
Forecasting fishing ground of calamary in the northern South China Sea according to habitat suitability index
-
摘要:
根据2009年~2014年南海捕捞信息网络获取的南海北部200 m等深线以内近海底拖网渔业数据,结合环境遥感数据,构建南海北部枪乌贼类(Uroteuthis sp.)栖息地指数(habitat suitability index, HSI)模型。选取海表水温(sea surface temperature, SST)、海面高度(sea surface height, SSH)和叶绿素a浓度(chlorophyll-a concentration, CHL)等环境因子,采用一元非线性回归建模,并分月份采用最小二乘法确定各环境因子的权重系数,从而建立南海北部枪乌贼类的栖息地指数综合模型。构建的模型能够有效解释渔场分布与环境要素之间的关系, 并可从模型中推断各环境因子的最适范围和变化趋势;单位捕捞努力量渔获量(catch per unite effect, CPUE)高的渔场主要分布于HSI大于0.5的海域,其他HSI高的海域可能为潜在渔场。对不同因子权重系数的研究发现,叶绿素a浓度是影响渔场分布的重要因素。对模型的验证结果表明,其准确率达到75%以上。该研究表明,基于不同权重系数的HSI模型能够较好地预测南海北部枪乌贼渔场。
Abstract:Based on the data of bottom trawl fishery within shelf area at depth of less than 200 m by a fishery information network from 2009 to 2014, and environmental data obtained from remote sensing, we established the habitat suitability index (HSI) models for calamary (Uroteuthis sp.) in the northern South China Sea. The sea surface temperature, sea surface height and concentration of chlorophyll-a were determined by nonlinear regression modeling. The integrated HSI models for each month, which were established using least square method for assigning weight coefficients to the environmental factors, could explain the relationship between distribution of fishing grounds and environmental factors effectively, and deduce the most suitable range and variation tendency of the environmental factors. High catches per unite effect (CPUE) values of the trawl fishery were mainly in the areas where the HSIs were higher than 0.5, and the other high HSI areas might be potential fishing grounds. Weight coefficients for the environmental factors in the integrated HSI models suggest that chlorophyll-a concentration is the most important factor influencing the distribution of fishing grounds. Verification of the HSI models shows an accuracy of over 75%. The study indicates that the integrated HSI models based on different weight coefficients for environmental factors can better predict fishing grounds of calamary in the northern South China Sea.
-
枪乌贼属(Uroteuthis)隶属于头足纲、枪形目、枪乌贼科,广泛分布于热带亚热带大陆架海域[1],在南海北部是重要的捕捞对象,近年来年产量均在15×104 t左右,并呈逐年上升的趋势[2]。已有的研究认为枪乌贼的分布跟海表温度(sea surface temperature, SST)、盐度、叶绿素a浓度(chlorophyll-a concentration, CHL)、海面高度(sea surface height, SSH)等环境因子密切相关[3-4]。陈新军等[5]研究了表温对北太平洋柔鱼(Ommastrephes bartramii)渔场形成的影响,孙典荣等[6]对北部湾剑尖枪乌贼(U.edulis)的资源变动特征进行了研究,WANG等[7]对东海南部的剑尖枪乌贼对环境的响应进行了分析。栖息地模型可以用来模拟生物对环境因子的反应,广泛应用于渔情预报的研究[8-9]。已有的研究大多没有分别考虑不同环境因子对渔场形成的影响[5, 7, 10]。不同因子对渔场形成的影响程度并不一样。为此,该研究基于栖息地指数(habitat suitability index, HSI)模型,采用不同环境因子,尝试建立基于HSI的南海北部枪乌贼渔情预报模型,对南海北部枪乌贼的渔场分布状况和中心渔场进行分析研究,为南海北部枪乌贼资源的合理开发利用提供科学依据。
1. 材料与方法
1.1 数据来源
1.1.1 渔业数据
渔业数据来自南海渔船信息动态监测网络,时间为2009年~2014年。在其中选取200 m等深线以内陆架区底拖网渔业枪乌贼的渔获统计数据(图 1),由于生活习性相似[4],故而将中国枪乌贼(U.chinensis)和剑尖枪乌贼合并为枪乌贼类进行统计处理。由于南海区伏季休渔为每年的5月16日~7月31日,该研究统计每年1月~5月和8月~12月共10个月的生产数据,为方便进行数据标准化,结合不同类型渔船数量与数据特点,仅选择底拖网渔船数据,总作业网次25 128网,渔船总数92艘。
1.1.2 环境数据
环境因子选取SST、CHL、SSH等,均采用遥感数据,来自美国国家海洋和大气管理局(National Oceanic and Atmospheric Administration, NOAA),数据分辨率为0.25°×0.25°。
1.2 处理方法
1.2.1 渔场中心和单位捕捞努力量渔获量(catch per unite effect, CPUE)
按经纬度0.25°×0.25°统计底拖网生产数据,按月计算其CPUE。
$ {\rm{CPUE}} = Y/M $
(1) 式中Y为该月的渔获量(kg),M为该月在此区域作业的渔船功率数(kW)。
对全部数据按月随机选取70%进行建模计算,剩余30%用于模型验证。
1.2.2 模型计算
利用CPUE分别与SST、SSH、CHL来建立相应的适应性指数(suitability in dex, SI)模型。该研究假定最高的CPUE为枪乌贼资源分布最多的海域,认定其SI为1;CPUE为0时,则认定是枪乌贼资源分布较少的海域,认定其SI为0。SI实际值的计算公式为:
$ {\rm{S}}{{\rm{I}}_i} = \frac{{{\rm{CPUE}}}}{{{\rm{CPU}}{{\rm{E}}_{\max }}}} $
(2) 式中SIi为i月得到的适应性指数;CPUEmax为i月的最大单位捕捞努力量渔获量。
利用一元非线性回归分别建立SST、SSH、CHL与SI之间的关系模型[11],计算公式为:
$ {\rm{S}}{{\rm{I}}_f} = {{\rm{e}}^{{\rm{A}} \times {{{\rm{(B + }}\mathit{X}{\rm{)}}}^2}}} $
(3) 式中f为各环境因子,A、B均为模型系数,X为各因子的值,利用DPS 12.0软件求解。
将建立的模型代入公式计算HSI理论值,计算公式为:
$ {\rm{HSI = a}} \times {\rm{S}}{{\rm{I}}_{{\rm{SST}}}} + {\rm{b}} \times {\rm{S}}{{\rm{I}}_{{\rm{SSH}}}} + {\rm{c}} \times {\rm{S}}{{\rm{I}}_{{\rm{CHL}}}} $
(4) 式中a、b、c分别代表各模型的权重系数,均大于等于0,小于等于1,且a+b+c=1。采用最小二乘法使得模型计算理论值HSI与实际值SI最接近,分月份利用Excel 2010的规划求解,确定a、b、c值,从而确定HSI模型。
1.2.3 模型精度验证
分别计算剩余30%数据实际值SIi和模型理论值HSI。如实际值与理论值之差绝对值小于等于0.4,则认为模型预报准确,如大于0.4则认为模型预报不准确。并采用2014年渔业资源调查站位中出现枪乌贼类的站点,共18个,数据处理参照上述方法,带入模型,验证模型精度。
2. 结果
2.1 各因子的SI
采用一元非线性回归,分不同月份分别建立各因子与CPUE间的SI,结果见表 1。表中各式,显著性水平均为P < 0.01,可知模型拟合准确。
表 1 各因子适应性指数模型Table 1 SI model for each factor月份month SISST SISSH SICHL 1 SI=e-0.153 8×(20.61-SST)2 SI=e-0.015 69×(108.2-SSH)2 SI=e-2.871×(1.279-CHL)2 2 SI=e-0.038 41×(19.24-SST)2 SI=e-0.086 13×(94.23-SSH)2 SI=e-1.661×(4.428-CHL)2 3 SI=e-0.056 05×(21.86-SST)2 SI=e-0.011 29×(70.11-SSH)2 SI=e-5.356×(1.931-CHL)2 4 SI=e-0.161 1×(23.65-SST)2 SI=e-0.004 74×(80.02-SSH)2 SI=e-0.427×(1.938-CHL)2 5 SI=e-0.934 2×(23.90-SST)2 SI=e-0.051 68×(93.56-SSH)2 SI=e-1.217×(1.015-CHL)2 8 SI=e-1.089 1×(27.80-SST)2 SI=e-0.062 05×(98.80-SSH)2 SI=e-8.530×(4.026-CHL)2 9 SI=e-0.280 8×(28.97-SST)2 SI=e-0.069 79×(99.49-SSH)2 SI=e-4.187×(1.461-CHL)2 10 SI=e-0.238 8×(27.28-SST)2 SI=e-0.004 93×(99.82-SSH)2 SI=e-6.694×(0.510-CHL)2 11 SI=e-0.104 5×(24.66-SST)2 SI=e-0.004 40×(99.01-SSH)2 SI=e-1.333×(0.956-CHL)2 12 SI=e-0.171 6×(23.15-SST)2 SI=e-0.004 20×(91.12-SSH)2 SI=e-3.807×(2.362-CHL)2 2.2 各月份各因子最适值
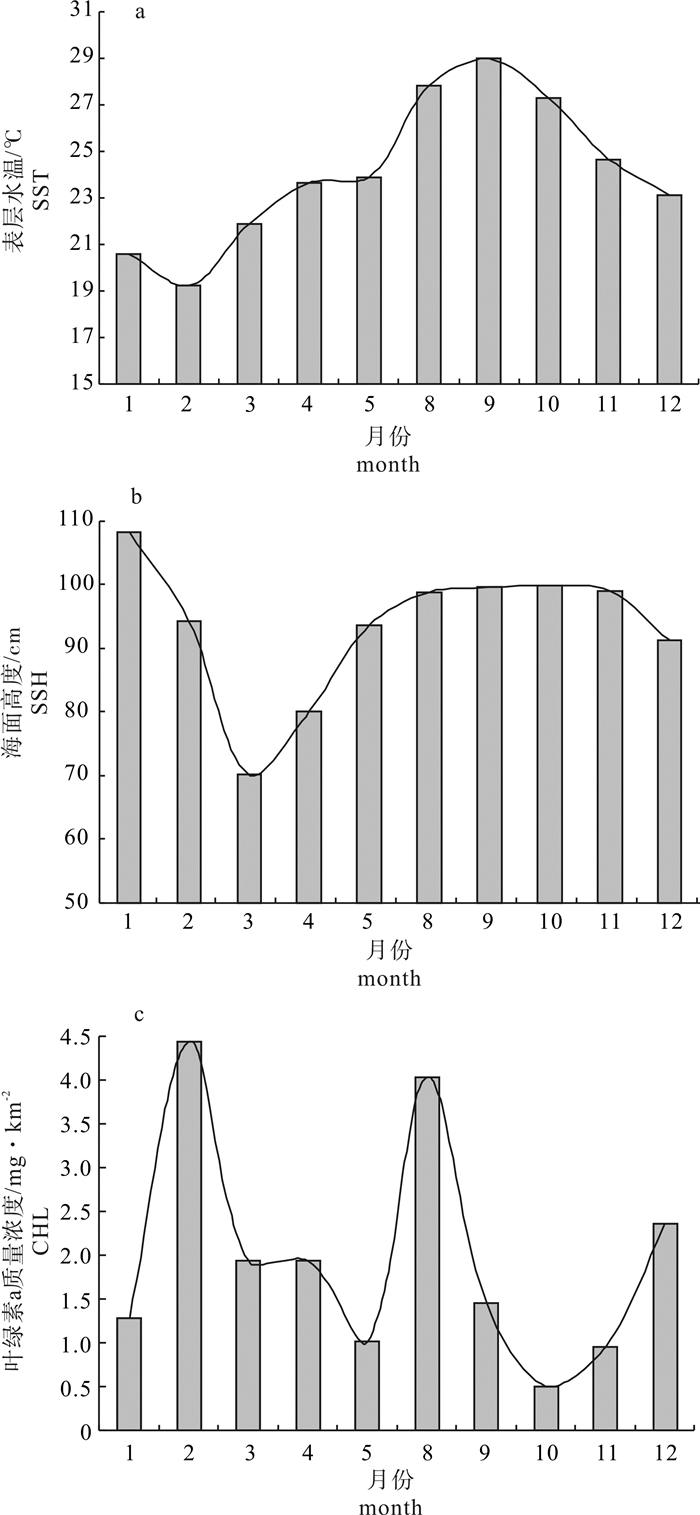
各月份各因子的最适值在SI接近1时取得(表 1),故公式(3)中的B值即为各月份各因子的最适值,结果见图 2。
SST的最适值的变化呈现先增大后减小的趋势,在9月达到最高(28.97 ℃),2月最低(19.24 ℃),基本呈正态型分布(图 2-a)。SSH的最适值呈先减小后增大的趋势,3月最低(70.11 cm),1月最高(108.2 cm),呈偏正态分布(图 2-b)。CHL的最适值全年有2个波峰,分别出现在2月和8月,波谷出现在5月和10月,最大值和最小值分别为4.428 mg · km-2和0.51 mg · km-2,呈明显的双峰分布(图 2-c)。
2.3 模型系数的确定
根据公式(4),采用最小二乘法使得模型计算HSI数值与实际SI最接近,从而确定各因子权重系数。分月份利用Excel 2010的规划求解,结果见表 2。
表 2 各模型权重系数Table 2 Weight coefficient for each model月份
montha b c 1 0.302 2 0.241 0 0.454 8 2 0.194 1 0.000 0 0.805 9 3 0.269 8 0.482 1 0.248 0 4 0.000 0 0.000 0 0.998 0 5 0.385 4 0.472 1 0.140 5 8 0.415 7 0.368 1 0.214 3 9 0.033 3 0.311 2 0.653 5 10 0.000 8 0.195 4 0.801 8 11 0.000 1 0.000 0 0.997 9 12 0.079 5 0.162 6 0.755 9 平均值 mean 0.168 1 0.223 3 0.607 1 表 2中a、b、c分别为SST、SSH、CHL的权重系数,可看出各因子的系数没有明显的季节变化趋势,但c值平均值超过0.5,所占权重较大,而a与b平均值均未超过0.5,可知渔场形成与SST和SSH的关系不大,而与CHL关系密切。
2.4 模型精度验证
根据已建立的HSI模型,将剩余的30%CPUE数据代入模型中计算相应的理论值HSI和实际值SI,理论值与实际值之差小于等于0.4,则认为模型预报准确,结果见表 3,作业渔区指按照0.25°精度统计的渔区个数,各月份渔区数大致相同。1月~5月、8月~12月模型预报准确率在59%~92%,平均准确率为77%,表明所建立的模型能够有效解释渔场分布与环境要素之间的关系。
表 3 渔场预报结果统计Table 3 Results of forecast月份
trmonth作业渔区个数
number of fishing area预测正确
correct forecast预测不正确
wrong forecast渔区数
number of fishing areas比例/%
proportion渔区数
number of fishing areas比例/%
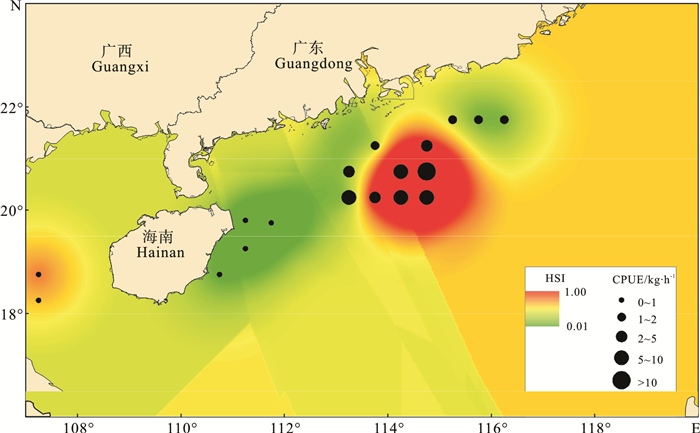
proportion1 23 18 78 5 22 2 22 13 59 9 41 3 24 15 63 9 38 4 25 18 72 7 28 5 24 20 83 4 17 8 24 19 79 5 21 9 27 22 81 5 19 10 26 21 81 5 19 11 25 20 80 5 20 12 26 24 92 2 8 合计total 246 190 77 56 23 为了进一步验证模型的准确性,该研究尝试用2014年春季(3月)渔业资源近岸调查的数据(底拖网)进行进一步检验,结果见图 3。可以看出CPUE值较高的点均出现在预报的高HSI区域。
3. 讨论
该研究采用遥感数据与捕捞数据相结合,构建了南海北部枪乌贼类的渔情预报模型,并采用最小二乘法确定模型中各因子的权重系数,提高了模型的准确性。而各因子的权重系数也能分别反映和解释不同的物理生物过程,可为南海北部枪乌贼渔场分析与资源管理提供理论依据,并对海洋生物变化过程进行相应解释。
南海北部海底地形复杂,海洋环境因子的变化有别于其他海区,因此渔场形成的机制复杂,对渔场判别与渔情预报的研究有更高的要求[2, 12-14]。已有的研究推测头足类渔场的形成和资源量变动与SST、SSH、CHL等海洋因子密切相关,但研究方向主要集中在其生物学特征[15-18]和资源量的时空变化上[6, 19-20],而其渔场与海洋环境因子关系的研究还较为少见。根据该研究建立的模型可推断出各月份SST、SSH和CHL的最适值,即为公式(3)中的B值。该研究的结果显示,最适SST呈明显的季节性变化;SSH在春季明显偏低,这可能受冬季季风结束而西南季风尚未盛行的影响;最适CHL的月份变化没有明显规律,但平均值均较高,明显高于海区的平均值。在实际渔情预报中,若环境数据获取不完全,可以通过一两个因子进行渔场位置的大致判断。
该研究分月份对不同SI模型赋予不同权重系数,探讨不同环境因子对枪乌贼分布的影响。从研究结果来看,SST的权重系数在全年中的占比均未超过0.5,而CHL的权重均比较高,与其他大洋性头足类分布有所差异[1, 5],这可能是由于南海的物理环境比较稳定,但海域生物生产力较低,头足类的分布更容易受到饵料生物分布的影响。
进一步探讨各环境因子权重系数的变化发现,CHL的权重系数均值超过0.5,表明CHL对HSI的影响较大,超过0.5的月份有2月、4月、9月、10月、11月和12月。尝试从枪乌贼的生活习性分析,枪乌贼主要摄食小型鱼类、甲壳类和头足类,而主要饵料种类的生长周期在2~3个月,故而CHL对枪乌贼渔场分布的影响要迟滞2~3个月[6]。另外,枪乌贼繁殖高峰在春夏季,此时枪乌贼的摄食强度较低[1],也从另一方面解释了此种现象。此外,从饵料生物的聚集效应来考虑,CHL高的区域有利于浮游动物的繁殖生长,从而会吸引小型鱼类等饵料生物聚集,进而会吸引枪乌贼聚集。这也有一个滞后效应,但会比生长周期短。
枪乌贼类有昼夜垂直移动的现象[21],夜间在表层进行觅食,白天沉入底层。该研究选取的是只在白天作业的底拖网渔船的数据,恰好能充分反映其分布规律和资源状况。但由于选用的是捕捞生产渔船的数据,数据存在一定的偏差,在今后的研究中应当结合多种作业方式和更多的环境因子,更全面地探讨枪乌贼渔场的形成机制和分布规律。
-
表 1 各因子适应性指数模型
Table 1 SI model for each factor
月份month SISST SISSH SICHL 1 SI=e-0.153 8×(20.61-SST)2 SI=e-0.015 69×(108.2-SSH)2 SI=e-2.871×(1.279-CHL)2 2 SI=e-0.038 41×(19.24-SST)2 SI=e-0.086 13×(94.23-SSH)2 SI=e-1.661×(4.428-CHL)2 3 SI=e-0.056 05×(21.86-SST)2 SI=e-0.011 29×(70.11-SSH)2 SI=e-5.356×(1.931-CHL)2 4 SI=e-0.161 1×(23.65-SST)2 SI=e-0.004 74×(80.02-SSH)2 SI=e-0.427×(1.938-CHL)2 5 SI=e-0.934 2×(23.90-SST)2 SI=e-0.051 68×(93.56-SSH)2 SI=e-1.217×(1.015-CHL)2 8 SI=e-1.089 1×(27.80-SST)2 SI=e-0.062 05×(98.80-SSH)2 SI=e-8.530×(4.026-CHL)2 9 SI=e-0.280 8×(28.97-SST)2 SI=e-0.069 79×(99.49-SSH)2 SI=e-4.187×(1.461-CHL)2 10 SI=e-0.238 8×(27.28-SST)2 SI=e-0.004 93×(99.82-SSH)2 SI=e-6.694×(0.510-CHL)2 11 SI=e-0.104 5×(24.66-SST)2 SI=e-0.004 40×(99.01-SSH)2 SI=e-1.333×(0.956-CHL)2 12 SI=e-0.171 6×(23.15-SST)2 SI=e-0.004 20×(91.12-SSH)2 SI=e-3.807×(2.362-CHL)2  下载: 导出CSV
下载: 导出CSV
表 2 各模型权重系数
Table 2 Weight coefficient for each model
月份
montha b c 1 0.302 2 0.241 0 0.454 8 2 0.194 1 0.000 0 0.805 9 3 0.269 8 0.482 1 0.248 0 4 0.000 0 0.000 0 0.998 0 5 0.385 4 0.472 1 0.140 5 8 0.415 7 0.368 1 0.214 3 9 0.033 3 0.311 2 0.653 5 10 0.000 8 0.195 4 0.801 8 11 0.000 1 0.000 0 0.997 9 12 0.079 5 0.162 6 0.755 9 平均值 mean 0.168 1 0.223 3 0.607 1
下载: 导出CSV
表 3 渔场预报结果统计
Table 3 Results of forecast
月份
trmonth作业渔区个数
number of fishing area预测正确
correct forecast预测不正确
wrong forecast渔区数
number of fishing areas比例/%
proportion渔区数
number of fishing areas比例/%
proportion1 23 18 78 5 22 2 22 13 59 9 41 3 24 15 63 9 38 4 25 18 72 7 28 5 24 20 83 4 17 8 24 19 79 5 21 9 27 22 81 5 19 10 26 21 81 5 19 11 25 20 80 5 20 12 26 24 92 2 8 合计total 246 190 77 56 23
下载: 导出CSV
-
[1] 陈新军, 刘必林, 王尧耕.世界头足类[M].北京:海洋出版社, 2009:25-49. [2] 黄梓荣.南海北部陆架区头足类的种类组成和资源密度分布[J].南方水产, 2008, 4(5):1-7. http://www.cqvip.com/QK/60642A/200805/28370238.html [3] ROBIN J P, DENIS V.Squid stock fluctuations and water temperature:temporal analysis of English Channel Loliginidae[J].J Appl Ecol, 1999, 36(1):101-110. doi: 10.1046/j.1365-2664.1999.00384.x
[4] YAN Y R, LI Y Y, YANG S Y, et al. Biological characteristics and spatial-temporal distribution of mitre squid, Uroteuthis chinensis, in the Beibu Gulf, South China Sea [J]. J Shellfish Res, 2013, 32(3):835-844. doi: 10.2983/035.032.0327
[5] 陈新军, 刘必林, 田思泉, 等.利用基于表温因子的栖息地模型预测西北太平洋柔鱼(Ommastrephes bartramii)渔场[J].海洋与湖沼, 2009, 40(6):707-713. doi: 10.11693/hyhz200906006006 [6] 孙典荣, 李渊, 王雪辉, 等.北部湾剑尖枪乌贼生物学特征及资源状况变化的初步研究[J].南方水产科学, 2011, 7(2):8-13. http://kns.cnki.net/kns/detail/detail.aspx?QueryID=0&CurRec=1&recid=&FileName=NFSC201102002&DbName=CJFD2011&DbCode=CJFQ&yx=&pr=&URLID= [7] WANG K Y, CHANG K Y, LIAO C H, et al. Growth strategies of the swordtip squid, Uroteuthis edulis, in response to environmental changes in the southern East China Sea:a cohort analysis [J]. Bull Mar Sci, 2013, 89(3):677-698. doi: 10.5343/bms.2012.1044
[8] BERTIGNAC M, LEHODEY P, HAMPTON J. A spatial population dynamics simulation model of tropical tunas using a habitat index based on environmental parameters[J]. Fish Oceanogr, 1998, 7(3/4):326-334. doi: 10.1046/j.1365-2419.1998.00065.x/references
[9] SONG L, ZHOU Y. Developing an integrated habitat index for bigeye tuna (Thunnus obesus)in the Indian Ocean based on longline fisheries data[J].Fish Res, 2010, 105(2):63-74. doi: 10.1016/j.fishres.2010.03.004
[10] 陈新军, 冯波, 许柳雄.印度洋大眼金枪鱼栖息地指数研究及其比较[J].中国水产科学, 2008, 15(2):269-278. http://kns.cnki.net/KCMS/detail/detail.aspx?dbcode=CJFQ&dbname=CJFD2008&filename=ZSCK200802011&v=MjI4NDBGckNVUkwyZVplZHVGeWptVjczSVB6N0laYkc0SHRuTXJZOUVaWVI4ZVgxTHV4WVM3RGgxVDNxVHJXTTE= [11] 李永孝.农业应用生物统计[M].济南:山东科学技术出版杜, 1989:313-327. [12] 陈作志, 邱永松.南海北部生态系统食物网结构、能量流动及系统特征[J].生态学报, 2010, 30(18):4855-4865. http://industry.wanfangdata.com.cn/dl/Detail/Conference?id=... [13] 郭金富, 陈丕茂.南海头足类资源开发利用研究[J].热带海洋, 2000, 19(4):51-58. http://www.cnki.com.cn/Article/CJFDTOTAL-ZMSK201003044.htm [14] 李建柱, 陈丕茂, 贾晓平, 等.中国南海北部剑尖枪乌贼资源现状及其合理利用对策[J].中国水产科学, 2010, 17(6):1309-1318. http://kns.cnki.net/KCMS/detail/detail.aspx?dbcode=CJFQ&dbname=CJFD2010&filename=ZSCK201006024&v=MDQ0MTdXTTFGckNVUkwyZVplZHVGeWptVjd2TVB6N0laYkc0SDlITXFZOUhZSVI4ZVgxTHV4WVM3RGgxVDNxVHI= [15] MUCHLISIN Z A, ZULKARNAINI B, PURNAWAN S, et al.Morphometric variations of three species of harvested cephalopods found in northern sea of Aceh Province, Indonesia[J].Biodiversitas, 2014, 15:142-146. doi: 10.13057/biodiv
[16] SIDDIQUE M A M, ARSHAD A, NURUL AMIN S M. Length-weight relationships of the tropical cephalopod Uroteuthis chinensis(Gray, 1849) from Sabah, Malaysia[J].Zool Ecol, 2014, 24(3):215-218. doi: 10.1080/21658005.2014.934515
[17] IKEDA Y, KOBAYASHI M.Statolith growth of juvenile oval squid Sepioteuthis lessoniana (Cephalopoda:Loliginidae) with special reference to ambient thermal condition[J].Mar Biol Res, 2010, 6(5):485-495. doi: 10.1080/17451000903334710
[18] JACKSON G, FORSYTHE J, HIXON R, et al.Age, growth, and maturation of Lolliguncula brevis(Cephalopoda:Loliginidae)in the northwestern Gulf of Mexico with a comparison of length-frequency versus statolith age analysis[J].Can J Fish Aquat Sci, 1997, 54(12):2907-2919. doi: 10.1139/f97-192
[19] 俞存根, 虞聪达, 宁平, 等.浙江南部外海头足类种类组成和数量分布[J].海洋渔业, 2009, 31(1):27-33. http://www.cqvip.com/QK/93342X/200901/29693201.html [20] 朱文斌, 薛利建, 卢占晖, 等.东海南部海域头足类群落结构特征及其与环境关系[J].海洋与湖沼, 2014, 45(2):436-442. doi: 10.11693/hyhz20121228001 [21] JACKSON G, MOLTSCHANIWSKYJ N. Spatial and temporal variation in growth rates and maturity in the Indo-Pacific squid Sepioteuthis lessoniana (Cephalopoda:Loliginidae) [J]. Mar Biol, 2002, 140(4):747-754. doi: 10.1007/s00227-001-0746-9
-
期刊类型引用(9)
1. 牛明香,李永涛,王俊,刘志国,左涛,程兆龙. 基于HSI的黄河口近岸海域鮻鱼卵、仔稚鱼栖息地适宜性研究. 海洋环境科学. 2023(05): 766-773 .  百度学术
百度学术
2. 范秀梅,杨胜龙,张胜茂,朱文斌,崔雪森. 基于栖息地指数的阿拉伯海鲐鱼渔情预报模型构建. 南方水产科学. 2020(04): 8-17 . 本站查看
3. 孙霄,张云雷,徐宾铎,张崇良,任一平,薛莹. 海州湾及邻近海域短吻红舌鳎产卵场的生境适宜性. 中国水产科学. 2020(12): 1505-1514 . 百度学术
4. 范秀梅,唐峰华,崔雪森,杨胜龙,朱文斌,黄良敏. 基于栖息地指数的西北太平洋日本鲭渔情预报模型构建. 海洋学报. 2020(12): 34-43 . 百度学术
5. 范江涛,黄梓荣,许友伟,孙铭帅,晏然,陈作志. 基于海表温和水深的南海北部短尾大眼鲷渔场分析. 中国水产科学. 2019(01): 26-33 . 百度学术
6. 刘祝楠,余景,陈丕茂,于杰,王言丰. 粤西海域灯光罩网渔场时空分布与海洋环境的关系. 南方农业学报. 2019(04): 867-874 . 百度学术
7. 王言丰,余景,陈丕茂,于杰,刘祝楠. 北部湾灯光罩网渔场时空分布与海洋环境关系分析. 热带海洋学报. 2019(05): 68-76 . 百度学术
8. 晏然,范江涛,徐姗楠,许友伟,孙铭帅,陈作志. 南海北部近海竹荚鱼栖息地分布特征. 生态学杂志. 2018(08): 2430-2435 . 百度学术
9. Liu Zhunan,Yu Jing. Response of Spatio-temporal Distribution of Fishery Resources to Mairne Environment in the Northern South China Sea. Animal Husbandry and Feed Science. 2018(Z1): 311-317+320 . 必应学术
其他类型引用(7)


计量
- 文章访问数: 3372
- HTML全文浏览量: 1047
- PDF下载量: 668
- 被引次数: 16
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
