
Study on swimming performance of juvenile bighead carp at different actuating state
-
摘要:
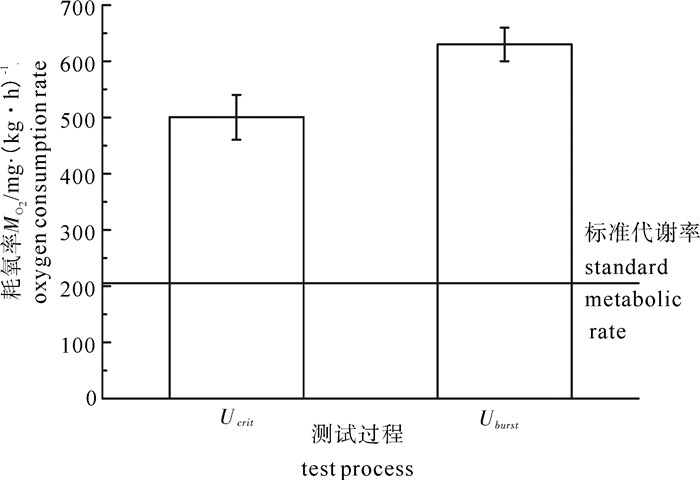
为了探究鱼类反复运动疲劳后游泳能力和游泳行为的变化,以鳙(Aristichthys nobilis)幼鱼为研究对象,利用环形水槽实验装置,采用流速递增法,通过LoliTrack视频软件分析鱼类游泳行为,研究不同运动状态下鳙幼鱼的游泳特性变化情况。结果表明,鳙幼鱼疲劳前耗氧率随游泳速度的增大显著增加,且临界游泳速度恢复过程中最大耗氧率小于突进游泳速度恢复过程中最大耗氧率。视频分析显示,摆尾频率与游泳速度呈线性正相关关系。同时发现,在2组反复疲劳运动状态下,连续进行2次临界游泳速度测定过程中,鳙幼鱼频繁使用爆发-滑行游泳行为,而在经过突进游泳速度测试后转入临界游泳速度测定过程中,实验鱼并未使用这一游泳行为。经过一次临界游泳速度测试后,第二次临界游泳速度显著大于突进游泳速度测试后的临界游泳速度(P < 0.05),第一次突进游泳速度与第二次突进游泳速度无显著差异(P>0.05)。无氧运动消耗导致鳙幼鱼有氧运动能力显著下降,而有氧运动消耗过程对无氧运动能力无显著影响。
Abstract:To investigate the swimming performance and swimming behavior during fatigue cycle,we tested the juvenile Aristichthys nobilis′ swimming capability by increasing velocity with mobile Brett-type swim tunnels by LoliTrack,and then assessed the swimming capability of juveniles at different actuating state.Results show that the oxygen consumption rate (MO2) before swiming fatigue increased with swimming speed significantly. The burst swimming speed MO2 was higher than the critical swimming speed MO2.Video analysis displays that there was significant linear correlation between tail beat frequency and swimming speed. Meanwhile,the juveniles showed burst-coast behavior continuously for two tests at the critical swimming speed in the two groups of repeat fatigue. However,during the transition from the test of burst swinmming speed to the test of critical swimming speed,the juveniles did not show that behavior. After the test of the first critical swimming speed,the second critical swimming speed was higher than that of the test of burst swimming speed. Neither of the burst swimming speed in two tests had significant difference. Anaerobic exercise consumption of bighead carp led to significant decrease of aerobic swimming performance,and aerobic exercise had no effect on anaerobic exercise.
-
Keywords:
- Aristichthys nobilis /
- swimming capability /
- increasing velocity /
- fatigue /
- energy metabolism
-
阳宗海是云南九大高原湖泊之一,属珠江流域南盘江水系,位于中国云南省昆明市,承担着所在流域及下游地区饮用、农业灌溉和工业用水三大功能[1]。随着流域开发强度的增加,阳宗海水体营养盐浓度逐渐升高,20世纪90年代曾多次出现水华现象。2008年砷 (As) 污染发生后,管理部门采取了一系列措施,治理及改善阳宗海水质[2]。截至2020年,阳宗海水质已从2008年的劣Ⅴ类恢复到III类。阳宗海水质虽通过治理取得了显著成效,但仍然面临富营养化风险[3]。多年来,管理部门通过增殖放流向阳宗海投放大量鱼苗来净化水质。据不完全统计,截至2022年,已累计放流以鲢 (Hypophthalmichthys molitrix)、鳙 (Aristichthys nobilis) 为主的1 000多万尾鱼苗 (阳宗海管理部门数据),并实施了严格的禁渔措施。以鱼净水是常用的生物调控方法之一,具有环保、投入低和持续时间长的优点,但前提是须明确鱼类群落结构及其资源现状[4]。安莉等[5]于2011—2012年对阳宗海鱼类资源进行了调查,然而至今未见新的报道,目前湖区渔业资源状况不明。
鱼类资源评估是河流渔业管理和资源保护的基本工作[6],目前常用的方法有数学模型评估[7-9]及水声学评估法[10-11]。数学模型评估主要利用鱼类体长、年龄等较容易获取的捕捞数据进行估算。水声学评估通过对研究区域的监测,可获得鱼类的尺寸规格、时空动态分布、行为特征及资源量等。目前声学技术已成为观测和掌握渔业资源变动的重要手段[12],能够同时在多个营养级 (大型食肉鱼、小型觅食鱼和浮游动物) 进行采样,结合网具捕捞数据,便可进行生物量评估[13-14]。
相关研究表明,鱼类存在昼夜迁移行为,在日落前开始扩散并向上移动,日出后开始聚集并下降[15]。例如,清河水库鲢、鳙昼间聚集于底层,夜间向上层迁移,且分散分布[16]。鱼类的昼夜行为变化导致资源评估结果在昼夜间出现差异。因此,本研究首次利用水声学技术分别于昼间和夜间对阳宗海鱼类资源进行评估,旨在掌握阳宗海鱼类资源现状,分析鱼类昼夜行为差异,评估阳宗海鱼类资源调查的适宜时间,以期为高原湖泊及封闭型水库渔业资源的保护及管理提供参考。
1. 材料与方法
1.1 研究区域
阳宗海 (102°58'E—103°01'E, 24°51'N—24°58'N) 位于昆明市东南,地跨宜良、呈贡和澄江三县,距阳宗镇约4 km,距昆明市区约36 km。阳宗海南北长12 km,东西平均宽2.65 km,流域面积约192 km2,平均水深约20 m,最大水深约30 m,是由地面断裂凹陷发育形成的地堑式断陷湖泊。其最大库容为6.17×109 m3,调查期间库容为5.96×109 m3。
1.2 鱼类调查
结合阳宗海形态特征,在主要入湖口、出湖口、湖湾和洞穴等不同生境处设置了10个采样点 (图1),于2022年3、5、8和10月进行了4次采样,使用网具为单层刺网 (网目0.65、1.2 、1.8 cm)、三层刺网 (网目3.5、6.0、15.0 cm) 和地笼网 (0.4 m×0.4 m×长20 m、1.2 m×1.2 m×长30 m),每个采样点放置不同规格网具各1张。于18:30—19:30下网,次日05:00—07:00收网,采集水深范围约为0~10 m,刺网采集水层为中上层,地笼为底层。渔获物现场鉴定、测量全长、体长、体质量等信息,鉴定主要依据文献 [17-20]。
1.3 声学调查
水声学设备为分裂波束鱼探仪 EY60 (Simrad,挪威),换能器中心频率为120 kHz,发射功率为200 W,−3 dB波束宽为7.0°,脉冲宽度为256 μs。通过不锈钢支架将换能器固定在探测船只右舷,距离船首1/3船体长度,入水深度0.5 m,波束垂直向下发射。通过Garmin 60CSx来实时确定探测位置,鱼探仪与笔记本电脑相连,实时显示和存储数据。探测之前采用碳化钨标准球 (Φ 23 mm) 对换能器进行校正[21]。
夜间和昼间的探测时间分别为2022年6月20日20:00—24:00和21日8:30—12:30。调查船为阳宗海管理部门执法船只,全长约10 m,调查期间湖面较平静,探测时以10 km·h−1的航速匀速行进。由于库区整体呈狭长型,且岸线整齐,采用非固定间距平行航线,在实际走航中略有偏差 (图1),总航程约38.8 km,调查覆盖率为6.47,达到了Aglen[22]提出的覆盖率在6.0 以上的标准。
1.4 数据分析
用Pinkas的相对重要性指数 (Index of relative importance, IRI) 研究鱼类群落优势种, 计算公式如下[23]:
$$ \mathrm{I}\mathrm{R}\mathrm{I}=(N+W)\times F $$ (1) 式中:N为某一种鱼类的尾数占总尾数的百分比 (%);W为某一种鱼类的生物量占总生物量的百分比 (%);F为某一种鱼类出现的站位数占总站位数的百分比 (%)。本文选取IRI≥500的鱼类为优势种。
声学原始数据 (.raw) 使用Echoview 5.4 软件处理。从声学映像中可看出调查区域鱼类主要为分散分布,因此采用回波计数法处理数据,目标提取使用单体目标检测及鱼体轨迹追踪实现[24]。通过定义表面线和底部线来排除表层1 m以上、底部0.3 m以下的所有声学信号。目标强度 (Target Strength, TS) 最小阈值设置为−58 dB,以确保大部分噪声信号被排除在外,其他区域噪声手动剔除。单体检测、鱼体追踪的参数设置参照Rudstam等[25]的研究。数据分析过程中,将150 m作为一个基本调查单元 (EDSU),单独评估鱼类密度、个体分布和行为特征等。每个 EDSU 的鱼类密度(尾·m−3) 的计算均采用回波计数 (EchoCounting) 方法进行[26],公式如下:
$$ \varphi = \frac{N}{{PV}} $$ (2) 式中:N为探测到的鱼类数目;
$ \varphi $ 为单位体积水体的鱼类数量,即鱼类体积密度;V为每一个ping探测的水体体积;P为分析数据的ping数量。体长与TS的关系使用基于基尔霍夫近似模型的鲢的目标强度经验公式计算[27]:
$$ S_{\mathrm{t}}=20 \log L-67.35 $$ (3) 式中:St为目标强度 (dB);L为体长 (cm)。
为分析不同规格鱼类昼夜行为差异,人为将鱼类回波信号分为3类:小个体 (−60~−53 dB)、中等个体 (−53~−47 dB) 和大个体 (>−47 dB),对应鱼类长度分别为小个体 (2~5 cm)、中等个体 (5~10 cm)和大个体 (>10 cm)。
采用 ArcGIS 10.1 软件进行鱼类资源空间分布建模,将鱼类密度及对应的经纬度数据导入ArcGIS,采用反距离加权插值方法进行栅格插值运算,导出每个栅格的鱼类密度,乘以相应的栅格体积,最后将所有栅格值求和得到阳宗海鱼类资源量。
通过TS、相对游泳速度、运动方向 (垂直和水平方向变化) 4个因子来分析调查期间阳宗海鱼类昼夜行为特征。TS可以表征昼夜鱼类体长差异,相对游泳速度可以体现鱼类昼间游泳速度的差异,鱼类在水平和垂直方向的运动情况可以通过鱼体与换能器之间的角度 (水平方向角度变化范围 0°~360°,垂直方向为−90°~90°) 来判定。
2. 结果
2.1 鱼类种类组成
本年度共采集到鱼类24种,32976尾,总质量约792 kg,隶属于7目14科23属 (表1)。其中泥鳅 (Misgurrnus anguillicaudatus)、黄鳝 (Monopterus albus)、鲫 (Carasius auratus)、花䱻 (Hemibarbus maculatus) 和云南光唇鱼 (Acrossocheilus yunnanensis) 为土著种,其他种类均为外来引入种。根据鱼类IRI值的大小排序,IRI≥500的鱼类为优势种。结果显示,太湖新银鱼 (Neosalanx taihuensis)、鲢、鳙、子陵吻鰕虎 (Rhinogobius giurinus)、䱗 (Hemiculter leucisculus)、间下鱵 (Hyporhamphus intermedius) 为阳宗海鱼类优势种。其中,太湖新银鱼、子陵吻鰕虎、间下鱵等小个体鱼类 (平均体长<6 cm) 在数量上占比超90%;从体质量组成来看,鲢、鳙等大个体鱼类 (平均体质量>1000 g) 占比较大。
表 1 阳宗海鱼类种类组成Table 1. Composition of fish species in Yangzonghai Lake种类
Species数量百分比
Quantity
proportion/%体质量量百分比
Body mass
ratio/%体长范围
Range of body
length/mm体质量范围
Range of body
mass/g相对重要
性指数
IRI栖息水层
Habitat太湖新银鱼 Neosalanx taihuensis 65.18 1.19 25~61 0.16~0.99 6 637 中上层 子陵吻鰕虎 Rhinogobius giurinus 15.43 0.37 20~62 0.10~3.57 1 580 底层 间下鱵 Hyporhamphus intermedius 4.85 0.84 85~170 1.72~11.28 569 上层 䱗 Hemiculter leucisculus 2.88 4.39 45~195 1.20~145.34 727 上层 黄颡鱼 Pelteobagrus fulvidraco 2.43 2.36 33~195 0.71~90.76 479 底层 黄䱂 Hypseleotris swinhonis 2.22 0.06 23~53 0.13~2.35 228 底层 高体鳑鲏 Rhodeus ocellatus 2.06 0.18 19~74 0.16~9.45 224 中下层 瓦氏黄颡鱼 Pelteobagrus vachelli 1.26 1.10 33~165 0.85~72.1 236 底层 鲢 Hypophthalmichthys molitrix 1.23 64.99 77~600 2.34~3 930.00 6 622 中上层 麦穗鱼 Pseudorasbora parva 0.72 0.14 35~94 0.60~15.22 86 底层 鲫 Carasius auratus 0.47 2.09 47~245 3.73~496.18 256 中下层 鳙 Aristichthys nobilis 0.31 17.84 77~620 10.00~4 950.00 1 815 中上层 尼罗罗非鱼 Oreochromis nilotica 0.29 1.55 25~270 0.55~680.00 147 中下层 粘皮鲻鰕虎 Mugilogobius myxodermus 0.17 0.00 20~35 0.20~0.60 5 底层 花䱻 Hemibarbus maculatus 0.14 0.40 103~220 16.61~172.67 43 中下层 大鳞副泥鳅 Paramisgurnus dabryanus 0.09 0.06 87~150 3.47~28.70 12 底层 云南光唇鱼 Acrossocheilus yunnanensis 0.08 0.09 65~141 4.34~130.00 17 中下层 泥鳅 Misgurrnus anguillicaudatus 0.05 0.02 85~130 3.61~21.40 4 底层 青鱼 Mylopharyngodon piceus 0.04 0.64 220~395 138.07~430.70 34 中下层 鲤 Cyprinus carpio 0.04 0.43 173~270 137.94~605.00 24 中下层 草鱼 Ctenopharyngodon idellus 0.04 0.70 137~490 45.90~2 660.00 22 中上层 鲇 Silurus asotus 0.04 0.54 220~500 120.00~2 000.00 12 底层 食蚊鱼 Gambusia affinis 0.01 <0.01 3.4 0.70~0.76 0.2 上层 黄鳝 Monopterus albus <0.01 <0.01 210 9.00 0.01 底层 2.2 鱼类水平分布
从声学映像图可以看出,在深水区,夜间大量弱声信号 (TS −70~−58 dB) 聚集在5~10 m水层,形成密集的声散射层 (图2-a),而昼间鱼类回波信号主要呈分散分布 (图2-b)。对探测江段鱼类资源水平分布进行GIS插值,得到探测江段的鱼类水平分布模型 (图3),昼夜鱼类水平分布主要表现为南北2个浅水区为高密度区,在湖区中心鱼类密度较低。独立样本t检验结果显示,阳宗海昼夜鱼类密度均值无显著性差异 (P>0.05)。对不同调查时间的鱼类密度进行方差齐性检验 (P<0.05),表明在显著性水平0.05下鱼类密度不服从正态分布。使用非参数检验进行分析,Kurskal-Wallis结果显示,阳宗海昼夜鱼类密度存在显著性差异 (P<0.05)。
![]() 图 2 同一区域夜间 (a) 和昼间 (b) 典型声学映像回波图Figure 2. Tipical acoustic echogram of same survey area in nighttime (a) and daytime (b)
图 2 同一区域夜间 (a) 和昼间 (b) 典型声学映像回波图Figure 2. Tipical acoustic echogram of same survey area in nighttime (a) and daytime (b)![]() 图 3 阳宗海鱼类密度昼夜分布图Figure 3. Longitudinal distribution of fish densities in two hydroacoustic detections
图 3 阳宗海鱼类密度昼夜分布图Figure 3. Longitudinal distribution of fish densities in two hydroacoustic detections2.3 鱼类垂直分布
整体上,鱼类主要分布在10 m以浅水层,10 m以下水层的密度极低,大部分区域密度为0。鱼类密度昼夜间分布存在显著性差异 (F=265.36, P<0.001),表现为鱼类在昼间相对于夜间整体向下游动。其中夜间鱼类平均水深为7.52 m,分布在5~10 m水层;昼间鱼类平均水深为9.99 m,分布在6~14 m水层。相关分析结果可知,夜间鱼类密度与水深呈显著性正相关 (P=0.01, r=0.22),而昼间鱼类密度与水深呈显著性负相关 (P<0.01, r=−0.43,图4)。
![]() 图 4 阳宗海昼夜鱼类密度与水深的线性回归图Figure 4. Linear regression diagram of diurnal fish density and water depth in Yangzonghai Lake
图 4 阳宗海昼夜鱼类密度与水深的线性回归图Figure 4. Linear regression diagram of diurnal fish density and water depth in Yangzonghai Lake从个体来看,大个体鱼类在夜间分布范围较大 (四分位距为6.31 m,中位数为9.13 m),在昼间分布范围较小且向上游动 (四分位距为3.00 m,中位数为8.26 m),表明大个体鱼类在垂直方向上的迁移更为活跃。小个体和中等个体鱼类在昼间均向下游动了约2.00 m (图5)。
![]() 图 5 阳宗海不同尺寸鱼类水深分布昼夜差异Figure 5. Diurnal variation of water depth distribution of fishes of different sizes in Yangzonghai Lake
图 5 阳宗海不同尺寸鱼类水深分布昼夜差异Figure 5. Diurnal variation of water depth distribution of fishes of different sizes in Yangzonghai Lake2.4 鱼类行为特征
如图6所示,鱼类TS、相对游泳速度、运动方向 (垂直和水平方向变化) 4个行为因子在昼夜间均存在显著性差异 (P<0.001)。昼间鱼类平均TS大于夜间,均值分别为−54.18和−55.52 dB,四分位数分别为−54.28和−55.93 dB。水平方向上,昼夜间50%鱼类游动方向均与调查船航向相反,昼间四分位距位于120°~180°,夜间位于180°~190°。垂直方向上,昼间第三分位数为负值 (−3.72),表明昼间至少有75%的鱼类向下游动;夜间第三分位数为正值 (0.02),表明夜间至少有75%的鱼类向上游动。相对游泳速度昼间大于夜间,平均值分别为2.80和2.57 m·s−1。
![]() 图 6 阳宗海鱼类昼夜行为特征Figure 6. Violin plot of fish behaviors characteristics in Yangzonghai Lake in daytime and nighttime
图 6 阳宗海鱼类昼夜行为特征Figure 6. Violin plot of fish behaviors characteristics in Yangzonghai Lake in daytime and nighttime2.5 鱼类资源量估算
昼夜鱼类平均密度分别为 (0.033±0.063) 和 (0.038±0.152) 尾·m−3,鱼类密度统计信息见表2。图4为鱼类密度频率分布图,从图中可以看出,鱼类密度右偏,表明阳宗海鱼类低密度单元格较多。基于GIS反距离权重插值结果,对调查区域资源量进行估算。对阳宗海底图进行投影,计算得出整个调查区域面积约为37 km2,调查期间库容为5.96×109 m3,初步估算,阳宗海资源量为1.96×107~2.25×107 尾。
表 2 不同时间鱼类密度分布Table 2. Fish density distribution in different periods变量
Variable鱼类平均密度
Average fish density/(尾·m−3)95%置信区间
95% confidence interval/(尾·m−3)偏度
Skewness峰度
Kurtosis资源量
Abundance/(107 尾)夜间 Nighttime 0.0378±0.153 0.020~0.045 7.964 71.824 2.25 昼间 Daytime 0.0329±0.064 0.001~0.066 2.683 6.849 1.96 3. 讨论
3.1 阳宗海鱼类种群变化
云南高原湖泊和水库经历了从自然渔业生产到现代渔业生产的转变,鱼类区系改变巨大[9]。历史资料记载的阳宗海有鱼类18种[19-20,28],由于过度捕捞、环境污染、外来鱼种的引入,阳宗海鱼类群落结构现已发生巨大变化[4,29]。2022年不同月份的4次采样,共采集到鱼类24种,其中土著种仅5种,分别为泥鳅、黄鳝、鲫、花䱻和云南光唇鱼,均为广布种,其中花䱻和云南光唇鱼为近年增殖放流引入。从种类组成来看,土著种占20.8%;从渔获量看,土著鱼类的数量仅占总量的0.88%,除鲫有一定的资源量外,其他种类数量极少。阳宗海特有种中仅阳宗白鱼 (Anabarilius yangzongensis) 于2010年采集到1尾,其他种类如阳宗金线鲃 (Sinocyclocheilus yangzongensis)、短尾鱊 (Acheilognathus brevicaudatus)、阳宗海云南鳅 (Yunnanilus yangzonghaiensis) 和纺锤云南鳅 (Y. elakatis) 已多年未见[4]。阳宗海土著鱼类资源严重衰退,目前已形成了以外来鱼类为优势种、土著鱼类种类和数量不断减少的新群落结构,这也与云南其他高原湖泊鱼类种群的变迁基本一致[30-32]。
从鱼类生态习性角度来看,主要优势种子陵吻鰕虎为底栖鱼类,由于渔探仪存在水底探测盲区,加之在数据处理中去除了底部30 cm以上的水体,因此该生态类型鱼类未包括在声学结果分析中;另外,太湖新银鱼为弱声学散射体,且在夜间聚集分布,因此声学数据处理时需单独分析。阳宗海其他种类均适用于水声学方法进行资源评估。
3.2 鱼类空间分布的影响因素
一般认为鱼类群落的动态变化受内源性和外源性因素影响[33]。内源性因素指鱼类自身繁殖、觅食、洄游及种内、种间竞争等行为导致的种群在数量、时空上的变化;外源性因素指栖息地、水文、环境等改变引起的变化[34]。阳宗海南部有阳宗大河、七星河、摆衣河等入湖河流,且湖泊南部人口密集,人类活动造成的污染较大,北部有火电厂的冷却水排入,导致湖泊南北两侧水体营养盐浓度高于其他区域,可为鱼类提供更为丰富的饵料生物[3]。因此鱼类高密度区主要集中在其南部和北部。
阳宗海鱼类垂直分布的主要影响因子是水深和水温。水深是鱼类生活空间的反映,鱼的体型大小、生活习性等决定了鱼类对水深的需求[35]。Miranda等[36]研究表明,鱼类分布与水深呈现出非随机关系,并根据季节和种类而变化,约75%的物种分布在12 m以浅。阳宗海鱼类主要分布在水体中上层,深水区密度较低,这与云南其他高原湖库的研究结果类似[5,14]。
鱼类缺乏调节体温的能力,其体温随着环境水温的变化而改变,并对温度变化产生有限的适应,水温过高或过低均会对鱼类的生长、活动产生影响[5]。阳宗海是暖单次混合型湖泊[37],存在明显的季节性分层与混合现象。在夏季存在明显的温跃层,表层相对恒温,介于21~24 ℃,温跃层位于9~13 m,与表层温度相差高达7~8 ℃[3];随着水温出现分层,溶解氧也呈现出与水温分层相似的垂直分层结构[38]。本次声学调查时间为夏季,此时温跃层的水温和溶解氧急剧下降,因此阳宗海鱼类大多分布在温跃层以上,即10 m以内水层。
3.3 鱼类行为昼夜差异的影响
鱼类行为在昼夜间存在显著性差异,主要表现为中小型鱼类在夜间上浮、昼间下沉的特点,而夜间大个体鱼类垂直迁移更为活跃,符合鲢、鳙等鱼类的生态习性[16]。鱼类昼夜行为节律差异的成因主要是昼夜光照和温度差异,促使浮游动植物等饵料生物呈现周期性昼夜垂直迁移行为[39],间接影响到鱼类的活动。另外,小型鱼类为逃避敌害白天会躲避于水底,夜间上浮觅食[40]。但这些差异也并非绝对,对突尼斯深水型水库的研究表明,不同季节生物量在昼夜间存在差异,表现为在冬春季,白天高于夜间,夏季则夜间高于白天,在多种鱼类混栖的水体中,昼夜差异虽然存在,但规律不同[41-42]。基于阳宗海鱼类昼夜分布情况,未来阳宗海鱼类声学调查可基于不同目的选择不同的调查策略。如:以太湖新银鱼等小型鱼类为主要调查对象,可在夜间进行调查,声学数据处理方法可选用回波积分法;以鲢、鳙等大中型鱼类为调查对象,可选择在白天调查,声学数据处理方法可选用回波计数法。
夜间大量弱声信号 (TS<−58 dB) 聚集在5~10 m水层,形成密集的声散射层 (图2-a)。本研究主要针对鲢、鳙等大中型鱼类资源进行估算,因此在数据分析时,将TS最小阈值设置为−58 dB,在资源量估算中,亦未将这部分弱声信号包含在内。结合渔获调查结果及鱼类生态习性,推测是太湖新银鱼等小型鱼类。
3.4 鱼类目标强度的影响因素
种类鉴别是渔业资源声学评估中的一个难点。由于淡水鱼类分布特征为多种类混栖,且多为分散分布,通常研究人员采用声学调查结合渔获物采样来分配积分值,以评估资源量[43]。此次调查过程中虽未同步进行渔获物采样,但通过不同季节的采样,基本上掌握了阳宗海鱼类的种类组成及群落结构。除鲢、鳙外,目前缺乏阳宗海其他主要优势种类 TS 值与鱼体大小关系的系统研究,无法准确根据鱼类TS值计算出相应的鱼类大小,因此本研究选择主要优势种鲢的体长与TS的经验公式。鱼类TS值受脉冲发射频率、有无鱼鳔、鱼类在波束中的位置、鱼类在波束中的倾角和河流环境等因素影响[44-45],因此文中估算出的鱼类尺寸会与实际有所偏差。本研究中鱼类TS值主要用于探讨不同规格的鱼类在昼夜行为上的差异,鱼类资源评估主要是通过鱼类密度估算得出。因此TS经验公式的选取对于讨论鱼类资源变化的影响较小,研究结果能够较为准确地反映湖泊鱼类的昼夜行为差异和资源空间分布结构。
3.5 阳宗海渔业资源保护与利用建议
随着渔业方式的转型升级,近年来大水面生态渔业逐步推广。其主要目的是通过自然修复和人工群落调控的手段维护湖泊生态系统健康、实现营养盐移出和净水渔业功能[46]。结果显示,小型鱼类在阳宗海鱼类群落中的占比很高,仅太湖新银鱼、子陵吻鰕虎在渔获物中的数量占比就超过了80% (表1);在声学调查结果中,小于−51 dB (6 cm)的个体占总数的94.5%,这些信号大部分来源于高体鳑鲏 (Rhodeus ocellatus)、黄䱂 (Hypseleotris swinhonis)、麦穗鱼 (Pseudorasbora parva) 等小型鱼类,这些鱼类的主要摄食对象为浮游动物,导致浮游植物生物量增加[47]。小型鱼类过多会对湖泊浮游动物种群造成巨大压力,导致湖泊浮游动物种类、丰度出现不同程度的下降[1],不利于浮游植物的调控[48]。研究表明,在洱海中通过对西太公鱼 (Hypomesus nipponensis) 的定量清除来加强浮游动物的牧食,进而控制浮游植物的密度和生物量,对于防控蓝藻水华爆发具有重要意义[49]。据了解,除太湖新银鱼的专项捕捞外,阳宗海实施了严格的禁渔制度。另外,资源评估结果显示,鲢、鳙的生物量为1.83~2.11 g·m−3,该生物密度远未达到可以有效遏制水华的生物水平[48]。本研究建议:1) 在每年开展太湖新银鱼捕捞的基础上,同时对其他小型鱼类进行生态捕捞;可投放阳宗海所属水系的中上层土著肉食性鱼类,将小型鱼类转化为优质水产品;2) 持续进行鲢、鳙等鱼类的增殖放流,以达到以鱼净水的目的;3) 对阳宗海生态容量进行评估,以制定更加科学合理的湖泊保护利用管理策略。
-
![]()
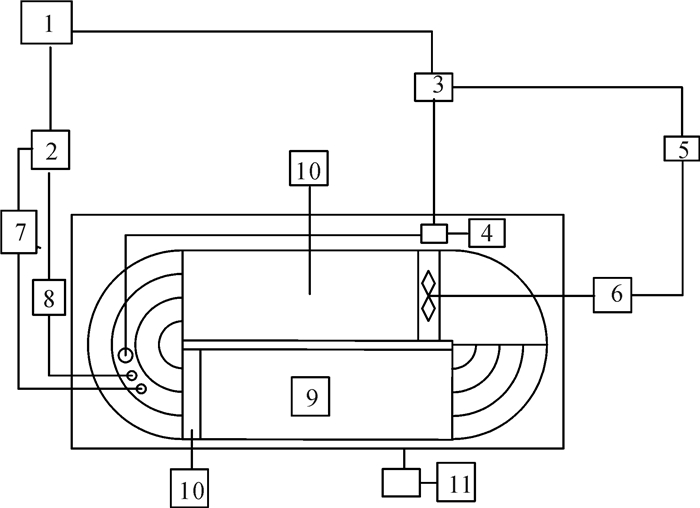
图 1 鱼类游泳能力测定装置示意图
1.笔记本电脑;2. Witrox1温度溶氧自动探测器;3.DAQ-M水泵控制器;4.水泵;5.动力输出控制器;6.电机;7.溶氧探头;8.温度探头;9.游泳区域;10.整流装置;11.摄像头
Figure 1. Test equipment of fish swimming capability
1.laptop; 2.fiber optic oxygen instrument Witorx1;3.controller of pump-DAQ-M; 4.pump; 5.wall outlet controller; 6.motor power; 7.dissolve oxygen probe; 8.temperature probe; 9.swimming tunel; 10.flow streamen; 11.camera
![]()
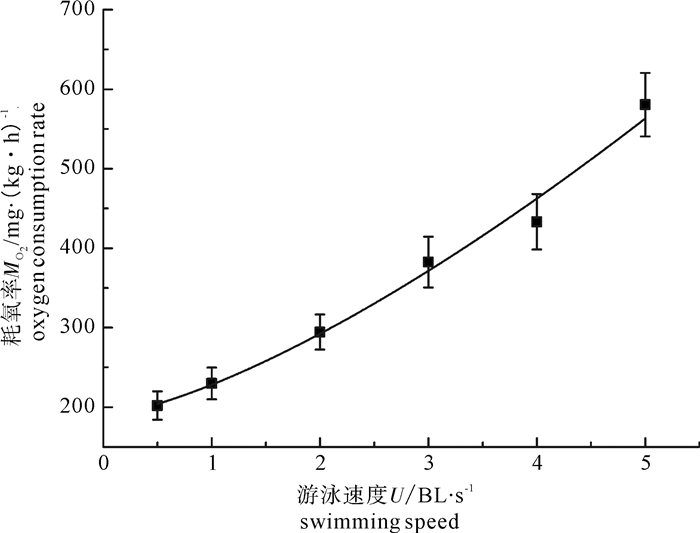
图 2 鳙耗氧率MO2(X±SD)与临界游泳速度U关系
Figure 2. Relatiouship of oxygen consumption rate and swimming speed of bighead
![]()
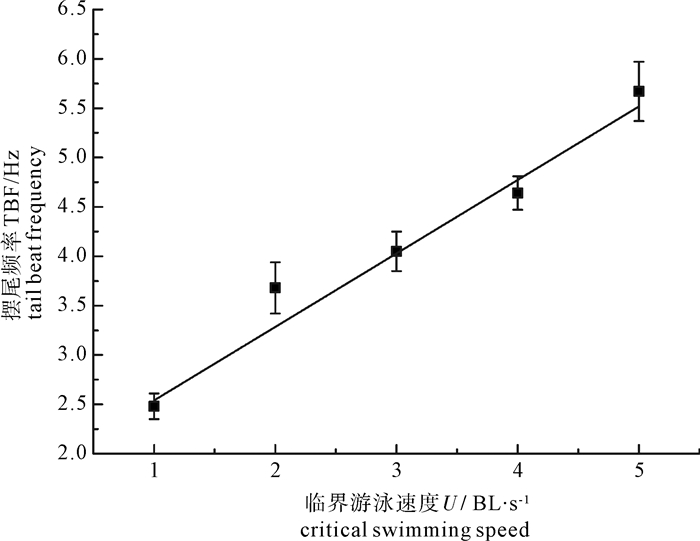
图 4 鳙摆尾频率与临界游泳速度关系
Figure 4. Relationship of tail beat frequency and swimming speed of bighead
![]()
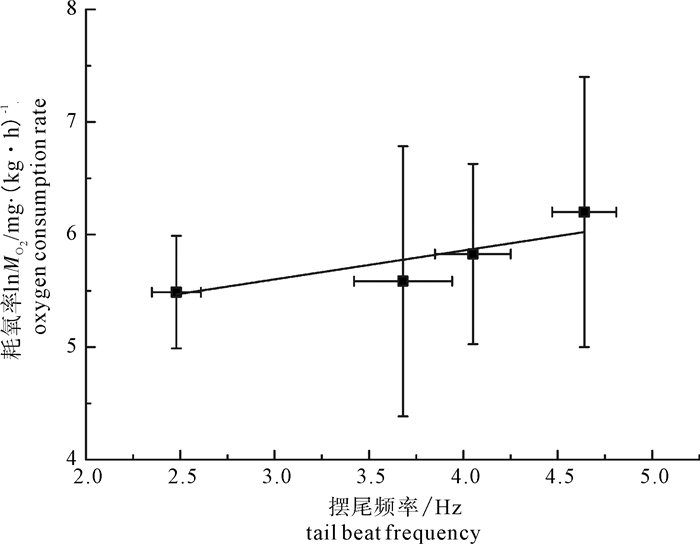
图 5 鳙耗氧率lnMO2(X±SD)与摆尾频率关系
Figure 5. Oxygen consumption rate of bighead in relation to tail beat frequency
![]()
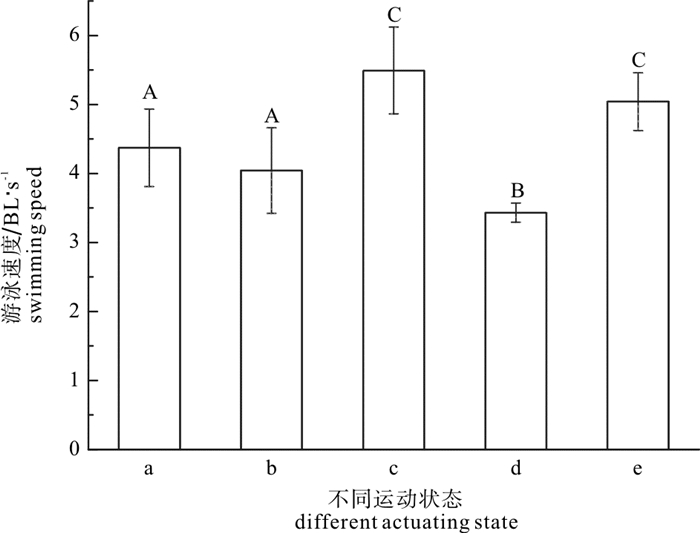
图 6 鳙幼鱼不同运动状态下游泳速度恢复情况
a.组1实验中第一次临界游泳速度;b.组1实验中第二次临界游泳速度;c.组2实验中第一次突进游泳速度;d.组2实验中临界游泳速度;e.组2实验中第二次突进游泳速度;方柱上不同大写字母表示差异显著(P<0.05)。
Figure 6. Recovery of swimming speed of juvenile bighead at different actuating state
a. degree of the first critical swimming speed in Group 1;b. degree of the second critical swimming speed in Group 1;c. degree of the first burst swimming speed in Group 2;d. degree of the critical swimming speed in Group 2;e. degree of the second burst swimming speed in Group 2;values with different capital letters on the bar were significantly different(P<0.05).
![]()
图 3 鳙幼鱼恢复期内2种运动方式耗氧率情况
Figure 3. Oxygen consumption rate of juvenile bighead by two different methods of exhaustion
表 1 不同运动状态下鳙幼鱼游泳能力
Table 1 Swimming ability of juvenile bighead carp at different actuating state
游泳速度
swimming speed组1 Group 1 组2 Group 2 第一次
first第二次
second第一次
first第二次
second绝对临界游泳速度/cm·s-1
absolute critical swimming speed56.92±7.49 51.12±7.66 37.04±0.98 相对临界游泳速度/BL·s-1
relative critical swimming speed4.37±0.56a 4.04±0.62 a 3.43±0.14 b 绝对突进游泳速度/cm·s-1 absolute burst swimming speed 56.17±7.09 51.81±6.35 相对突进游泳速度/BL·s-1
relative burst swimming speed5.49±0.63c 5.04±0.42 c 注:同一行上标不同字母表示差异性显著(P<0.05)。
Note:Values with different superscript letters were significantly different (P<0.05). 下载: 导出CSV
下载: 导出CSV
-
[1] 吴伟军, 何安尤, 施军, 等.红水河四大家鱼资源现状调查分析[J].南方农业学报, 2016, 47(1):134-139. http://d.wanfangdata.com.cn/Periodical_gxnykx201601024.aspx [2] 武仙竹, 肖琳.三峡地区渔业资源及渔业经济发展的历史考察[J].重庆师范大学学报(自然科学版), 2016, 33(1):47-54. http://www.wenkuxiazai.com/doc/6262f0887e21af45b207a803-3.html [3] 熊飞, 刘红艳, 段辛斌, 等.长江上游宜宾江段渔业资源现状研究[J].西南大学学报(自然科学版), 2015, 37(11):43-50. http://industry.wanfangdata.com.cn/dl/Detail/Periodical?id=... [4] 袁喜, 李丽萍, 涂志英, 等.鱼类生理和生态行为对河流生态因子响应研究进展[J].长江流域资源与环境, 2012, 21(S1):24-29. http://www.cqvip.com/QK/97642X/2012S1/1002325382.html [5] 高东红, 刘亚坤, 高梦露, 等.三维鱼道水力特性及鱼体行进能力数值模拟研究[J].水利与建筑工程学报, 2015, 13(2):103-109. http://industry.wanfangdata.com.cn/dl/Detail/Periodical?id=... [6] 刘志雄, 周赤, 黄明海.鱼道应用现状和研究进展[J].长江科学院院报, 2010, 27(4):28-31, 35. doi: 10.3969/j.issn.1001-5485.2010.04.007 [7] 莫伟均, 王从锋, 刘德富, 等.北盘江2种典型增殖放流鱼类突进游速研究[J].南方水产科学, 2014, 10(3):9-14. http://www.schinafish.cn/CN/abstract/abstract9127.shtml [8] YAN G J, HE X K, CAO Z D, et al.Effects of fasting and feeding on the fast-start swimming performance of southern catfish Silurus meridionalis[J].J Fish Biol, 2015, 86(2):605-614. doi: 10.1111/jfb.2015.86.issue-2
[9] 袁喜, 黄应平, 靖锦杰, 等.铜暴露对草鱼幼鱼代谢行为的影响[J].农业环境科学学报, 2016, 35(2):261-265. doi: 10.11654/jaes.2016.02.008 [10] 蔡露, 刘国勇, TAUPIER R, 等.齐口裂腹鱼幼鱼疲劳后游泳特性恢复状况研究[J].水生生物学报, 2013, 37(6):993-998. http://or.nsfc.gov.cn/handle/00001903-5/279630 [11] CAI L, CHEN L, JOHNSON D, et al.Integrating water flow, locomotor performance and respiration of Chinese sturgeon during multiple Fatigue-Recovery cycles[J].PLoS ONE, 2014, 9(4):e94345. doi: 10.1371/journal.pone.0094345
[12] PON L B, HINCH S G, COOKE S J, et al.Physiological, energetic and behavioural correlates of successful fishway passage of adult sockeye salmon Oncorhynchus nerka in the Seton River, British Columbia[J].J Fish Biol, 2009, 74(6):1323-1336. doi: 10.1111/jfb.2009.74.issue-6
[13] 蔡露, 房敏, 涂志英, 等.与鱼类洄游相关的鱼类游泳特性研究进展[J].武汉大学学报(理学版), 2013, 59(4):363-368. http://www.cqvip.com/QK/93869A/201304/47147298.html [14] 吴国犀, 刘乐和, 王志玲.葛洲坝水利枢纽坝下宜昌江段鱼类区系和渔获物组成[J].淡水渔业, 1988(3):8-13. http://www.cnki.com.cn/Article/CJFDTOTAL-DSYY198803002.htm [15] 牛宋芳, 路波, 罗佳, 等.鲢快速逃逸游泳行为研究[J].水生生物学报, 2014, 39(2):394-398. http://www.cnki.com.cn/Article/CJFDTotal-SSWX201502020.htm [16] 熊锋, 王从锋, 刘德富, 等.松花江流域青鱼, 草鱼, 鲢及鳙突进游速比较研究[J].生态科学, 2014, 33(2):339-343. http://www.cnki.com.cn/Article/CJFDTOTAL-STKX201402025.htm [17] YUAN X, LI L P, TU Z Y, et al.The effect of temperature on fatigue induced changes in the physiology and swimming ability of juvenile Aristichthys nobilis (bighead carp)[J].水生生物学报, 2014, 38(3):505-509. [18] TU Z Y, YUAN X, HAN J C, et al. Aerobic swimming performance of juvenile Schizothorax chongi (Pisces, Cyprinidae) in the Yalong River, southwestern China[J].Hydrobiologia, 2011, 675(1):119-127. doi: 10.1007/s10750-011-0809-y
[19] HERSKIN J, STEFFENSEN J F. Energy savings in sea bass swimming in a school:measurements of tail beat frequency and oxygen consumption at different swimming speeds[J].J Fish Biol, 1998, 53(2):366-376. doi: 10.1111/jfb.1998.53.issue-2
[20] LOWE C G. Metabolic rates of juvenile scalloped hammerhead sharks (Sphyrna lewini)[J].Mar Biol, 2001, 139(3):447-453. doi: 10.1007/s002270100585
[21] PENGHAN L Y, CAO Z D, FU S J.Effect of temperature and dissolved oxygen on swimming performance in crucian carp[J].Aquat Biol, 2014, 21(1):57-65. doi: 10.3354/ab00571
[22] STEINHAUSEN M F, STEFFENSEN J F, ANDERSEN N G.The effects of swimming pattern on the energy use of gilthead seabream (Sparus aurata L.)[J].Mar Freshw Behav Physiol, 2010, 43(4):227-241. doi: 10.1080/10236244.2010.501135
[23] CAI L, LIU G Y, TAUPIER R, et al. Effect of temperature on swimming performance of juvenile Schizothorax prenanti[J].Fish Physiol Biochem, 2014, 40(2):491-498. doi: 10.1007/s10695-013-9860-0
[24] 张沙龙, 侯轶群, 王龙涛, 等.长丝裂腹鱼的游泳能力和游泳行为研究[J].淡水渔业, 2014, 44(5):32-37. doi: 10.3969/j.issn.1000-6907.2014.05.007 [25] 房敏, 蔡露, 高勇, 等.温度对鲢幼鱼游泳能力及耗氧率的影响[J].水生态学杂志, 2013, 34(3):49-53. http://www.cnki.com.cn/Article/CJFDTOTAL-SCAN201303012.htm [26] 蔡露, 涂志英, 袁喜, 等.鳙幼鱼游泳能力和游泳行为的研究与评价[J].长江流域资源与环境, 2012, 21(S2):89-95. http://www.cqvip.com/QK/97642X/2012S2/1002325528.html [27] 张永泉, 尹家胜, 徐革峰, 等.水温和体重对白斑红点鲑临界游泳速度和游动耗氧率的影响[J].水生生物学报, 2015, 39(4):661-668. doi: 10.7541/2015.88 [28] LEE C G, FARRELL A P, LOTTO A, et al. Excess post-exercise oxygen consumption in adult sockeye (Oncorhynchus nerka) and coho (O.kisutch) salmon following critical speed swimming[J].J Exp Biol, 2003, 206(18):3253-3260. doi: 10.1242/jeb.00548
[29] PEAKE S J, FARRELL A P. Fatigue is a behavioural response in respirometer-confined smallmouth bass[J].J Fish Biol, 2006, 68(6):1742-1755. doi: 10.1111/jfb.2006.68.issue-6
[30] 龚丽, 吴一红, 白音包力皋, 等.草鱼幼鱼游泳能力及游泳行为试验研究[J].中国水利水电科学研究院学报, 2015, 13(3):211-216. http://www.cqvip.com/QK/87185X/201503/665715958.html [31] 涂志英, 袁喜, 王从锋, 等.亚成体巨须裂腹鱼游泳能力及活动代谢研究[J].水生生物学报, 2012, 36(4):682-688. http://med.wanfangdata.com.cn/Paper/Detail?id=PeriodicalPaper... [32] 刘慧杰, 王从锋, 刘德富, 等.双频识别声呐在鱼类资源调查中的应用进展[J].三峡大学学报(自然科学版), 2015, 37(3):28-34. http://www.docin.com/p-1298843559.html [33] MUELLER A M, BURWEN D L, BOSWELL K M. Tail-beat patterns in dual-frequency identification sonar echograms and their potential use for species identification and bioenergetics studies[J].Trans Am Fish Soc, 2010, 139(3):900-910. doi: 10.1577/T09-089.1
[34] 涂志英, 袁喜, 韩京成, 等.鱼类游泳能力研究进展[J].长江流域资源与环境, 2011, 20(S1):59-65. http://www.cnki.com.cn/Article/CJFDTOTAL-CJLY2011S1011.htm [35] VIDELER J J, WEIHS D. Energetic advantages of burst-and-coast swimming of fish at high speeds[J].J Exp Biol, 1982, 97:169-178. http://jeb.biologists.org/content/97/1/169
[36] 郑金秀, 韩德举, 胡望斌, 等.与鱼道设计相关的鱼类游泳行为研究[J].水生态学杂志, 2010, 3(5):104-110. http://www.docin.com/p-618832674.html [37] REIDY S P, KERR S R, NELSON J A. Aerobic and anaerobic swimming performance of individual Atlantic cod[J].J Exp Biol, 2000, 203(2):347-357. http://jeb.biologists.org/content/203/2/347
[38] 房敏, 蔡露, 高勇, 等.运动消耗对草鱼幼鱼游泳能力的影响[J].长江流域资源与环境, 2014, 23(6):816-820. http://industry.wanfangdata.com.cn/dl/Detail/Periodical?id=... [39] PANG X, YUAN X Z, CAO Z D, et al.The effect of temperature on repeat swimming performance in juvenile qingbo (Spinibarbus sinensis)[J].Fish Physiol Biochem, 2015, 41(1):19-29. doi: 10.1007/s10695-014-0002-0
-
期刊类型引用(9)
1. 张漫,胡江,刘双平. 草鱼在深潭变速水流中的游泳运动特征研究. 工程科学与技术. 2025(02): 125-132 .  百度学术
百度学术
2. 钱振家,徐金铖,张成林,余友斌,刘晃. 不同流速对豹纹鳃棘鲈摆尾频率和血液生理的影响. 南方水产科学. 2023(02): 89-97 . 本站查看
3. 蔡露,Katopodis Christos,金瑶,黄应平,韩德举,胡望斌,陈小娟,陶江平,侯轶群. 中国鲤科鱼类游泳能力综合分析和应用. 湖泊科学. 2022(06): 1788-1801 . 百度学术
4. 任开元,沈修俊,王明云,石小涛,刘国勇. 人工饲养倒刺鲃幼鱼的野化训练. 南方水产科学. 2020(03): 18-24 . 本站查看
5. 王晓臣,吕彬彬,邢娟娟,李丹,王彩宁,任胜杰. 黄河上游2种裂腹鱼感应流速及其与体长的关系. 南方水产科学. 2020(04): 47-53 . 本站查看
6. 王萍,陈凯琳,桂福坤,冯德军. 不同规格凡纳滨对虾可持续游泳能力比较研究. 渔业现代化. 2020(05): 8-14 . 百度学术
7. 王瑶,王从锋,刘慧杰,冯三杰,谭宏. 鲢幼鱼运动能量代谢研究. 水生态学杂志. 2019(04): 101-107 . 百度学术
8. 郝文超,王从锋,刘慧杰,王瑶,冯三杰. 不同运动模式下鲢幼鱼游泳特性研究. 淡水渔业. 2018(02): 3-9+15 . 百度学术
9. 李丹丹,陈丕茂,朱爱意,袁华荣,陈文静,龙鑫玲,王文杰. 黑鲷幼鱼力竭运动后代谢酶活性的恢复水平. 南方水产科学. 2018(06): 59-65 . 本站查看
其他类型引用(7)










计量
- 文章访问数: 3204
- HTML全文浏览量: 1049
- PDF下载量: 755
- 被引次数: 16
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
