
Community structure of phytoplankton in the Nansha sea area of Pearl River Estuary
-
摘要:
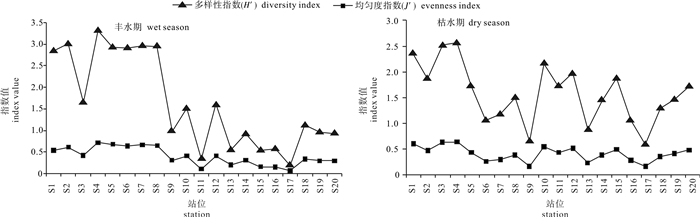
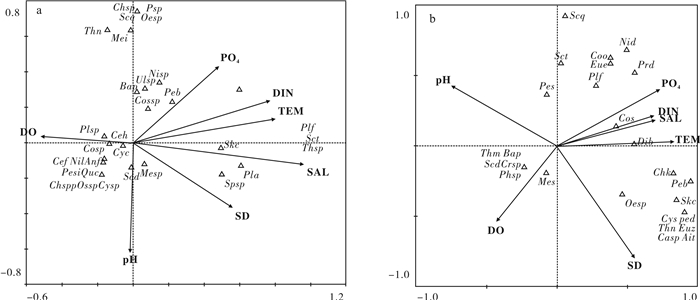
基于2015年4月(丰水期)和2015年11月(枯水期)珠江口南沙海域的调查资料,分析该海域浮游植物的种类组成、优势度、细胞丰度及多样性。共鉴定出浮游植物5门44属76种,硅藻为最主要优势种群。丰水期优势种有2种,分别为新月筒柱藻(Cylindrotheca closterium)和中肋骨条藻(Skeletonema costatum);枯水期优势种有3种,分别为萎软海链藻(Thalassiosira mala)、中肋骨条藻和有棘圆筛藻(Coscinodiscus spinosus)。丰水期和枯水期浮游植物细胞丰度分别为651.91×104个· L-1和129.21×104个· L-1,2个时期细胞丰度分布都表现为近岸向离岸递增的趋势。调查海区的浮游植物Shannon-Wiener多样性指数(H′)和Pielou均匀度指数(J′)的平均值在丰水期分别为1.64和0.40,而在枯水期分别为1.58和0.41,多样性总体处于较低水平。典范对应分析结果表明,丰水期影响浮游植物群落结构的主要环境因子为盐度、温度、总氮和pH,枯水期为温度、透明度、磷酸盐和盐度。
Abstract:We analyzed the species composition, dominant species, abundance and community diversity of phytoplankton based on the data collected from the ecological investigation of the Nansha sea area of Pearl River Estuary in both wet season (April, 2015) and dry season (November, 2015).A total of 76 species among 44 genera belonging to 5 phyla were identified in the survey area. Diatom was the predominant group of all the phyla. In wet season there were 2 dominant species, Cylindrotheca closterium and Skeletonema costatum. In dry season, there were 3 dominant species, Thalassiosira mala, S.costatum and Coscinodiscus spinosus. The average cell abundance of phytoplankton in wet and dry seasons was 651.91×104cells · L-1 and 129.21×104cells · L-1, respectively. The cell abundance distribution during the two seasons showed an increasing trend from inshore to offshore. In general, the Shannon-Wiener index (H′) and the Pielou evenness index (J′) were 1.64 and 0.40 in wet season, and 1.58 and 0.41 in dry season, suggesting low diversity of the phytoplankton community. Ordination diagram of CCA results indicate that salinity, temperature, DIN and pH are the main environmental factors which affect phytoplankton community in wet season, while temperature, transparency, phosphate and salinity are the main affecting factors in dry season.
-
华贵栉孔扇贝(Chlamys nobilis)属于热带海水贝类,具有肉质鲜美、营养价值高等优点,自20世纪70年代起,有关其人工育苗和规模化养殖迅速在南方各沿海省份展开,并取得了较好的经济效益,但由于长期养殖,华贵栉孔扇贝也面临种质质量退化的问题,严重影响了该产业的健康发展。目前相关机构已经对其开展了选育研究,并取得了初步成效。数量遗传学作为一种新型研究遗传育种的学科,在水产动物遗传育种中展现了很大的优越性,但将此方法运用于华贵栉孔扇贝良种选育的相关研究尚不多见。该研究在此方向作了初步探索,解决华贵栉孔扇贝养殖产业健康发展的关键所在是建立有效的良种选育方法[3-4]。
三亚崖州湾海区是海南省华贵栉孔扇贝主养区之一,已有5年养殖经历。笔者通过对该海区养殖的华贵栉孔扇贝数量性状的测量和分析,采用多元分析和回归方法,建立形态性状与质量性状间的多元线性回归方程[5],以期为华贵栉孔扇贝通过数量性状辅助选择育种提供相关理论依据。
1. 材料与方法
1.1 材料
受测的华贵栉孔扇贝来自于三亚崖州湾养殖海区,由三亚意源养殖有限公司提供。
1.2 取样和测量方法
2010年3月30日一次性随机选取各年龄阶段526个扇贝标本测量其数量性状。测量前清除其表面附着物。5个形态性状(壳高、壳长、绞合线长、壳厚和贝柱直径)用数显游标卡尺测量(精度为0.02 mm),6个质量性状(总质量、壳质量、贝柱鲜质量、鲜肉质量、干肉质量和贝柱干质量)用电子天平测量(精度为0.01 g)。鲜肉质量指活体去壳后含水分的质量,干肉质量指将鲜肉放入烘箱去除水分后的质量。
1.3 数据分析方法
测量的数据使用SPSS 13软件进行数据分析,对数据采用t检验,用多元回归分析法,分析5个测量指标[壳高(x1)、壳长(x2)、绞合线长(x3)、壳厚(x4)和总质量(x5)]对扇贝3个经济性状[鲜肉质量(y1)、肉柱鲜质量(y2)和肉柱干质量(y3)]的影响,同时作通径分析。
2. 结果与分析
2.1 各性状的参数分析
华贵栉孔扇贝壳高、壳长、绞合线长、壳厚、贝柱直径、总质量、壳质量、鲜肉质量、贝柱鲜质量、干肉质量和贝柱干质量等11个数量性状的各种参数见表 1。其中不同性状的参数单位有所不同,不能进行性状间比较。变异系数的单位相同,因而可以进行性状间比较。性状的变异系数越大,表明品种相应指标可供选择的范围越大,选择的潜力就越大。11个性状的变异系数由大到小依次为贝柱干质量>干肉质量>贝柱鲜质量>鲜肉质量>总质量>壳质量>贝柱直径>壳厚>绞合线长>壳长>壳高。因此,在华贵栉孔扇贝高产品种培育中对这些性状进行选择时,应首先考虑贝柱干质量,其次是干肉质量、贝柱鲜质量、鲜肉质量、总质量、壳质量、贝柱直径、壳厚、绞合线长和壳长,最后是壳宽。
表 1 各性状的表型统计量(n=526)Table 1. Phenotypic statistics of various quantitative traits参数parameter 壳高/mm SH 壳长/mm SL 绞合线长/mm HL 壳厚/mm ST 贝柱直径/mm CP 总质量/g TW 壳质量/g WWAM 鲜肉质量/g FW 贝柱鲜质量/g CFW 干肉质量/g DFW 贝柱干质量/g DWAM max 76.00 72.30 42.00 26.30 23.10 54.01 29.93 24.74 5.32 3.66 1.30 min 44.10 41.00 21.90 13.10 8.90 10.23 7.28 1.33 0.55 0.40 0.07 mean 60.52 55.57 32.58 18.97 15.83 27.86 17.07 10.79 2.40 1.46 0.53 R 31.90 31.30 20.10 13.20 14.20 43.78 22.65 23.41 4.77 3.26 1.23 SD 5.52 5.44 3.32 2.04 2.46 7.08 4.17 3.29 0.87 0.57 0.22 SE 0.24 0.24 0.14 0.09 0.11 0.31 0.18 0.14 0.04 0.02 0.01 CV 9.12% 9.80% 10.19% 10.77% 15.56% 25.43% 24.42% 30.51% 36.31% 38.99% 42.19% 2.2 5个数量性状的回归分析及通径分析
2.2.1 鲜肉质量
分析5个测量指标[壳高(x1)、壳长(x2)、绞合线长(x3)、壳厚(x4)和总质量(x5)]对扇贝鲜肉质量(y)的影响(表 2)。总质量与鲜肉质量为极显著的正相关(相关系数为0.839 4,P < 0.01),壳厚与贝柱鲜质量为显著的负相关(相关系数为-0.110 7,0.01 < P < 0.05)。y=1.295-0.033 15x1-0.091 6x4+0.474 9x5,决定系数R2=0.878 563,剩余标准差SSE=1.150 2。
表 2 5个数量性状对鲜肉质量的逐步回归分析Table 2. Stepwise regression analysis between 5 quantitative traits and shell-free weight变量variable 回归系数regression coefficient 标准回归系数standard regression coefficient 偏相关partial correlation t P x1 -0.033 1 -0.055 5 -0.092 8 2.126 9 0.033 9 x4 -0.091 6 -0.056 8 -0.107 4 2.464 6 0.014 0 x5 0.474 9 1.022 5 0.839 4 35.246 5 0.000 0 通径系数反映自变量对依变量的直接影响,总质量对鲜肉质量的影响最为明显,直接系数为1.024 9(表 3)。x5与鲜肉质量的相关系数最大,而x1和x4与鲜肉质量呈负相关,x5与鲜肉质量呈正相关。
表 3 5个数量性状对鲜肉质量的通径分析Table 3. Path analysis between 5 quantitative traits and shell-free weight变量variable 直接系数direct factor 通过x1 by x1 通过x4 by x4 通过x5 by x5 总影响total impact x1 -0.055 5 - -0.037 8 0.822 6 0.729 3 x4 -0.056 8 -0.036 9 - 0.757 1 0.663 4 x5 1.024 9 -0.044 7 -0.042 1 - 0.935 7 2.2.2 贝柱鲜质量
分析5个测量指标[壳高(x1)、壳长(x2)、绞合线长(x3)、壳厚(x4)和总质量(x5)]对扇贝贝柱鲜质量(y)的影响(表 4),总质量与贝柱鲜质量呈极显著的正相关(相关系数为0.622 9,P < 0.01),绞合线长与贝柱鲜质量呈极显著的负相关(相关系数为-0.196 6,P < 0.01)。y=-0.168 8+0.027 34x2-0.046 04x3+0.091 4x5,决定系数R2=0.617 847,剩余标准差SSE=0.539 5。
表 4 5个数量性状对贝柱鲜质量的逐步回归分析Table 4. Stepwise regression analysis between 5 quantitative traits and adductor muscle weight变量variable 回归系数regression coefficient 标准回归系数standard regression coefficient 偏相关partial correlation t P x2 0.027 3 0.171 0 0.154 5 3.569 8 0.000 4 x3 -0.046 0 -0.175 7 -0.196 6 4.576 6 0.000 0 x5 0.091 4 0.744 1 0.629 9 18.513 1 0.000 0 从表 5中可以看出,通径系数反映自变量对依变量的直接影响。结果显示,总质量对贝柱鲜质量的影响最为明显,直接系数为0.744 1;x2和x5与贝柱鲜质量呈正相关,x3与贝柱鲜质量呈负相关; x5的总体影响最大(0.775 4),x2其次(0.596 0)。
表 5 5个数量性状对贝柱鲜质量的通径分析Table 5. Path analysis between 5 quantitative traits and adductor muscle weight变量variable 直接系数direct factor 通过x1 by x1 通过x4 by x4 通过x5 by x5 总影响total impact x2 0.171 0 - -0.124 4 0.549 5 0.596 0 x3 -0.175 7 0.121 1 - 0.402 1 0.347 5 x5 0.744 1 0.126 3 -0.094 9 - 0.775 4 2.2.3 贝柱干质量
分析5个测量指标[壳高(x1)、壳长(x2)、绞合线长(x3)、壳厚(x4)和总质量(x5)]对扇贝贝柱干质量(y)的影响(表 6)。总质量与贝柱干质量为极显著的正相关(相关系数为0.743 0,P < 0.01),壳长与贝柱干质量为显著的负相关(相关系数为-0.092 2,P < 0.05),绞合线长与贝柱干质量为极显著的负相关(相关系数为-0.141 1,P < 0.01)。y=0.228 7-0.005 0x2-0.007 8x3+0.029 9x5,决定系数R2=0.664 705,剩余标准差SSE=0.129 1。
表 6 5个数量性状对贝柱干质量的逐步回归分析Table 6. Stepwise regression analysis between 5 quantitative traits and adductor muscle dry weight变量variable 回归系数regression coefficient 标准回归系数standard regression coefficient 偏相关partial correlation t P x2 -0.005 0 -0.123 7 -0.119 9 2.756 7 0.006 0 x3 -0.007 8 -0.117 0 -0.141 1 3.253 9 0.001 2 x5 0.029 9 0.954 0 0.743 0 25.341 1 0.000 0 从表 7可以看出,通径系数反映自变量对依变量的直接影响。结果显示,总质量对贝柱鲜质量的影响最为明显,直接系数为0.954 0。
表 7 5个数量性状对贝柱干质量的通径分析Table 7. Path analysis between 5 quantitative traits and adductor muscle dry weight变量variable 直接系数direct factor 通过x1 by x1 通过x4 by x4 通过x5 by x5 总影响total impact x2 -0.123 7 - -0.082 9 0.704 5 0.497 9 x3 -0.117 0 -0.087 6 - 0.515 5 0.310 9 x5 0.954 0 -0.091 3 -0.063 2 - 0.799 4 x5与贝柱干质量呈正相关,x2和x3与贝柱干质量呈负相关,x5对贝柱干质量产生的影响最为明显。
3. 讨论
3.1 通径分析的运用
性状间的相关系数是进行多元分析的基础,但相关系数是两变量间关系的综合体现,其既包括变量间的直接关系,又包括通过其他变量影响的间接关系。单纯采用性状间的相关系数不能正确表述两性状间的真实关系。通径分析可用于分析多个自变量与变量之间的线性关系,是回归分析的拓展,不仅可以处理较为复杂的变量关系,而且能把性状间相关性分成直接影响和间接影响[6-8]。该研究中华贵栉孔扇贝壳高、壳长、绞合线长、壳厚和总质量与扇贝的鲜肉质量、贝柱鲜质量及贝柱干质量的相关系数均极显著,建立回归方程时将其余几个对经济性状相关系数影响不显著的予以剔除以建立理想的回归方程。
3.2 数量性状的选育
衡量华贵栉孔扇贝生长状况的数量性状一般包括长度(高度)和质量,该研究选取长度和质量2个性状来探讨华贵栉孔扇贝数量性状间的相关性。以相关形态性状和总质量为自变量,对3个主要经济性状的回归相关性极显著。把简单易测的性状作为自变量是符合期望的[9-11]。因此,在华贵栉孔扇贝的遗传育种中把壳长和总质量作为重要生长指标即可。通径分析在水产动物的遗传育种中运用非常广泛[12-16],如王爱民等[17]对马氏珠母贝(Pinctada martensii)不同地理种群内自繁和种群间杂交子一代形态性状参数进行了相关分析,郑怀平等[18]对近江牡蛎(Ostrea rivularis gould)养殖群体数量性状间的相关性进行了通径分析,喻子牛等[19]对香港巨牡蛎(Crassostrea hongkongensis)雌雄群体数量性状进行了通径分析。
3.3 选育对加工产业的意义
选育优质的良种对华贵栉孔扇贝加工产业具有相当重要的意义。加工企业需用扇贝的贝柱来加工产品,因而对贝柱的质量格外关注。该研究中对扇贝贝柱鲜质量和贝柱干质量均作了相关分析[20]与通径分析,研究结果显示,总质量对于3个经济性状的正面影响最大。因此,在高产选育时应选择总质量大的品种来进行选育,使获得的优良品种能够产生大的贝柱。
-
![]()
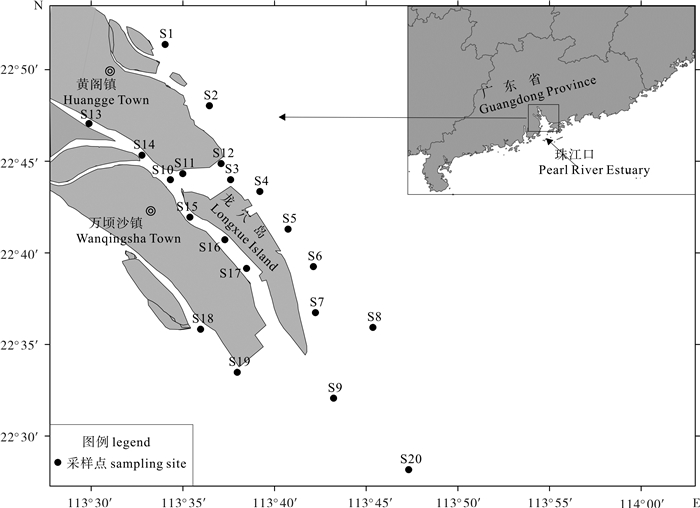
图 1 珠江口南沙海域浮游植物调查示意图
Figure 1. Sampling sites of phytoplankton in the Nansha sea area of Pearl River Estuary
![]()
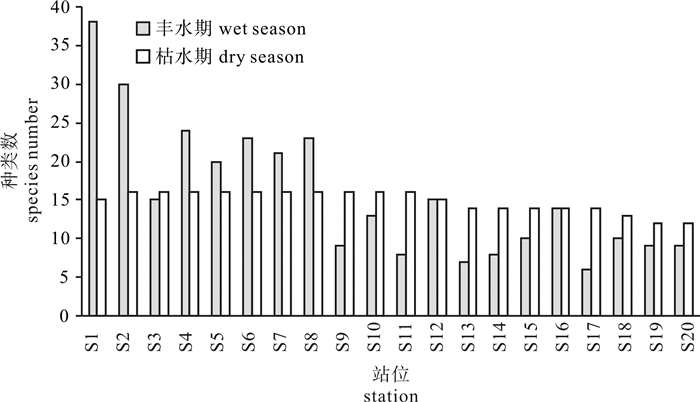
图 2 珠江口南沙海域各站位浮游植物种类数
Figure 2. Number of phytoplankton species of each station in the Nansha sea area of Pearl River Estuary
![]()
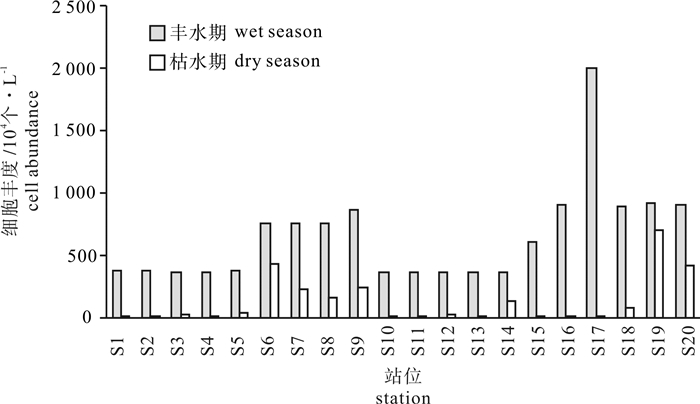
图 3 珠江口南沙海域各站位浮游植物细胞丰度
Figure 3. Abundance of phytoplankton cells of each station in the Nansha sea area of Pearl River Estuary
![]()
图 5 丰水期(a)、枯水期(b)珠江口南沙海域浮游植物物种-环境因子典范对应分析排序图
Figure 5. Ordination diagram of CCA based on phytoplankton species and environmental factors in the Nansha sea area of Pearl River Estuary
表 1 浮游植物多样性指标的评判阈值
Table 1 Threshold value evaluation for diversity indices of phytoplankton
多样性指标
diversity index很低
very low
(Ⅰ)低
low
(Ⅱ)一般
normal
(Ⅲ)高
high
(Ⅳ)很高
very high
(Ⅴ)种类数species number <15 15~24 25~34 35~44 >45 多样性指数(H′)Shannon-Wiener diversity index <1.5 1.5~2.49 2.5~3.49 3.5~4.5 >4.5 均匀度指数(J′)Pielous evenness index <0.68 0.68~0.73 0.74~0.79 0.80~0.88 >0.88  下载: 导出CSV
下载: 导出CSV
表 2 珠江口南沙海域浮游植物物种组成
Table 2 Species composition of phytoplankton in the Nansha sea area of Pearl River Estuary
门类
phylum总种类数
total species number丰水期(5月)
wet season (May)枯水期(11月)
dry season (November)种类
species比例/%
percentage种类
species比例/%
percentage种类
species比例/%
percentage硅藻Bacillariophyta 39 51.32 39 57.35 33 55.00 甲藻Pyrrophyta 15 19.74 7 10.29 14 23.33 绿藻Chlorophyta 15 19.74 15 22.06 8 13.33 蓝藻Cyanophyta 2 2.63 2 2.94 2 3.33 裸藻Euglenophyta 5 6.58 5 7.35 3 5.00 总计total 76 - 68 - 60 -
下载: 导出CSV
表 3 珠江口南沙海域浮游植物多样性评价
Table 3 Diversity evaluation for phytoplankton in the Nansha sea area of Pearl River Estuary
季节
season很低
very low
(Ⅰ)低
low
(Ⅱ)一般
normal
(Ⅲ)高
high
(Ⅳ)很高
very high
(Ⅴ)总评价
general evaluation丰水期wet season 种类数(S)species number 87.50% 12.50% Ⅰ 多样性指数(H′)Shannon-Wiener diversity index 62.50% 37.50% Ⅰ 均匀度指数(J′)Pielous evenness index 100% Ⅰ 枯水期dry season 种类数(S) species number 40% 60% Ⅱ 多样性指数(H′)Shannon-Wiener diversity index 45% 45% 10% Ⅱ 均匀度指数(J′)Pielous evenness index 100% Ⅰ
下载: 导出CSV
表 4 珠江口南沙海域浮游植物细胞丰度与环境因子的相关性
Table 4 Correlation between cell density of phytoplankton and environmental factors in the Nansha sea area of Pearl River Estuary
季节
season水温
temperature盐度
salinity溶解氧
DO酸碱度
pH丰水期wet season -0.670* -0.209 0.208 0.378 枯水期dry season 0.491 0.597** -0.233 0.133 注:*.P < 0.05;* *.P < 0.01
下载: 导出CSV
-
[1] CHARLSON R J, LOVELOCK J E, ANDERAE M O, et al.Oceanic phytoplankton, atmospheric sulphur, cloud albedo and climate[J].Nature, 1987, 326:655-661. doi: 10.1038/326655a0
[2] LEPISTO L, HOLOPAINEN L L, VUORISTO H.Type-specific and indicator taxa of phytoplankton as a quality criterion for assessing the ecological status of Finnish boreal lakes[J].Limnologica, 2004, 34(3):236-248. doi: 10.1016/S0075-9511(04)80048-3
[3] 柯志新, 黄良民, 谭烨辉, 等.2007年夏季南海北部浮游植物的物种组成及丰度分布[J].热带海洋学报, 2011, 30(1):131-143. doi: 10.11978/j.issn.1009-5470.2011.01.131 [4] 戴明, 巩秀玉, 刘华雪, 等.2013年春季南沙海域不同水团中网采浮游植物群落特征[J].南方水产科学, 2015, 11(5):38-46. http://kns.cnki.net/KCMS/detail/detail.aspx?dbcode=CJFQ&dbname=CJFDLAST2015&filename=NFSC201505006&v=MDA1NDRiYkc0SDlUTXFvOUZZb1I4ZVgxTHV4WVM3RGgxVDNxVHJXTTFGckNVUkwyZVplZHVGeXprVjcvSkt5dlk= [5] 粟丽, 黄梓荣, 陈作志.水东湾春、秋季浮游植物群落结构特征[J].南方水产科学, 2015, 11(4):27-33. http://kns.cnki.net/KCMS/detail/detail.aspx?dbcode=CJFQ&dbname=CJFDLAST2015&filename=NFSC201504005&v=MTg1ODYyZVplZHVGeXprVjd6Tkt5dlliYkc0SDlUTXE0OUZZWVI4ZVgxTHV4WVM3RGgxVDNxVHJXTTFGckNVUkw= [6] 姜胜, 顾继光, 冯佳和, 等.广州海域环境质量评价[J].应用生态学报, 2006, 17(5):894-898. http://www.cnki.com.cn/Article/CJFDTOTAL-SCKX198804002.htm [7] 周燕遐.珠江口及邻近海域水质状况分析[J].海洋通报, 1994, 13(3):24-30. http://www.oalib.com/references/19444297 [8] 戴明, 李纯厚, 贾晓平, 等.珠江口近海浮游植物生态特征研究[J].应用生态学报, 2004, 15(8):1389-1394. http://www.oalib.com/paper/1479515 [9] 戴娟, 王超, 赖子尼, 等.2006年夏季珠江口5大口门网采浮游植物群落的调查[J].淡水渔业, 2007, 37(3):63-64. http://www.cqvip.com/qk/93836X/200703/24654293.html [10] 冯洁娉, 姜胜, 乔永民, 等.珠江口广州段浮游植物群落特征研究[J].生态科学, 2012, 31(3):285-287. http://www.cqvip.com/QK/97324X/201203/43134014.html [11] 刘凯然. 珠江口浮游植物生物多样性变化趋势[D]. 大连: 大连海事大学, 2008: 2-35. http://kns.cnki.net/KCMS/detail/detail.aspx?dbcode=CMFD&dbname=CMFD2009&filename=2009074966.nh&v=MjYyNDIxTHV4WVM3RGgxVDNxVHJXTTFGckNVUkwyZVplZHVGeXprVkx6TlYxMjdGN08vR3RqS3FaRWJQSVI4ZVg= [12] 李开枝, 黄良明, 张建林, 等.珠江河口咸潮期间浮游植物的群落特征[J].热带海洋学报, 2010, 29(1):62-68. doi: 10.11978/j.issn.1009-5470.2010.01.062 [13] 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB/T 12763. 6—2007. 海洋调查规范(第6部分-海洋生物调查)[S]. 北京: 中国标准出版社, 2007: 34-38. http://dbpub.cnki.net/grid2008/dbpub/detail.aspx?dbcode=SCSF&dbname=SCSF&filename=SCSF00001255 [14] 国家海洋监测中心. GB17378. 3—2007海洋监测规范第3部分: 样品采集、贮存与运输[S]. 北京: 中国标准出版社, 2008: 4-8. [15] SHANNON C E, WEAVER W. The mathematical theory of communication[M].Illinois:University of Illinoi Press, 1949:1-117.
[16] PIELOU E C.Species-diversity and pattern-diversity in the study of ecological succesion[J].J Theor Biol, 1966, 10(2):370-383. http://www.oalib.com/references/14451572
[17] 孙军, 刘东艳, 魏皓, 等.琉球群岛邻近海域浮游植物多样性的模糊综合评判[J].海洋与湖沼, 2001, 32(4):946-955. https://www.researchgate.net/publication/263747753... [18] 张峰, 上官铁梁.山西南方红豆杉(Taxus mairei)森林群落的生态优势度分析[J].山西大学学报(自然科学版), 1988, 11(3):82-87. http://kns.cnki.net/kns/detail/detail.aspx?QueryID=39&CurRec=1&recid=&FileName=SXDR198803016&DbName=CJFD8589&DbCode=CJFQ&yx=&pr=&URLID= [19] 曾江宁, 曾淦宁, 黄卫民, 等.赤潮影响因素研究进展[J].东海海洋, 2004, 22:40-47. doi: 10.3969/j.issn.1001-909X.2004.02.007 [20] 王迪, 陈丕茂, 逯晶晶, 等.钦州湾浮游植物周年生态特征[J].应用生态学报, 2013, 24(6):1686-1692. http://www.oalib.com/paper/4376280 [21] 吴建新, 李强, 张晴.吕泗大洋港浮游植物群落与环境因子关系[J].生态学杂志, 2013, 32(2):396-400. http://edu.wanfangdata.com.cn/Periodical/Detail/stxzz201302025 [22] 庄军莲, 许铭本, 张荣灿, 等.广西防城港周年浮游植物生态特征[J].应用生态学报, 2011, 22(5):1309-1315. http://www.cjae.net/CN/abstract/abstract12386.shtml [23] 廖秀丽, 陈丕茂, 马胜伟, 等.大亚湾杨梅坑海域投礁前后浮游植物群落结构及其与环境因子的关系[J].南方水产科学, 2013, 9(5):109-119. http://kns.cnki.net/KCMS/detail/detail.aspx?dbcode=CJFQ&dbname=CJFDHIS2&filename=NFSC201305023&v=MzI0MTVIWjRSOGVYMUx1eFlTN0RoMVQzcVRyV00xRnJDVVJMMmVaZWR1Rnl6a1ZMdk5LeXZZYmJHNEg5TE1xbzk= [24] 张才学, 龚玉艳, 孙省利.湛江港湾潜在赤潮生物的时空分布及其影响因素[J].生态学杂志, 2012, 31(7):1763-1770. http://www.wenkuxiazai.com/doc/5ddeec452b160b4e767fcf8f.html [25] 郭凯旋, 张瑜斌, 章洁香, 等.雷州半岛近海夏季浮游植物和浮游细菌生物量的分布及影响因[J].生态学杂志, 2012, 31(1):8-15. http://edu.wanfangdata.com.cn/Periodical/Detail/stxzz201201002 -
期刊类型引用(10)
1. 刘永新,邵长伟,郑先虎. 中国淡水重要养殖鱼类抗病和抗逆性状育种研究进展. 水产学报. 2025(02): 4-21 .  百度学术
百度学术
2. 张元,刘志刚,王春德,姚高友,张柯馨,展建强,陈麟广,吕文刚,钟茂程,刘付少梅. 扇贝新品系“紫海墨”F_4的生长特征和生产性能分析. 南方水产科学. 2022(01): 68-76 . 本站查看
3. 申奔龙,薛宝宝,孟德龙,沈和定. 缢蛏早期耐高温家系建立及抗氧化能力测定. 浙江农业学报. 2022(02): 266-274 . 百度学术
4. 朱佳杰,敖秋桅,谭芸,罗永巨,蒋和生. 吉富罗非鱼组蛋白酶B基因克隆及无乳链球菌感染后的表达分析. 南方农业学报. 2022(08): 2294-2301 . 百度学术
5. 周欣,高风英,卢迈新. 鱼类抗病育种研究进展. 大连海洋大学学报. 2021(03): 510-523 . 百度学术
6. 严楚,彭凯,张志妮. 罗非鱼链球菌综合防控技术研究进展. 广东畜牧兽医科技. 2021(06): 13-16 . 百度学术
7. 敖秋桅,朱佳杰. 3个罗非鱼种群对4种病原菌的抗病力差异比较. 南方农业学报. 2020(03): 695-701 . 百度学术
8. 付璐璐,王峥,王磊,李学军. 近10年水产动物抗逆育种研究进展. 江苏农业科学. 2020(16): 52-58 . 百度学术
9. 方佳峰,李琪. 长牡蛎壳橙品系幼虫和稚贝的生长性状遗传参数评估. 中国海洋大学学报(自然科学版). 2020(11): 38-44 . 百度学术
10. 郑雄,赵立朝,吴志强,张洁,黄亮亮,汪开成,张曼. 广西右江流域野生罗非鱼种群及其杂合性研究. 南方农业学报. 2019(12): 2797-2805 . 百度学术
其他类型引用(8)
计量
- 文章访问数: 3462
- HTML全文浏览量: 1095
- PDF下载量: 753
- 被引次数: 18
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
