
Preliminary analysis of structure and function of Daya Bay ecosystem based on Ecopath model
-
摘要:
大亚湾是位于广东省东部的重要经济活动区域,渔业资源捕捞和航道运输等行为改变了其生态系统结构和功能,因此,评估和可持续利用渔业资源需要计算该生态系统各功能组之间的相互作用。该研究通过Ecopath with Ecosim(EwE 6.4)软件,利用2012年大亚湾海域渔业资源调查数据将大亚湾生态系统划分为18个功能组和1个碎屑组,整体了解该生态系统能量流动、总体特征和各食物网结构。研究结果表明:大亚湾生态系统各营养级呈现金字塔结构,营养级范围在1~3.29级之间。食物链通道主要有2条,一条为牧食食物链,另一条为碎屑食物链。大亚湾生态系统营养级转换效率较低,生态系统总转换效率仅为8%。大亚湾生态系统总流量为6 249.573 t·(km2·年)-1,系统总生产量为2 827.584 t·(km2·年)-1,总净初级生产量为2 468.36 t·(km2·年)-1,总初级生产量/总呼吸量(TPP/TR)为2.185,Finn循环指数(FCI)和平均能流路径(MPL)分别为4.8%和3.53,联结指数(CI)和系统杂食性指数(SOI)分别为0.324和0.174。综上,大亚湾生态系统食物网简单,稳定性较差,系统处于幼期阶段,亟须加强捕捞限制和资源环境保护。
Abstract:Daya Bay, located in the eastern Guangdong Province, is an important economic region. Human activities such as fishing and waterway transportation have changed its ecological structure and function. Thus, assessment and sustainable utilization of fishery resources require study on the interaction among functional groups in the ecosystem. In the present study, Ecopath with Ecosim (EwE 6.4) software was applied to study the energy flow, general characteristics and food web structure in the whole ecosystem. Daya Bay has two main channels of food chain (grazing food chain and detritus food chain). The conversion efficiency among trophic levels in Daya Bay ecosystem was relatively low, the total energy transfer efficiency being just 8%. The total system throughput of the ecosystem was 6 249.573 t·(km2·year)-1. The total production was 2 827.584 t·(km2·year)-1. The total net primary production was 2 468.36 t·(km2·year)-1. The ratio of total primary productivity to total respiration (TPP/TR) was 2.185. Finn′s cycling index and the mean path length of the energy flow were 4.8% and 3.53, respectively. Connectance index and system omnivory index were 0.324 and 0.174, respectively. To sum up, the Daya Bay ecosystem is at an immature stage which has a simple food web, low maturity and poor stability. It is needed to enhance fishing restriction and improve environmental protection.
-
Keywords:
- Daya Bay /
- Ecopath model /
- ecosystem /
- food web
-
在天然环境和养殖池塘中,对虾摄食的饵料种类非常多,食谱广泛[1-7],但同时对虾对某些饵料种类表现出明显的偏好,在多种饵料同时存在时表现出明显的选择性[8, 9]。以往的研究发现饵料密度、寻觅的难易、处理的难易等因素会影响动物对饵料的选择性[10-16],这种选择性部分或全部地受到使摄食过程中获得的能量净值与处理食物消耗能量的比值最大化规律的支配[12, 17, 18]。在实验条件下,不同饵料均过量同时提供时,尽管对虾偏好某种饵料,对虾也摄食其它饵料[8]。对虾的这种杂食性行为显然具有某些优势,如Chamberlain和Lawrene[20](1981)发现南美白对虾同时摄食鱿鱼、虾类、多毛纲动物和蛤肉时生长要比单独摄食其中任何一种饵料时生长要快。日本对虾同时摄食鳀鱼和蛤时生长比仅摄食其中一种饵料好[8]。对虾杂食性可能具有的其它优点包括通过不同饵料的搭配来改善摄入食物的营养平衡和能量特征,从而达到对饵料资源的节约和更有效利用。本实验以中国明对虾Fenneropenaeus chinensis为研究对象,对这些可能的优越性进行探讨。
1. 材料和方法
1.1 饵料的准备
实验用的5种饵料分别为沙丁鱼肌肉(FF,除去头、内脏、骨、鳞、鳍的鱼)、鹰爪虾肌肉(SF,除去头、壳、内脏)、菲律宾蛤仔足肌(CF,蛤蜊的斧足)、沙蚕(PW,日本刺沙蚕)和配合饲料(FD,海马牌配合饲料)。每种饵料在投喂前都切成和配合饲料大小(长4 mm,直径2 mm左右)一致的小块。5种饵料的营养和能量特征见表 1。
表 1 5种饵料及混合摄入饵料的成分(平均值±SE)Table 1. Composition of five provided diets and ingested mixed diets (mean±SE)饵料种类
diets水分/%
moisture蛋白质/%
protein脂肪/%
lipid能量/kJ·g-1
energy能蛋比(E/P)
energy/protein脂蛋比(L/P)
lipid/proteinFF 82.62±1.50 91.41±0.017h 2.40±0.14a 22.61±0.02f 24.74±0.21a 0.026±0.001a SF 88.25±0.44 86.88±0.08g 3.84±0.18b 22.87±0.04g 26.32±0.35b 0.044±0.001b CF 78.19±0.77 66.49±0.06cd 4.31±0.47c 20.99±0.09cd 31.57±0.48de 0.065±0.001c PW 75.66±0.54 70.93±0.11f 11.00±0.18g 21.31±0.10e 30.04±0.78c 0.155±0.011ef FD 8.00±1.04 45.97±0.05a 9.16±0.17d 19.37±0.03a 42.12±0.16g 0.199±0.006e CF+PW …… 70.64±0.04f 10.53±0.06fg 21.29±0.01e 30.13±0.01c 0.149±0.001d CF+FD …… 53.92±1.41b 7.28±0.33d 20.00±0.11b 37.14±0.79f 0.136±0.010f PW+FD …… 64.81±0.73c 10.53±0.05fg 20.83±0.06c 32.15±0.23e 0.162±0.001ef CF+FD+PW …… 67.92±1.01de 10.42±0.16f 21.08±0.08d 31.04±0.35cd 0.153±0.002ef FF+CF+PW+FD …… 67.75±1.09de 9.90±0.32e 21.04±0.08d 31.07±0.37cd 0.146±0.005de FF+SF+CF+PW+FD …… 68.92±1.17ef 9.45±0.15de 21.15±0.09de 30.70±0.39cd 0.137±0.002d 注:同一列中未标有相同字母的数值相互之间存在显著差异。混合投喂处理的各项指标根据实际不同饵料摄入比例加权计算获得
Note: Values without same letter in the same column are different from each other. The values of mixed diet were weighed in accordance with the portion of each diet in total ingested diet.1.2 实验养殖设施及条件
实验在中国海洋大学教育部海水养殖重点实验室进行。对虾按每个水族箱4尾的密度(45 cm×30 cm×30 cm,水体35 L)养殖,室内温度用空调控制;连续充气,每2天换水1/2,所用海水用筛绢过滤;水族箱内溶氧保持在5.5 mg · L-1以上,氨氮含量不高于2.2 mg · L-1,海水温度为25±0.5℃,盐度为30~32,pH值为7.8±0.3;光照周期为14L:10D。
1.3 实验用虾来源及驯化
实验所用对虾从山东省胶南市购买,运回实验室后分至水族箱内按实验条件进行为期7 d的驯化。驯化期间每天过量投饵2次(早上6:00和晚上6:00),投饵后2.5~3 h时间段内清除水族箱内的残饵和粪便。
1.4 实验设计
驯化结束后,对虾禁食24 h,挑选规格一致的对虾用精确到0.1 mg的电子天平称重,每一水族箱内放入体重(湿重)为0.805±0.005 g (平均值±SE)的对虾4尾。实验共持续30 d。实验共设11个处理,分别投喂不同的饵料,前5个处理分别投喂过量的沙丁鱼肉(FF)、虾肉(SF)、蛤蜊肉(CF)、沙蚕(PW)和配合饲料(FD),后6个处理的饵料组合分别为CF+PW、CF+FD、PW+FD、CF+FD+PW、FF+FD+PW+FD、FF+SF+CF+PW+FD,每一种都过量投喂,使每种饵料的剩余量达到投喂量的一半以上。每一处理设4个重复,共用44个水族箱,水族箱的排列采用完全随机化区组设计进行排列。
1.5 样品的收集和测定
挑选实验用对虾的同时取对虾样品3个,每个样品8尾虾,分析对虾初始成分。每种饵料也分别随机取样3个,作为分析饵料成分的样品。每天投喂的饵料都精确称重,投喂后在2.5~3 h时间段内收集残饵,并对每种饵料进行溶失实验,用于校正最后收集的残饵的量。实验结束后,每个水族箱内的虾分别称量湿重,作为一个样品。收集的样品均用烘箱在65℃下烘干至恒重,计算对虾和饵料样品的水分含量。除残饵外,所有样品用凯氏定氮法测定氮含量并换算成粗蛋白质(P)含量(凯氏氮×6.25)。对虾和饵料的粗脂肪含量(L)用索氏抽提法测定。样品的灰份含量用马福炉在550℃下燃烧至恒重测得[21]。样品的能量值(GE)用PARR1281型卡路里计测定。每个样品测定3次。
1.6 数据计算
对虾的增重率(WG, %初始体重)和饵料转化效率(FCE,%)的计算公式分别如下:
$$ \begin{aligned} & W G=100 \times\left(W_t-W_o\right) / W_o \\ & F C E=100 \times\left(W_t-W_o\right) / C \end{aligned} $$ 其中对虾体重为湿重(W,g),而饵料均为干重。Wt为实验结束时对虾的体重,Wo为实验开始时对虾的体重,T为实验持续时间(d),C为摄食量。
相应地,所有样品以干物质(D,g)、蛋白质(P,g)和能量(E,kJ)表示的特定生长率、增重率、饵料转化效率也按以上公式进行计算[22]。
饵料的能量蛋白比(GE/P,kJ · g-1)、脂肪蛋白比(L/P)按如下公式进行计算:
$$ \begin{aligned} & \mathrm{GE} / \mathrm{P}=\mathrm{GE} \div \mathrm{P} \\ & \mathrm{~L} / \mathrm{P}=\mathrm{L} \div \mathrm{P} \end{aligned} $$ 在假设不同单种饵料对混合饵料的转化效率没有相互作用的前提下,预测混合饵料转化效率计算公式如下:
$$ \mathrm{e} F C E=\sum\limits_{i=1}^5 F C E_i \times R_i $$ 其中FCEi为单种饵料投喂时某种饵料的实际饵料转化效率,Ri表示相应饵料在总饵料摄入量中的比例。
1.7 数据的统计分析
对数据的统计分析用SPSS 10.0进行。对数据进行了单因子方差分析和Duncan氏多重比较,以P<0.05作为差异显著的标准。
2. 实验结果
2.1 中国明对虾的生长
实验结束时不同处理的中国明对虾在湿重、干重、蛋白质、能量等方面都存在显著差异,以FF处理最小,相应的湿重、干重、蛋白质、能量分别为0.993、0.189、0.134 g和3.31 kJ,CF+PW处理最大,相应的湿重、干重、蛋白质、能量分别为2.089、0.501、0.343 g和9.13 kJ;对虾以湿重、干重、蛋白质、能量表示的增重率也表现为相同的趋势(表 2)。
表 2 不同形式表示的放养对虾和收获对虾的重量和增长率(平均值±SE)Table 2. The weight of initial shrimp and final shrimp and weight gain (mean±SE)饵料搭配
diets combination放养对虾initial shrimp 收获对虾final shrimp 增重率/% weight gain WW/g DW/g P/g E/kJ WW/g DW/g P/g E/kJ WW DW P E FF 0.807±0.006 0.175±0.001 0.127±0.001 3.26±0.05 0.993±0.063a 0.189±0.001a 0.134±0.001a 3.31±0.17a 22.76±6.37a 7.60±1.56a 5.45±0.59a 1.30±0.13a SF 0.803±0.007 0.174±0.001 0.126±0.001 3.25±0.01 1.090±0.071ab 0.200±0.010a 0.141±0.001a 3.51±0.01a 35.76±3.09ab 14.81±1.82a 11.24±1.77a 8.30±0.1.72a CF 0.805±0.005 0.174±0.002 0.127±0.001 3.25±0.02 1.324±0.007abc 0.270±0.004ab 0.194±0.001ab 4.90±0.13ab 64.55±3.76abc 54.66±3.62ab 53.23±3.58ab 50.76±3.53ab PW 0.807±0.004 0.175±0.001 0.127±0.001 3.26±0.01 1.970±0.175ef 0.472±0.099de 0.326±0.002de 9.13±0.16e 144.19±8.67ef 169.91±9.68de 157.01±9.67de 180.33±9.52e FD 0.801±0.009 0.174±0.001 0.126±0.001 3.24±0.09 1.408±0.097bcd 0.286±0.068ab 0.203±0.008ab 5.32±0.31abc 76.00±6.25bcd 64.90±5.37e 61.43±3.95abc 64.65±4.03abc CF+PW 0.802±0.006 0.174±0.002 0.126±0.001 3.24±0.03 2.089±0.175f 0.501±0.051e 0.343±0.007e 9.42±0.14e 160.88±9.31f 188.68±9.34bcd 172.13±9.89e 191.23±8.74e CF+FD 0.802±0.008 0.174±0.001 0.126±0.001 3.24±0.05 1.607±0.109cde 0.378±0.078bcd 0.268±0.007bcde 7.38±0.48cde 100.39±5.29cde 117.79±6.58de 112.33±6.37bcde 127.86±5.36cde PW+FD 0.803±0.001 0.174±0.001 0.126±0.001 3.24±0.07 1.848±0.171ef 0.430±0.041de 0.301±0.001de 8.38±0.51de 129.86±9.18ef 147.02±7.23bcd 138.41±7.54de 158.16±5.78de CF+PW+FD 0.804±0.002 0.174±0.001 0.126±0.001 3.25±0.03 1.670±0.031cde 0.367±0.095bcd 0.260±0.004bcd 6.72±0.41bcd 107.62±9.57cde 110.82±5.36bcd 105.54±4.29bcd 106.97±4.19bcd FF+CF+PW+FD 0.807±0.004 0.175±0.001 0.127±0.001 3.26±0.01 1.676±0.109cde 0.395±0.035cde 0.277±0.001cde 7.45±0.50cde 107.60±8.65cde 125.67±4.31cde 118.27±4.68cde 128.48±4.33cde FF+SF+CF+PW+FD 0.809±0.001 0.175±0.001 0.127±0.001 3.26±0.01 1.732±0.076def 0.356±0.069cde 0.275±0.009cde 7.31±0.36cde 113.96±9.76de 122.81±7.69cde 116.02±5.17bcde 123.41±4.69cde 注:同一列中没有相同字母上标的数值相互之间存在显著差异。WW、DW、P、E分别表示数值以湿重、干重、蛋白质、能量表示
Note: Values without same letter in the same column are different from each other. WW, DW, P, and E mean values expressed in wet weight, dry weight, protein and energy, respectively.2.2 不同处理中中国明对虾对不同饵料的总摄食量及组成
中国明对虾在不同处理中摄入的各种饵料的比例不同,在所有与PW同时投喂的处理中,PW以干重、蛋白质、能量形式表示时都占总摄入量的大部分,超过65%(表 3)。而在CF+FD处理中,FD占摄入饵料的大部分,以干重、蛋白质、能量表示时分别为61.27%、52.80%、59.43%(表 3)。在所有其它与FD同时投喂的处理中,FD都占总摄食量的一定比例,超过7%(表 3)。
表 3 不同处理中不同饵料的百分比例(平均值±SE)Table 3. Percentages of different diets in different treatments (mean±SE)饵料搭配
diets combination干重dry weight 蛋白质protein 能量energy FF SF CF PW FD FF SF CF PW FD FF SF CF PW FD FF 100b 0a 0a 0a 0a 100b 0a 0a 0a 0a 100b 0a 0a 0a 0a SF 0a 100b 0a 0a 0a 0a 100b 0a 0a 0a 0a 100b 0a 0a 0a CF 0a 0a 100b 0a 0a 0a 0a 100b 0a 0a 0a 0a 100b 0a 0a PW 0a 0a 0a 100b 0a 0a 0a 0a 100b 0a 0a 0a 0a 100b 0a FD 0a 0a 0a 0a 100b 0a 0a 0a 0a 100b 0a 0a 0a 0a 100b CF+PW 0a 0a 6.64±0.87b 93.39±0.86c 0a 0a 0a 6.25±0.82b 93.75±0.82c 0a 0a 0a 6.55±0.86b 93.45±0.86c 0a CF+FD 0a 0a 38.76±6.87b 0a 61.27±6.89c 0a 0a 47.20±7.48b 0a 52.80±7.48b 0a 0a 40.57±7.03b 0a 59.43±7.03c PW+FD 0a 0a 0a 75.49±2.92c 24.50±2.92b 0a 0a 0a 82.542.27c 17.46±2.27b 0a 0a 0a 77.20±2.77c 22.80±2.77b CF+PW+FD 0a 0a 5.23±1.95ab 83.64±4.75c 11.13±3.98b 0a 0a 5.19±1.93ab 87.153.75d 7.66±2.79c 0a 0a 5.26±1.95ab 84.47±4.49c 10.27±3.69b FF+CF+PW+FD 7.09±2.65ab 0a 2.06±0.85a 72.58±6.58c 18.19±4.42b 9.55±3.49ab 0a 2.04±0.23ab 75.926.32c 12.49±3.19b 7.62±2.83ab 0a 2.06±1.19a 73.52±6.51c 16.80±4.14b FF+SF+CF+PW+FD 8.52±1.02a 5.51±0.49a 0.98±0.15a 66.61±5.28c 18.37±4.74b 11.29±1.30b 6.97±0.69ab 0.96±0.31a 68.364.35c 12.42±3.35b 9.11±1.08ab 5.96±0.54a 0.98±0.57a 67.07±5.05c 16.88±4.40b 注:同一行中没有相同字母上标的数值相互之间存在显著差异
Note: Values without same letter in the same column are different from each other.中国明对虾在不同处理中的总摄食量不同,以单独投喂PW的处理最大,以干重、蛋白质、能量表示时分别为1.352、0.959 g、28.810 kJ;单独投喂FF时摄食量最小,相应的量分别为0.197、0.180 g、4.444 kJ;6个混合投喂处理的摄食量都没有超过单独投喂PW处理的摄食量(表 4)。
表 4 不同处理中对虾摄入的饵料干重、蛋白质和能量(平均值±SE)Table 4. Total dry weight, protein, and energy of ingested diets in different treatments (mean±SE)处理treatment 干重/g dry weight 蛋白质/g protein 能量/kJ energy FF 0.197±0.027a 0.180±0.024a 4.444±0.603a SF 0.527±0.044b 0.458±0.038b 12.047±1.012b CF 0.733±0.044bc 0.487±0.029bc 15.379±0.922bc PW 1.352±0.125e 0.959±0.089e 28.810±2.660f FD 1.144±0.033de 0.526±0.015bcd 22.162±0.647de CF+PW 1.150±0.069de 0.813±0.049de 24.490±1.478cde CF+FD 0.976±0.124cd 0.528±0.070bcd 19.530±2.498bc PW+FD 1.250±0.086e 0.809±0.048de 26.028±1.732ef CF+FD+PW 0.933±0.079cd 0.633±0.052cd 19.660±1.648cd FF +CF+PW+FD 0.909±0.114cd 0.614±0.074bcd 19.111±2.356cd FF+SF+CF+PW+FD 0.983±0.068cd 0.676±0.039cd 20.776±1.375cde 注:同一列中未标有相同字母的数值相互之间存在显著差异
Note: Values without same letter in the same column are different from each other.2.3 不同处理中国明对虾对饵料转化效率(FCE)的预测值与实际值
中国明对虾摄食不同饵料后的实际饵料转化效率(FCE)不同,CF+PW处理的FCE最高,以干重、蛋白质、能量表示时分别为27.86%、26.08%、24.47%,单独投喂FF和SF的处理最低,以3种形式表示时都低于7%(表 5)。在不同饵料搭配的处理中,饵料的实际转化效率除在PW+FD和CF+FD+PW两个处理以干重形式表示时略小于单独投喂PW处理外,均高于所有单独投喂一种饵料的处理(表 5)。所有混合投喂处理的实际饵料转化效率都比预测值高,并且除了PW+FD和CF+FD+PW两个处理的实际值与预测值差异不显著外,其它处理的饵料转化效率实际值都显著高于预测值。
表 5 不同处理中的预测饵料转化效率与实际饵料转化效率(平均值±SE)Table 5. The estimated food conversion efficiencies and observed food conversion efficiencies of different treatments(mean±SE)% 处理
treatment干重dry weigh 蛋白质protein 能量energy 预测值
estimated实际值
observed预测值
estimated实际值
observed预测值
estimated实际值
observedFF 6.37 6.37±0.47a 3.50 3.50±0.43a 0.48 0.48±0.13a SF 4.92 4.92±0.54a 3.10 3.10±0.44a 2.22 2.22±0.45ab CF 13.07 13.07±0.86ab 13.89 13.89±0.92ab 10.78 10.78±0.73bcd PW 21.25 21.25±2.75bc 20.05 20.05±2.74bc 19.76 19.76±2.46de FD 9.85 9.85±1.59a 14.71 14.71±2.45ab 9.44 9.44±1.53bc CF+PW 20.71±0.07 *27.86±3.43c 19.66±0.05 *26.08±3.38bc 19.17±0.08 *24.74±3.02e CF+FD 11.10±0.22 *22.03±5.63bc 14.32±0.06 *28.90±4.56c 9.99±0.09 *22.39±5.71e PW+FD 18.45±0.33 20.10±2.41bc 19.12±0.12 21.26±2.82bc 17.41±0.29 19.39±2.27de CF+FD+PW 19.56±0.51 21.11±2.93bc 19.32±0.21 21.58±3.28bc 18.23±0.45 18.05±2.58cde FF +CF+PW+FD 17.93±0.79 *24.67±4.13c 17.68±0.70 *24.92±4.41bc 16.37±0.86 *22.37±3.75e FF+SF+CF+PW+FD 16.91±0.62 *21.91±2.31bc 16.28±0.39 *21.84±2.51bc 15.13±0.58 *19.43±2.07de 注:同一列没有相同字母标记的数值相互之间存在显著差异。标有“*”的实测值显著比预测值大(P < 0.05)
Note: Values without same letter in the same column are different from each other. Observed values with‘*’ were significantly higher than the estimated.3. 讨论
3.1 中国明对虾同时摄食不同饵料的优势
中国明对虾摄食不同的混合饵料后,在生长速度上都比除了PW外的任何单种饵料投喂的处理有了不同程度的提高,并且CF+PW处理的对虾生长比单独投喂PW快(表 2),这与南美白对虾[20]和日本对虾[9]摄食混合饵料时生长比摄食单种饵料时生长快的现象相同,表明对虾通过不同饵料的搭配,能够提高生长速度,并且这些较高的生长速度不是通过摄食量的显著增加来获得的(表 4)。同时,对虾摄食不同的混合饵料后,饵料转化效率绝大多数比摄食单种饵料时有不同程度的提高,并比假设不同饵料同时投喂时相互之间对饵料转化效率无影响前提下预测的饵料转化效率高(表 5),而且这种转化效率的提高不是以提高摄食量来实现的(表 4),表明不同饵料之间存在一种积极的相互作用。这种积极相互作用对对虾摄食策略可能有重要影响,可能是导致对虾杂食性的主要原因之一。一般认为由于喜食饵料的缺乏和季节波动[23, 24],对虾为减少饥饿和营养不良的胁迫而摄入不十分喜食的饵料。但实验条件下发现即使是喜食饵料过量存在时,对虾也摄入部分不十分喜食的饵料[8]。由于对虾在天然环境中最喜食的饵料是有限的,如果只摄食某种饵料就需要花较多的能量觅食,而采用不同饵料同时摄入的策略,同样可以获得与摄食单种较好饵料的相当生长速度和饵料转化效率,这样不仅可以节约觅食的能量消耗,也能更有效地利用饵料资源。
3.2 不同饵料搭配对饵料营养和能量特征的影响
不同饵料同时投喂对虾时,对虾在生长和饵料转化效率比投喂单种饵料时好,这种相互作用的产生有可能是由于不同饵料中的氨基酸组成不同,混合摄入后相互补充了必需氨基酸,产生蛋白质的互补作用或氨基酸互补作用[25],从而提高了整体的转化效率,也有可能是饵料的其它因素起了重要的作用。尽管饵料中哪些因素对这些积极作用起了主要作用还不十分清楚,但在中国明对虾摄入的饵料中,不同混合饵料的营养和能量特征与单种饵料相比都发生变化(表 1)。值得注意的是在混合饵料中饵料的能量蛋白比(E/P)发生了变化,E/P较高的FD与PW混合后,E/P由42.12降低到37.14 kJ · g-1,而其它混合组的E/P都为30.70~32.15 kJ · g-1,这与薛敏等[26]得到的体重为0.368~0.699和1.025~1.525 g的中国明对虾幼虾最适可消化能量与可消化蛋白(DE/DP)比分别为30.88和28.93 kJ · g-1的结果非常接近,也与本实验中使对虾生长最好的单种饵料PW的E/P(30.04 kJ · g-1)非常接近。混合饵料的脂肪含量也比脂肪含量较低的FF、SF、CF有显著提高,达到7.28%~10.53%;并使饵料的脂肪蛋白比(L/P)比FF、SF、CF显著提高,达到0.136~0.162的范围,与使对虾生长最好和饵料转化效率最高的单种饵料PW的L/P值0.155非常接近。而在以往的研究中,都发现了甲壳动物有一个最适的饵料能量蛋白比(E/P)或脂肪蛋白比(L/P),使动物摄入的饵料能量最大限度地转化为生长能[26-29],并且发现在蛋白质得到满足的前提下,能量和脂肪含量的提高都能够显著提高动物对蛋白质的利用率,产生蛋白质的节约作用[19, 26, 29, 30]。在本实验中,可能是由于对虾选择性摄食后,混合饵料中能量蛋白比、脂肪含量和脂肪蛋白比的得到调整,使中国明对虾对饵料的蛋白质利用率提高,从而提高了饵料的转化效率,并获得了不同饵料搭配的积极相互作用。
-
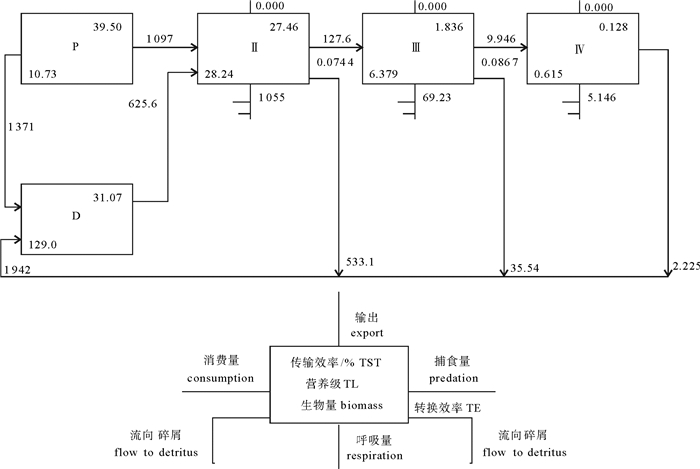
![]()
图 2 大亚湾生态系统营养层级间能流效率示意图
P.初级生产者; D.碎屑; TST.系统总流量
Figure 2. Trophic flow and transfer efficiency through trophic levels in Daya Bay ecosystem
P. primary; D. detritus; TST. total system throughput
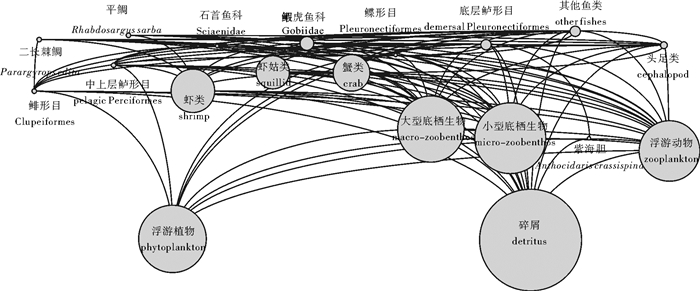
表 1 大亚湾生态系统Ecopath模型功能组及其主要种类
Table 1 Functional groups and main species in Daya Bay ecosystem based on Ecopath model
序号
No.功能组
functional group功能组描述
group′s description拉丁名
Latin name1 二长棘鲷 二长棘鲷 Parargyrops edita Tanaka 2 平鲷 平鲷 Rhabdosargus sarba 3 石首鱼科 白姑鱼 Argyrosomus argentatus 皮氏叫姑鱼 Johnius belengeri 勒氏短须石首鱼 Umbrina russelli Cuvier et Valenciennes 杜氏叫姑鱼 Johnius dussumieri 4 虎鱼科 绿斑细棘虎鱼 Acentrogobius chlorostigmatoides 矛尾虎鱼 Chaeturichthys stigmatias Richardson 巴布亚沟虎鱼 Oxyurichthys papuensis 拟矛尾虎鱼 Parachaeturichthys polynema 长丝虎鱼 Cryptocentrus filifer 5 鲽形目 卵鳎 Solea ovata Richardson 青缨鲆 Crossorhombus azureus 大鳞舌鳎 Cynoglossus macrolepidotus 6 鲱形目 斑 Clupanodon punctatus 康氏小公鱼 Anchoviella commerson 7 底层鲈形目 黑鲷 Sparus macrocephalus 短尾大眼鲷 Priacanthus macracanthus Cuvier et Valenciennes 李氏 Callionymus richardsoni Bleeker 黄斑蓝子鱼 Siganus oramin 列牙 Pelates quadrilineatus 六指马鲅 Polynemus sextarius Bloch et Schneider 丽叶鲹 Caranx kalla Cuvier et Valencienne 黄带鲱鲤 Upeneus sulphureus Cuvier et Valenciennes 印度无齿鲳 Ariomma indica 眶棘双边鱼 Ambassis gymnocephalus 长棘银鲈 Gerres filamentosus Cuvier 少鳞 Sillago japonica 黑鳃天竺鱼 Apogonichthys arafurae 中线天竺鲷 Apogon kiensis Jordan et Snyder 四线天竺鲷 Apogon quadrifasciatus Cuvier et Valenciennes 8 中上层鲈形目 圆鳞发光鲷 Acropoma hanedai Matsubara 短吻鲾 Leiognathus brevirostris 四指马鲅 Eleutheronema tetradactylum 短棘银鲈 Gerres lucidus Cuvier 鹿斑鲾 Leiognathus ruconius 蓝圆鲹 Decapterus maruadsi 9 其他 前鳞骨鲻 Osteomugil ophuyseni 月腹刺鲀 Gastrophysus lunaris 长蛇鲻 Saurida elongata 鲬 Platycephalus indicus 大鳞鳞鲬 Onigocia macrolepis 艾氏蛇鳗 Ophichthus evermanni Jordan et Richardson 10 虾类 宽突赤虾 Metapenaeopsis palmensis 刀额新对虾 Metapenaeus ensis 近缘新对虾 Metapenaeus affinis 鹰爪虾 Trachypenaeus curvirostris 11 虾蛄类 口虾蛄 Oratosquilla oratoria 断脊口虾蛄 Oratosquilla interrupa 条尾近虾蛄 Anchisquilla fasciata 棘突猛虾蛄 Harpiosquilla raphidea 长叉口虾蛄 Oratosquilla nepa 短沟对虾 Penaeus semisulcatus 12 蟹类 矛形梭子蟹 Portunus pelagicus 隆线强蟹 Eucrate crenata 直额 Charybdis truncata 伪装关公蟹 Dorippe facchino 锐齿 Charybdis hellerii 疾进 Charybdis vadorum Alcock 美人 Charybdis callianassa 太阳强蟹 Eucrate solaris Yang et Sun 远海梭子蟹 Portunus pelagicus 秀丽长方蟹 Metaplax elegans 锈斑 Charybdis feriatus 变态 Charybdis variegata 13 头足类 短蛸 Octopus ocellatus Gray 杜氏枪乌贼 Loligo duvaucelii Orbigny 中国枪乌贼 Loligo chinensis Gray 针乌贼 Sepia andreana Steenstrup 田乡枪乌贼 Loligo duvaucelii Orbigny 柏氏四盘耳乌贼 Euprymna berryi Sasaki 14 小型底栖生物 多毛类等 Polychaeta 15 大型底栖生物 覆瓦牡蛎 Parahyotissa imbricate 条蜒螺等 Nerita striata 16 紫海胆 紫海胆 Anthocidaris crassispina 17 浮游动物 桡足类 Copepoda 浮游幼虫等 larval plankton 18 浮游植物 中肋骨条藻 Skeletonema costatum 尖刺菱形藻 Nitzschia pungens Grunow 密聚角毛藻 Chaetoceros coarctatus Lauder 中华盒形藻 Biddulphia sinensis Greville 奇异棍形藻等 Bacillaria paradoxa Gmelin 19 碎屑 有机物  下载: 导出CSV
下载: 导出CSV
表 2 大亚湾生态系统Ecopath模型功能组基本参数(粗体为计算参数)
Table 2 Basic parameters of functional groups in Daya Bay based on Ecopath model (calculating parameters are in bold)
序号
No.功能组
functional group营养级
trophicB/t·km-2 (P/B)/年-1 (Q/B)/年-1 EE P/Q 1 二长棘鲷 3.24 0.012 8 2.825 13.5 0.99 0.21 2 平鲷 3.29 0.016 9 1.76 4.8 0.99 0.36 3 石首鱼科 3.17 0.006 1.85 10.4 0.99 0.18 4 虎鱼科 3.20 0.12 2.66 7.71 0.96 0.35 5 鲽形目 3.06 0.007 3.09 13.61 0.99 0.23 6 鲱形目 2.67 0.02 5.54 29.3 0.99 0.19 7 底层鲈形目 3.19 0.075 3.91 19.4 0.95 0.20 8 中上层鲈形目 2.96 0.014 1.91 10.2 0.99 0.19 9 其他鱼类 3.34 0.07 3.4 14 0.96 0.24 10 虾类 2.67 1.83 6.5 16.35 0.99 0.40 11 虾蛄类 2.88 0.814 5 6 20.3 0.99 0.30 12 蟹类 2.87 1.042 3 5.65 26.9 0.99 0.21 13 头足类 3.18 0.032 3.1 12.8 0.98 0.24 14 大型底栖生物 2.24 10.15 3 12.5 0.71 0.24 15 小型底栖生物 2.14 14.5 6.57 27.4 0.38 0.24 16 紫海胆 2.13 0.011 4.2 16.7 0.16 0.25 17 浮游动物 2.00 6.55 32 192 0.38 0.17 18 浮游植物 1.00 10.732 230 0.44 19 碎屑 1.00 129 0.54
下载: 导出CSV
表 3 大亚湾生态系统总能量分布
Table 3 Distribution of total throughput in Daya Bay ecosystem
t·(km2·年)-1 营养级
trophic level被摄食量
consumption by predation输出量
export流向碎屑量
flow to detritus呼吸量
respiration总流量
throughputⅣ 0.641 0 2.225 5.146 8.012 Ⅲ 9.946 0 35.54 69.23 114.7 Ⅱ 127.6 0 533.1 1 055 1 716 Ⅰ 1 723 1 316 1 371 0 4 410 合计total 1 861 1 316 1 942 1 130 6 249
下载: 导出CSV
表 4 大亚湾各营养级间转换效率
Table 4 Transfer efficiency among trophic levels in Daya Bay
来源source 营养级trophic level Ⅱ Ⅲ Ⅳ Ⅴ 生产者producer 6.7 8.4 9.9 2.5 碎屑detritus 8.7 9.1 5.2 2.9 总能流all flow 7.4 8.7 8 2.6 碎屑所占能流比值proportion of total flow originating from detritus:0.4 转化效率transfer efficiency 初级生产者转化效率from primary producers:8.2% 碎屑转化效率from detritus:7.4% 总转化效率total:8%
下载: 导出CSV
表 5 大亚湾生态系统与其他海域生态系统参数比较
Table 5 Comparison of parameters of Daya Bay ecosystem with those of the other sea areas
参数parameter 莱州湾[5]
Laizhou Bay大亚湾
Daya Bay南海北部[11]
northern South China Sea总消耗量/t·(km2·年)-1 sum of all consumption 1 314.32 1 861.424 3 324.185 总输出量/t·(km2·年)-1 sum of all exports 434.41 1 316.293 3 236.992 总呼吸量/t·(km2·年)-1 sum of all respiratory flows 826.97 1 129.915 2 027.59 流向碎屑总量/t·(km2·年)-1 sum of all flows into detritus 1 042.24 1 941.941 4 448.65 系统总流量/t·(km2·年)-1 total system throughput 3 618 6 249.573 13 037 系统总生产量/t·(km2·年)-1 sum of all production 1 549 2 827.584 5 895 总净初级生产量/t·(km2·年)-1 calculated total net primary production 1 261.38 2 468.36 5 264.58 总初级生产量/总呼吸量total primary production/total respiration 1.53 2.185 2.596 系统净生产量/t·(km2·年)-1 net system production 434.41 1 338.445 3 236.99 总初级生产量/总生物量total primary production/total biomass 25.54 53.655 92 25 总生物量/总流量total biomass/total throughput - 0.007 0.016 总生物量/t·(km2·年)-1 total biomass 51.39 46.0035 212.761
下载: 导出CSV
-
[1] De MUTSERT K, STEENBEEK J, LEWIS K, et al.Exploring effects of hypoxia on fish and fisheries in the northern Gulf of Mexico using a dynamic spatially explicit ecosystem model[J].Ecol Model, 2016, 331:142-150. doi: 10.1016/j.ecolmodel.2015.10.013
[2] HEYMANS J J, COLL M, LINK J S, et al.Best practice in Ecopath with Ecosim food-web models for ecosystem-based management[J].Ecol Model, 2016, 331:173-184. doi: 10.1016/j.ecolmodel.2015.12.007
[3] OCAMPO R M, MILESSI A C, ALEJANDRA R M, et al.Assessing the effects of demersal fishing and conservation strategies of marine mammals over a Patagonian food web[J].Ecol Model, 2016, 331:31-43. doi: 10.1016/j.ecolmodel.2015.10.025
[4] RUIZ D J, BANKS S, WOLFF M.Elucidating fishing effects in a large-predator dominated system:the case of Darwin and Wolf Islands (Galápagos)[J].J Sea Res, 2016, 107(2):1-11. http://adsabs.harvard.edu/abs/2016JSR...107....1R
[5] 林群, 李显森, 李忠义, 等.基于Ecopath模型的莱州湾中国对虾增殖生态容量[J].应用生态学报, 2013, 24(4):1131-1140. http://www.cqvip.com/QK/90626A/201304/45518132.html [6] 吴忠鑫, 张秀梅, 张磊, 等.基于Ecopath模型的荣成俚岛人工鱼礁区生态系统结构和功能评价[J].应用生态学报, 2012, 23(10):2878-2886. http://www.cqvip.com/QK/90626A/201210/43585545.html [7] 李睿, 韩震, 程和琴, 等.基于ECOPATH模型的东海区生物资源能量流动规律的初步研究[J].资源科学, 2010, 32(4):600-605. http://www.oalib.com/paper/1702659 [8] 杨林林, 姜亚洲, 袁兴伟, 等.象山港生态系统结构与功能的Ecopath模型评价[J].海洋渔业, 2015, 37(5):399-408. http://www.doc88.com/p-2032365917443.html [9] 林群, 金显仕, 郭学武, 等.基于Ecopath模型的长江口及毗邻水域生态系统结构和能量流动研究[J].水生态学杂志, 2009, 2(2):28-36. http://www.oalib.com/paper/1595486 [10] 陈作志, 邱永松, 贾晓平, 等.基于Ecopath模型的北部湾生态系统结构和功能[J].中国水产科学, 2008, 15(3):460-468. http://www.cjae.net/CN/abstract/abstract19640.shtml [11] 陈作志, 邱永松.南海北部生态系统食物网结构、能量流动及系统特征[J].生态学报, 2010, 30(18):4855-4865. http://d.wanfangdata.com.cn/Periodical/stxb201018005 [12] 李云凯, 刘恩生, 王辉, 等.基于Ecopath模型的太湖生态系统结构与功能分析[J].应用生态学报, 2014, 25(7):2033-2040. http://www.cqvip.com/QK/90626A/201407/661659815.html [13] 刘恩生, 李云凯, 臧日伟, 等.基于Ecopath模型的巢湖生态系统结构与功能初步分析[J].水产学报, 2014, 38(3):417-425. http://www.doc88.com/p-2721315195196.html [14] 武震, 贾佩峤, 胡忠军, 等.基于Ecopath模型分析分水江水库生态系统结构和功能[J].应用生态学报, 2012, 23(3):812-818. http://d.wanfangdata.com.cn/Periodical/yystxb201203034 [15] 国家海洋局第三研究所.大亚湾海洋生态文集[M].北京:海洋出版社, 1990:273-292. [16] LI K Z, YAN Y, YIN J Q, et al.Seasonal occurrence of Calanus sinicus in the northern South China Sea:a case study in Daya Bay[J].J Mar Syst, 2016, 159:132-141. doi: 10.1016/j.jmarsys.2016.03.014
[17] 王雪辉, 杜飞雁, 邱永松, 等.大亚湾海域生态系统模型研究I:能量流动模型初探[J].南方水产, 2005, 1(3):1-8. http://industry.wanfangdata.com.cn/dl/Detail/Periodical?id=... [18] 何秀婷, 王奇, 聂湘平, 等.广东典型海水养殖区沉积物及鱼体中磺胺类药物的残留及其对人体的健康风险评价[J].环境科学, 2014, 35(7):2728-2735. http://www.cqvip.com/QK/91181X/201407/663121666.html [19] 刘宝林, 张鸿, 谢刘伟, 等.深圳近岸海域全氟化合物的污染特征[J].环境科学, 2015, 36(6):2028-2037. http://industry.wanfangdata.com.cn/yj/Detail/Periodical?id=... [20] 徐姗楠, 李纯厚, 徐娇娇, 等.大亚湾石化排污海域重金属污染及生态风险评价[J].环境科学, 2014, 35(6):2075-2084. doi: 10.13227/j.hjkx.2014.06.006.html [21] 方良, 李纯厚, 杜飞雁, 等.大亚湾海域浮游动物生态特征[J].生态学报, 2010, 30(11):2981-2991. http://www.oalib.com/paper/1403159 [22] HAN J H, KUMAR H K, LEE J H, et al.Integrative trophic network assessments of a lentic ecosystem by key ecological approaches of water chemistry, trophic guilds, and ecosystem health assessments along with an ECOPATH model[J].Ecol Model, 2011, 222(19):3457-3472. doi: 10.1016/j.ecolmodel.2011.08.001
[23] 王增焕, 李纯厚, 贾晓平.应用初级生产力估算南海北部的渔业资源量[J].渔业科学进展, 2005, 26(3):9-15. http://www.cqvip.com/QK/90269X/200503/15851786.html [24] AHLGREN G.Comparison of methods for estimation of phyto-plankton carbon[J].Arch Hydrobiol, 1983, 98(4):489-508. doi: 10.1002/iroh.19790640111/abstract?...
[25] 杜飞雁, 王雪辉, 贾晓平, 等.大亚湾海域浮游动物种类组成和优势种的季节变化[J].水产学报, 2013, 37(8):1213-1219. http://dspace.xmu.edu.cn/bitstream/handle/2288/108228/大亚湾... [26] 舒黎明, 陈丕茂, 秦传新, 等.柘林湾-南澳岛潮间带冬夏两季大型底栖动物种类组成及优势种[J].生态学杂志, 2016, 35(2):423-430. http://www.cje.net.cn/CN/abstract/abstract6348.shtml [27] PALOMARES M L D, PAULY D.Predicting food consumption of fish populations as functions of mortality, food type, morphometrics, temperature and salinity[J].Mar Freshw Res, 1998, 49(5):447-453. doi: 10.1071/MF98015
[28] PAULY D.On the interrelationships between natural mortality, growth-parameters, and mean environmental-temperature in 175 fish stocks[J].ICES J Mar Sci, 1980, 39(2):175-192. doi: 10.1093/icesjms/39.2.175
[29] GEERS T M, PIKITCH E K, FRISK M G.An original model of the northern Gulf of Mexico using Ecopath with Ecosim and its implications for the effects of fishing on ecosystem structure and maturity[J].Deep Sea Res Ⅱ, 2014, 113:255-257. http://www.sciencedirect.com/science/article/pii/S096706451400023X
[30] 陈作志, 邱永松.北部湾二长棘鲷生长和死亡参数估计[J].水产学报, 2003, 27(3):251-257. http://www.oalib.com/paper/5093085 [31] WANG Y, DUAN L J, LI S Y, et al.Modeling the effect of the seasonal fishing moratorium on the Pearl River Estuary using ecosystem simulation[J].Ecol Model, 2015, 312:406-416. doi: 10.1016/j.ecolmodel.2015.06.011
[32] FROESE R, PAULY D. Fishbase[EB/OL].[2016-05-26].http://www.fishbase.org .
[33] 徐姗楠, 陈作志, 何培民.杭州湾北岸大型围隔海域人工生态系统的能量流动和网络分析[J].生态学报, 2008, 28(5):2065-2072. http://www.doc88.com/p-91573405230.html [34] 侯刚, 朱立新, 卢伙胜.北部湾二长棘鲷生长、死亡及其群体组成[J].广东海洋大学学报, 2008, 28(3):50-55. http://cpfd.cnki.com.cn/Article/CPFDTOTAL-OGSB200711001270.htm [35] 徐恭昭.大亚湾环境与资源[M].合肥:安徽科学技术出版社, 1989:274-287. [36] 代春桃. 北部湾二长棘鲷食性和营养级的研究[D]. 湛江: 广东海洋大学, 2010: 40-41. http://so.med.wanfangdata.com.cn/ViewHTML/DegreePaper_Y1806113.aspx [37] 杨国欢, 孙省利, 侯秀琼, 等.基于稳定同位素方法的珊瑚礁鱼类营养层次研究[J].中国水产科学, 2012, 19(1):105-115. http://www.oalib.com/paper/4847733 [38] 徐娇娇. 大亚湾石化排污对邻近海域生态环境的影响及生态系统健康评价[D]. 上海: 上海海洋大学, 2013: 42-53. http://d.wanfangdata.com.cn/Thesis/D372773 [39] 陈作志, 邱永松, 贾晓平, 等.基于Ecopath模型的北部湾生态系统结构和功能[J].中国水产科学, 2008, 15(3):460-468. http://www.cjae.net/CN/abstract/abstract19640.shtml [40] YÁÑEZ-ARANCIBIA A, LARA-DOMÍNGUEZ A L, SÁNCHEZ-GIL P, et al.Estuary-sea ecological interactions:a theoretical framework for the management of coastal environment[J]. Environ Anal Gulf Mexico, Special Publication Series, 2007, 1:271-301. https://www.researchgate.net/profile/Alejandro_Ortega-Argueta/...
[41] LIN H J, SHAO K T, HWANG J S.A trophic model for Kuosheng Bay in northern Taiwan[J].J Mar Sci Technol, 2004, 12(5):424-432. http://www.airitilibrary.com/Publication/alDetailedMesh?DocID=10232796-200412-12-5-424-432-a
[42] ULLAH M H, RASHED-UN-NABI M, AL-MAMUN M A.Trophic model of the coastal ecosystem of the Bay of Bengal using mass balance Ecopath model[J].Ecol Model, 2012, 225(3):82-94. doi: 10.1007/s12562-016-1060-2.pdf
[43] RUIZ-CAUICH L E, SCHMITTER-SOTO J J, BARBA-MACIAS E.Stability vs.organization:potential of a trophic model for the management of shallow tropical streams[J].Food Webs, 2016, 6:38-47. https://www.researchgate.net/publication/294871362_Stability_vs...
[44] 杨文波, 李应仁, 李继龙, 等.国际渔业管理制度对我国渔业资源保护和利用的影响[J].中国渔业经济, 2015, 33(6):23-30. http://www.cqvip.com/Main/Detail.aspx?id=3078742
计量
- 文章访问数: 3621
- HTML全文浏览量: 1120
- PDF下载量: 688
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
