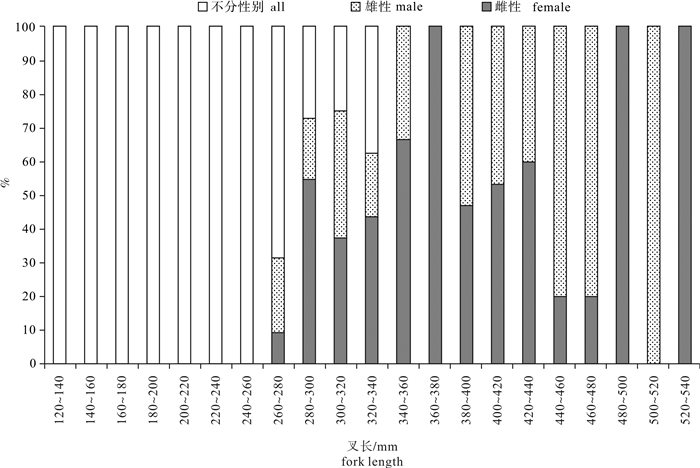
图 5 春季南沙海域鲣性比与叉长关系
Figure 5. Relationship between sex ratio and fork length of K.pelamis in the Nansha Islands sea area in spring
![]()
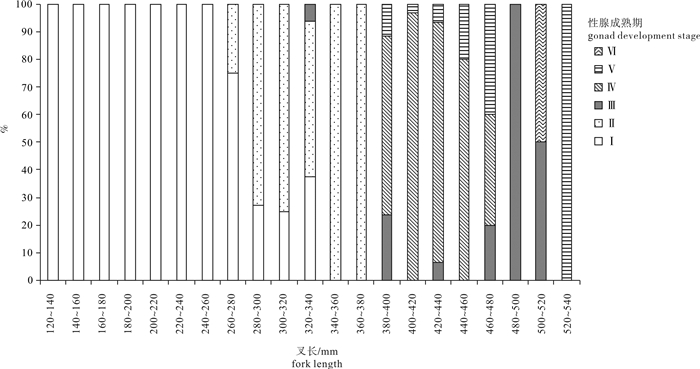
图 6 春季南沙海域鲣不同叉长组性腺成熟度百分比组成
Figure 6. Percentage of gonad development stages in different fork length groups of K.pelamis in the Nansha Islands sea area in spring
![]()
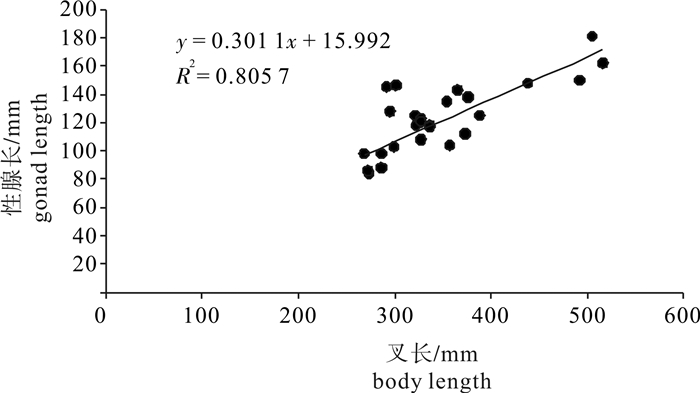
图 7 春季南沙海域鲣叉长与性腺长关系
Figure 7. Relationship between gonad length and body length of K.pelamis in the Nansha Islands sea area in spring
![]()
图 8 春季南沙海域鲣性成熟比例观测值与ASR模型拟合的逻辑斯蒂曲线
Figure 8. Observed proportion of mature individuals and logistic curve fitted by ASR model of K.Pelamis in the Nansha Islands sea area in spring
![]()
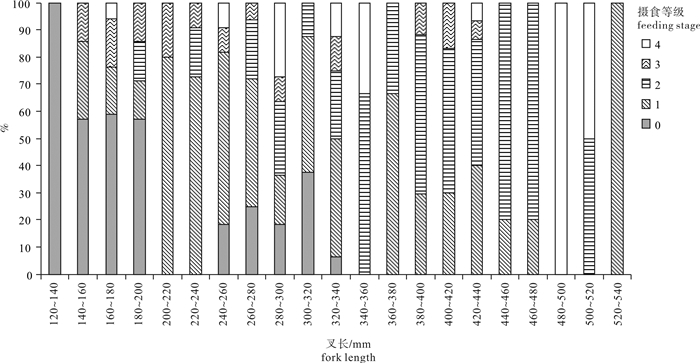
图 9 春季南沙海域鲣摄食等级与叉长关系
Figure 9. Relationship between feeding stages and fork length of K.pelamis in the Nansha Islands sea area in spring
-
[1] 黄锡昌, 苗振清.远洋金枪鱼渔业[M].上海:上海科学技术文献出版社, 2003:55-60. [2] 张仁斋.三种金枪鱼类(鲣、黄鳍金枪鱼、扁舵鲣)的仔、稚鱼在南海的分布与产卵期[J].海洋学报, 1983, 5(3):368-375. http://www.oalib.com/paper/4563791 [3] 勒少非, 樊伟.鲣鱼资源开发利用研究现状及未来气候变化背景下研究展望[J].渔业信息与战略, 2014, 29(4):272-279. http://www.cnki.com.cn/Article/CJFDTotal-XYYZ201404006.htm [4] THANH H. Ocean tuna fishing and marketing in Vietnam [J].Vietfish, 2009, 6(1):56-59. http://www.un.org/Depts/los/nippon/unnff_programme_home/fellows_pages/fellows_papers/nasuchon_0809_thailand.pdf
[5] 张青, 王锡昌, 刘源.中国金枪鱼渔业现状及发展趋势[J].南方水产, 2009, 5(1):68-74. http://www.schinafish.cn/CN/Y2009/V5/I1/68 [6] 张鹏, 张俊, 李渊, 等.秋季南海中南部海域的一次灯光罩网探捕调查[J].南方水产科学, 2016, 12(2):67-74. http://kns.cnki.net/KCMS/detail/detail.aspx?dbcode=CJFQ&dbname=CJFDLAST2016&filename=NFSC201602010&v=MzE3NzBoMVQzcVRyV00xRnJDVVJMMmVaZWR1Rnkvblc3N0JLeXZZYmJHNEg5Zk1yWTlFWklSOGVYMUx1eFlTN0Q= [7] 杨晓明, 戴小杰, 田思泉, 等.中西太平洋鲣鱼围网渔业资源的热点分析和空间异质性[J].生态学报, 2014, 34(13):3771-3778. http://www.ecologica.cn/stxb/ch/html/2014/13/stxb201211101579.htm [8] 汪金涛, 陈新军.中西太平洋鲣鱼渔场的重心变化及其预测模型建立[J].中国海洋大学学报, 2013, 43(8):44-48. http://industry.wanfangdata.com.cn/dl/Detail/Periodical?id=... [9] 叶泰豪, 冯波, 颜云榕, 等.中西太平洋鲣渔场与温盐垂直结构关系的研究[J].海洋湖沼通报, 2012, 6(1):49-55. http://www.cqvip.com/QK/95761X/201201/41392554.html [10] 胡奎伟, 朱国平, 王学昉, 等.中西太平洋鲣鱼丰度的时空分布及其与表温的关系[J].海洋渔业, 2011, 33(4):417-422. http://www.cqvip.com/QK/93342X/201104/40142978.html [11] 杨胜龙, 周甦芳, 周为峰, 等.基于Argo数据的中西太平洋鲣渔获量与水温、表层盐度关系的初步研究[J].大连水产学院学报, 2010, 25(1):34-40. http://www.doc88.com/p-7945823428983.html [12] 苏阳, 章超桦, 曹文红, 等.南海产3种金枪鱼普通肉、暗色肉营养成分分析与评价[J].广东海洋大学学报, 2015, 35(3):87-93. http://www.cqvip.com/QK/93246X/201503/665453271.html [13] 童铃, 金毅, 徐坤华, 等. 3种鲣鱼背部肌肉的营养成分分析及评价[J].南方水产科学, 2014, 10(5):51-59. http://kns.cnki.net/KCMS/detail/detail.aspx?dbcode=CJFQ&dbname=CJFD2014&filename=NFSC201405009&v=MTcwNDRiYkc0SDlYTXFvOUZiWVI4ZVgxTHV4WVM3RGgxVDNxVHJXTTFGckNVUkwyZVplZHVGeS9uVzd6SUt5dlk= [14] 高萌, 张宾, 王强, 等.流化冰保鲜对鲣鱼蛋白质功能特性的影响[J].食品科学, 2014, 35(22):304-309. doi: 10.7506/spkx1002-6630-201422059 [15] 王春雷, 许柳雄, 叶旭昌, 等.中西太平洋金枪鱼围网起水鱼群与流木鱼群渔获物中鲣生物学特性的比较[J].大连水产学院学报, 2008, 23(3):235-241. https://wenku.baidu.com/view/ad6f9359be23482fb4da4cbf.html [16] 费鸿年, 张诗全.水产资源学[M].北京:中国科学技术出版社, 1990:114-285. [17] KING M. Fisheries biology, assessment and management [M].New York:John Wiley & Sons, 2013:1-400.
[18] 李忠炉, 金显仕, 单秀娟, 等.小黄鱼体长-体质量关系和肥满度的年际变化[J].中国水产科学, 2011, 18(3):602-610. https://wenku.baidu.com/view/e128d88eb9d528ea81c77968.html [19] CHEN Y, PALOHEIMO J E. Estimating fish length and age at 50% maturity using a logistic type model [J].Aquat Sci, 1994, 56(3):206-219. doi: 10.1007/BF00879965
[20] 许柳雄, 王学昉, 朱国平, 等.中西太平洋金枪鱼围网鲣鱼起水鱼群种群结构分析[J].海洋湖沼通报, 2010, 16(2):22-28. http://www.wenkuxiazai.com/doc/6284188202d276a200292ebf-3.html [21] 杨松, 陈新军, 许柳雄.中西太平洋金枪鱼围网渔业渔获组成及叉长与体重关系[J].上海水产大学学报, 2005, 14(3):337-340. https://wenku.baidu.com/view/ad6f9359be23482fb4da4cbf.html [22] BATTS B S. Age and growth of the skipjack tuna, Katsuwonus pelamis (Linnaeus), in North Carolina waters [J].Chesap Sci, 1972, 13(4):237-244. doi: 10.2307/1351107
[23] 詹秉义.渔业资源评估[M].北京:中国农业出版社, 1995:20-21. [24] ANDRADE H A, CAMPOS R O. Allometry coefficient variations of the length-weight relationship of skipjack tuna (Katsuwonus pelamis) caught in the southwest South Atlantic[J].Fish Res, 2002, 55(1/2/3):307-312. http://www.academia.edu/...OF_AQUACULTURE_SPECIES_AND_ENHANCEMENT_OF_STOCKS
[25] MATSUMOTO W M, SKILLMAN R A, DIZON A E. Synopsis of biological data on skipjack tuna, Katsuwonus pelamis [J].NOAA Techn Rep NMFS, 1984, 451:1-92. doi: 10.1007/BF00428558.pdf
[26] 许柳雄, 王学昉, 朱国平, 等.中西太平洋金枪鱼围网流木鱼群中鲣鱼种群结构分析[J].生态学杂志, 2009, 28(2):293-299. http://image.sciencenet.cn/olddata/kexue.com.cn/upload/blog/... [27] BROCK V E. A preliminary report on Parathunnus sibi in Hawaiian waters and a key to the tunas and tuna-like fishes of Hawaii[J].Pacific Sci, 1949, 3(3):271-277. http://scholarspace.manoa.hawaii.edu/bitstream/handle/10125/8934/vol3n3-271-277.pdf?sequence=1
[28] GRANDE M, MURUA H, ZUDAIRE I, et al. Reproductive timing and reproductive capacity of the skipjack tuna (Katsuwonus pelamis) in the western Indian Ocean [J].Fish Res, 2014, 156(4):14-22. http://www.iotc.org/documents/reproductive-biology-skipjack-tuna...
[29] MARR J C. Observation on the spawning of oceanic skipjack tuna (Katsuwonus pelamis) and yellowfin tuna (Neothunnus macropterus) in the northern Marshall Islands [J].Fish Bull, 1948, 51(44):201-206.
[30] ASHIDA H, TANABE T, SATOH K, et al. Reproductive biology of male skipjack tuna Katsuwonus pelamis (Linnaeus) in the tropical western and central Pacific Ocean[J].Fish Sci, 2010, 76(5):785-793. doi: 10.1007/s12562-010-0282-y
[31] SCHAEFER M B, ORANGE C J. Studies on the sexual development and spawning of yellow tuna (Neothunnus maropterus) and skipjack (Katsuwonus pelamis) in three areas of the eastern Pacific Ocean, by examination of gonads[J].Inter-Amer Trop Tuna Comm Bull, 1956, 19(6):281-349. http://scientiamarina.revistas.csic.es/index.php/scientiamarina/article/...
[32] VILELA M J A, CASTELLO J P. Dinámica poblacional del barrilete (Katsuwonus pelamis) explotado en la región sudeste-sur del Brasil en el periodo 1980-1986[J].Frent Mar, 1993, 14 (Sec.A):111-124. http://www.scielo.cl/scielo.php?script=sci_arttext&pid=S0718-560X2014000300013
[33] 王学昉, 许柳雄, 周成, 等.金枪鱼围网鲣的捕获率与其摄食状态的关系[J].大连海洋大学学报, 2013, 28(6):622-626. http://www.cqvip.com/QK/94241A/201306/1002795335.html [34] OFDC (Overseas Fisheries Development Council of Chinese Taipei). 世界鲣鲔围网渔业简介[EB/OL]. [2016-08-10]. http://www.ofdc.org.tw/catchstatus/02/WholeWorld_PURSE_Introdution.Pdf. -
期刊类型引用(1)
1. 张静,胡长圣,刘前,戴佳玥,王学锋,汤保贵. 流速对卵形鲳鲹游泳行为及运动生理的影响. 中国水产科学. 2024(04): 381-390 .  百度学术
百度学术
其他类型引用(0)
 下载:
下载:
计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 1
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
