
Research on grass carp conveyance performance of jet fish pump
-
摘要:
为了分析射流式鱼泵输送草鱼的性能及其损伤因素,文章设计了一台喉管直径为60 mm的射流式鱼泵,开展了草鱼输送实验,并采用高速摄影和计算流体力学方法进行了研究。结果显示,该射流式鱼泵在扬程2.24 m时最高草鱼输送能力达918 kg·h-1,其所需水功率为2.83 kW。进一步的检测表明,部分实验鱼有鳞片脱落的情况,但未出现游泳异常,解剖后也未发现内脏受损等情况;实验鱼在过泵后呼吸频率及部分血液指标存在明显变化,但在24 h内基本可以恢复。数值模拟和高速摄影方法分析得出,剪切层是造成实验鱼泵内鳞片脱落的主要原因,撞击伤是由内流偏转诱导实验鱼撞击泵内壁面产生的,包含压力梯度在内的水力因素都可能使实验鱼产生应激反应。但由于鱼类在泵内时间极短,上述因素都不会致实验鱼死亡。
Abstract:To study grass carps (Ctenopharyngodon idellus) conveyance performance and injuries factors of jet fish pumps (JFP), a JFP with throat diameter of 60 mm was designed and evaluated by high-speed video observation as well as computational fluid dynamics (CFD). The results show that the peak conveyance capacity of JFP can reach 918 kg·h-1 at lift height of 2.24 m with input power of 2.83 kW. No test fish died or with viscera damage after conveyance but part of descaling was found occasionally, which was mainly caused by the shear layer in JFP. Further tests reveal that, after passing the JFP, respiratory rate and some blood indices of the test fish changed but all of them elementarily recovered within 24 h. It should be noted that collision injury might be caused by the deflection of inner flow. The rapid change of pressure may also affect the test fish. However, due to the short passing time in JFP, none of the above factors killed the test fish.
-
Keywords:
- jet fish pump /
- grass carp /
- jet pump /
- computational fluid dynamics (CFD)
-
相比于传统人工取鱼方法的低效率、高劳动强度和较大鱼损,鱼泵在水产养殖业有着极大的应用前景和发展空间。鱼泵是一种以水或空气为介质输送鱼的流体机械,在20世纪四五十年代,美国、日本和苏联等渔业发达国家已开始鱼泵的研制工作[1-5]。根据工作原理,鱼泵可分为离心式鱼泵、真空式鱼泵、气力式鱼泵和射流式鱼泵。其中,真空式鱼泵间歇式的吸、排动作产生较大的无用功耗,导致效率偏低[6-7];气力式鱼泵不能用于活鱼输送;离心式鱼泵产量高但鱼损较大[8]。射流式鱼泵利用高压水流卷吸鱼水混合流体从而实现鱼类输送[9-13],它本质上是一类环形射流泵。射流式鱼泵具有结构简单、泵内不含运动部件、加工和维护方便、制造和运行成本低、活鱼输送中鱼损小和可输送鱼的种类多等特点,是一种非常有潜力的鱼泵。美国ETI公司1988年研制了SILKSTREAM射流吸鱼泵并拥有其专利权,但相关参数无详细介绍。叶燮明等[5]对射流式鱼泵原理进行了研究。黄小华等[12]提出了射流式鱼泵的设计方案并对其性能进行了计算。XIAO等[14]观察了草鱼在射流式鱼泵内的运动情况并对鱼在扩散管内游动进行了力学分析。
如何在提高鱼泵输送能力的同时,使鱼泵对鱼类的损伤保持在较低水平在水产业中有着十分重要的意义。鱼类通过水力机械(鱼泵、压力通道和水轮机)时可能受到机械损伤和压力伤(由于压力的迅速降低)。机械损伤主要由于旋转部件造成,如水轮机和水泵的叶片。压力损伤更为常见,当鱼周围的压力迅速降低时,鱼鳔内的气体膨胀,可能导致鱼鳔破裂、鱼自身失去平衡,同时鱼眼可能受到压力作用而突出甚至损伤[15-25]。同时,剪切层、不溶气和空化等因素都可能对鱼造成损伤[26]。近年来,计算流体力学(CFD)方法逐渐成为学者们研究鱼压力损伤的重要方法之一[27]。
综上所述,目前射流式鱼泵相关研究还在起步阶段,很少涉及泵内流动及鱼泵的输鱼能力等问题。虽然射流式鱼泵内部没有运动部件,流道宽敞光滑,但其内部存在剪切力、湍流、压力梯度、回流和空化等[13-17],这些水力因素可能对鱼造成一定的伤害,但目前尚未见有文献报道。为此,文章以草鱼(Ctenopharyngodon idellus)为例,通过实验和计算流体力学(CFD)方法对射流式鱼泵的内部流场进行研究,得到该泵不同运行参数(简称工况)下的输送能力,并通过高速摄影与生理指标检测,分析草鱼在泵内的输送过程和可能损伤的部位,为射流式鱼泵结构的进一步优化以及鱼类输送研究提供参考。
1. 材料与方法
1.1 实验装置
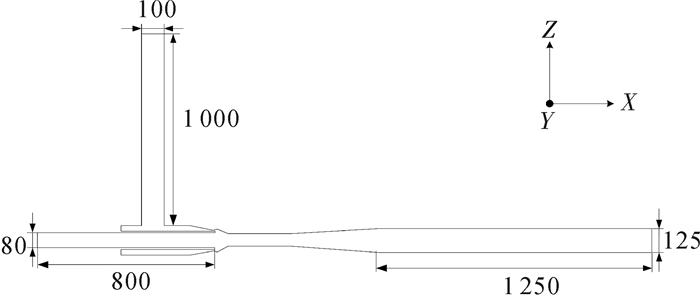
实验装置为借鉴环形射流泵相关设计理论[28-29]自主设计的射流式鱼泵,主要由环形喷嘴、吸入室、喉管和扩散管等组成,其结构如图 1所示。其工作原理为高压工作流体从环形喷嘴高速喷出,卷吸含有鱼水混合物的被吸流体,两股流体在收缩室及喉管中混合并进行动量交换,使被输送流体的动能增大,最后通过扩散管将大部分动能转换为压能,鱼水混合物通过出口直管输送到目的地。
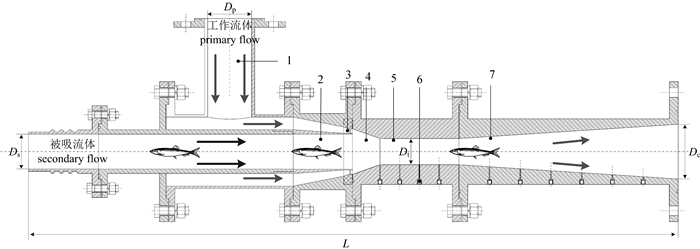
![]() 图 1 射流式鱼泵结构示意图1.工作流体输送管;2.被吸流体输送管;3.环形喷嘴;4.收缩室;5.喉管;6.测压孔;7.扩散管Fig. 1 Sketch of jet fish pump structure1. primary flow tube; 2.secondary flow tube; 3.annular nozzle; 4. suction chamber; 5. throat tube; 6. pressure measuring hole; 7. diffuser
图 1 射流式鱼泵结构示意图1.工作流体输送管;2.被吸流体输送管;3.环形喷嘴;4.收缩室;5.喉管;6.测压孔;7.扩散管Fig. 1 Sketch of jet fish pump structure1. primary flow tube; 2.secondary flow tube; 3.annular nozzle; 4. suction chamber; 5. throat tube; 6. pressure measuring hole; 7. diffuser射流式鱼泵性能具有自模性,其性能参数常常由一组无量纲参数描述(表 1)。其中面积比很重要,决定了泵的性能。
表 1 射流式鱼泵性能参数Table 1 Jet fish pump performance parameters性能参数
performance parameter面积比m
area ratio流量比q
flow ratio压力比h
pressure ratio泵效率η
pump efficiency输送能力ηf
conveyance capacity表达式 expression m=At/Aj q=Qs/Qj h=(Pc-Ps)/(Pj-Pc) η=qh ηf=mf/t 表 1中Q、P、A分别为体积流量、总压和面积;下标t、c、s、j分别表示喉管、泵出口、吸入室入口和环形喷嘴位置;mf为通过射流式鱼泵的鱼质量,t为通过时间。
该泵主要结构参数Ds、Dt、Dp、Dc和L(D表示直径,L表示泵轴向长,下标p表示工作流体入口)分别为80 mm、60 mm、100 mm、125 mm和1 480 mm。喉管直径Dt是关键尺寸,决定了过鱼尺寸的大小,可将其定义为该泵的特征尺寸。
1.2 实验材料及环境
实验用草鱼购自湖北团风养殖场,鉴于该射流式鱼泵喉管直径较小(60 mm),研究选用的实验草鱼需要同时满足输送实验(尺寸不可过大)及血液指标检测实验(尺寸不可过小),故实验鱼的体型参数选取为体质量(28.81±0.78) g,体长(126.3±1.2) mm,体宽(24.9±2.1)mm。实验用鱼通过充氧水包运到武汉大学射流式鱼泵实验台,该实验台为9.00 m×4.70 m×1.60 m(长×宽×高)的室外水泥池,池内设有过滤装置,由潜水泵驱动池内水循环,池内并排放入3个1.60 m×1.00 m×1.00 m的网箱,所有实验鱼均分养殖在3个网箱内,网箱每天24 h曝气,暂养14 d后进行实验,实验时水温(25.11±0.06)℃,pH 8.25±0.03,溶氧(8.32±0.01)mg·L-1。为了模拟实际应用情况,射流式鱼泵被吸流体输送管与锥形网管(以钢筋为骨架,外面编织无结渔网而形成的锥形的网状容器)相连以便将实验鱼限制在狭小空间内,同时出口压力通过出口管扬程控制,以避免阀门对实验鱼的损伤。
1.3 实验方法
1.3.1 射流式鱼泵草鱼输送
实验使用面积比为1.75的射流式鱼泵并设置3个工况,实验开始时,捞取7.14 kg草鱼缓慢移入射流式鱼泵入口处锥形网管内,并开始计时,待所有草鱼通过射流式鱼泵停止计时,记录通过时间,高速摄像仪(Photron FASTCAM SA5)记录输送过程,每组实验重复2次。
1.3.2 草鱼过泵后生理指标检测
实验使用面积比为1.75的射流式鱼泵并设置4个工况,过泵前为对照组,过泵后设置0 h、0.5 h、1 h、6 h、24 h共5个恢复时间点,每个时间点设置4个生物学重复。实验开始时,捞取30尾实验鱼缓慢移入射流式鱼泵入口处锥形网管内,待全部实验鱼通过泵体后,于回收箱中将实验鱼全部收集起来放入水泥池内的网箱中使鱼恢复并开始计时。依次于设定的各恢复时间点取出实验鱼,立即放入含有200 mg·L-1 MS-222(磺化甲烷三卡因)的水中麻醉,取出放置于工作台,用一次性无菌注射器(1 mL)尾动脉采血。室温静置2 h后,用转速为4 000 r·min-1离心机离心15 min,吸取血清,放置于-80 ℃密封保存。每个时间恢复点取3尾实验鱼,分置于3个透明的鱼缸内,用摄像机录像5 min,保存录像并统计其呼吸频率。呼吸频率:以鳃盖一张一合作为1次呼吸,统计15 s内各个实验鱼的呼吸次数,每间隔15 s统计1次,共统计4次,求4次统计的平均值,计算出1 min的呼吸次数,即呼吸频率(次·min-1)。检测皮质醇(COR)采用竞争性放射性免疫测定法(RIA),试剂盒购自北京北方生物技术研究所。谷草转氨酶(AST)、谷丙转氨酶(ALT)、肌酐(CREA)、总胆固醇(TC)、乳酸脱氢酶(LDH)、血糖(GLU)均在贝克曼AU5400全自动生化分析仪上检测。
1.4 数值模拟
采用ANSYS FLUENT 14.5软件对射流式鱼泵进行全尺寸数值模拟。以面积比1.75的射流式鱼泵为研究对象,建立了该泵三维模型,其几何尺寸与实验用射流式鱼泵一致,并且进、出口直管段长度均延伸各管10倍管径以确保来流均匀,其计算区域和局部网格如图 2所示。网格无关性分析表明,网格数量为454万和654万时泵的外特性和壁面压力基本一致,误差在计算允许的范围内。故文章采用网格数为454万对射流式鱼泵进行模拟计算。
数值模拟中泵内流动假设为恒定、不可压缩流动。控制方程为雷诺平均N-S方程和质量守恒方程,由于射流式鱼泵内部流动属于包含射流和混合流的管道内流动,故采用Realizable k-ε湍流模型封闭方程[30],采用无滑移标准壁面函数处理壁面应力。工作流体和被吸流体均为水,工作流体和被吸流体入口边界均给定为速度入口,出口边界设定为压力出口,并采用SIMPLEC算法求解速度和压力耦合。
2. 结果与分析
2.1 射流式鱼泵草鱼输送能力
实验中测定了不同工况下射流式鱼泵在扬程为2.24 m时的草鱼输送能力,为了衡量鱼泵所需工作流体能量大小,特计入工作流体的水功率Pw, j大小,Pw, j=Pj·Qj。实际应用中工作泵与射流式鱼泵之间管路较短,管路损失可以忽略,可认为水功率与工作泵输出功率相等。相关结果见表 2和表 3。
表 2 射流式鱼泵输送参数Table 2 Jet fish pump transport parameters组别
group工作流体流量/m3·h-1
primary flow rate流量比
flow ratio工作流体压力/kPa
primary flow pressuret/s 鱼水比
fish to water ratio1 69.81 0.33 72.85 52 1:46 2 69.81 0.33 72.85 55 1:48 3 80.20 0.42 96.20 49 1:63 4 80.20 0.42 96.20 34 1:44 5 90.51 0.45 112.48 28 1:44 6 90.51 0.45 112.48 34 1:54 表 3 射流式鱼泵输送计算Table 3 Jet fish pump transport calculation组别
group输送能力/kg·h-1
transport efficiency平均输送能力/kg·h-1
average transport efficiency工作流体的水功率/kW
water power输送单位质量鱼能耗/kW·h·t-1
transport energy consumption per unit mass of grass carp平均输送单位质量鱼能耗/kW·h·t-1
average transport energy consumption per unit mass of grass carp1 494.38 480.90 1.410 2.86 2.94 2 467.410 1.410 3.02 3 524.57 640.29 2.143 4.09 3.46 4 756.00 2.143 2.83 5 918.00 837.00 2.830 3.08 3.41 6 756.00 2.830 3.74 实验中由于草鱼移入射流式鱼泵入口处的锥形网管的过程受人工影响,而草鱼在网管内游动空间虽然受到限制,但是仍可自由游动,故在相同工况下所测量的输送时间有一定的差异,但其随流量比变化的趋势是明确的。同样,对于鱼水比的计算也受到输送时间的影响,但差异不是特别明显。
由上表,射流式鱼泵草鱼输送能力随着流量比的增加而增大,3种工况中最高输送能力达918 kg·h-1。而射流式鱼泵能耗低,每吨草鱼输送所需水功率约3 kW·h,综合效能高。另外,该泵喉管尺寸仅60 mm,若设计更大喉管尺寸的射流式鱼泵,则输送能力将进一步提升。
2.2 数值模拟结果分析
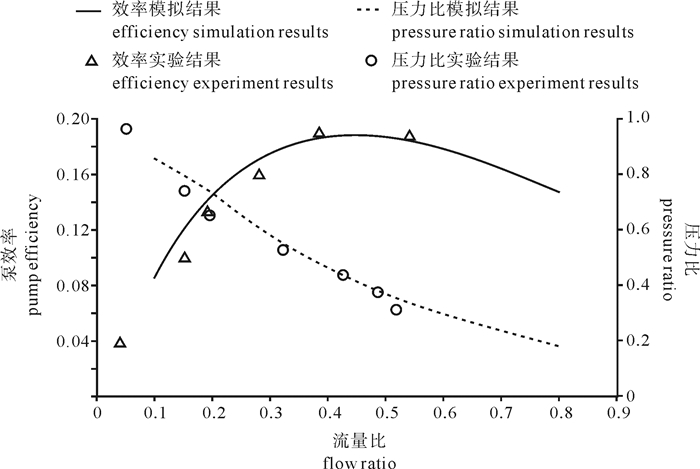
图 3为面积比1.75射流式鱼泵的压力比h和泵效率η随流量比q变化的数值模拟结果与实验数据的比较。两者吻合较好,说明了射流式鱼泵内部流场数值模拟的可靠性,可以结合实验进行进一步分析。
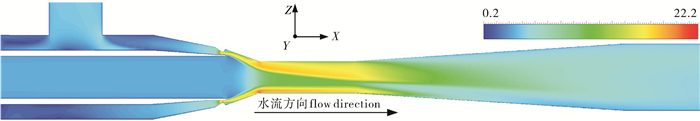
图 4为面积比1.75射流式鱼泵在流量比0.4下的速度云图,可以发现泵内流体流动并不是对称的,泵内流体在喉管及扩散管向一侧壁面偏移。这是因为泵自身结构并非完全轴对称,高压工作流体从泵的一侧进入,该不对称的泵结构导致不对称的流场分布,即内流偏转。草鱼在泵内被输送的过程中极可能受到内流偏转的影响,无法维持沿泵中心轴线方向运动,而有径向方向的运动,甚至与泵内壁面发生碰撞,以致造成草鱼损伤。
且图 4中高速工作流体与低速被吸流体在收缩室及喉管处混合时,径向方向存在着明显的速度梯度,即剪切层。若草鱼在被输送的过程中接触到剪切层,则其可能受到剪切作用的影响,导致其鳃盖和鳞片受到损伤。
由于射流式鱼泵的收缩室及扩散管结构,流体在其内部压力分布并不均匀。在收缩管内流体流速加快而压力迅速降低;喉管内流体速度达到最大值,同时压力也降至最低;而在扩散管的作用下,流体流速降低,同时压力也相应升高。因此,实验鱼在泵内经历了环境压力先降低再升高的过程。这种快速的压力变化可能对实验鱼鱼鳔和眼睛等器官造成影响,使其受到很大的负载,甚至可能失去应有的功能。
2.3 实验鱼损伤统计
实验中对过泵后的草鱼进行了外表观察,统计了呼吸频率,并对其血清进行了取样并检测其中ATL、AST、CREA、TC、GLU、COR和LDH等指标[32-35]。通过对比对照组和0 h、0.5 h、1 h、6 h、24 h组血液指标发现,相对于对照组,上述指标大部分在0 h、0.5 h和1 h组中有较为明显的变化,并在6 h和24 h组得到恢复;少数指标在0 h到6 h组内都有较为明显的变化,直到24 h组恢复。在实验中未发现实验鱼游泳异常,且通过解剖实验鱼未发现器官损伤。即射流式鱼泵的输送对草鱼的生理指标产生一定影响,但是这种影响是可恢复的,不会对草鱼内脏等器官造成损伤。
实验中共观察并统计了297尾草鱼的外部损伤情况,发现32.32%的草鱼出现了明显的鳞片脱落现象(鳞片脱落面积占总表面积的5% ~10%),5.39%的草鱼出现了严重的鳞片脱落现象(鳞片脱落面积占总表面积比例大于10%),1.01%的鱼背鳍出现碰伤,没有草鱼死亡。
3. 讨论
射流式鱼泵是鱼类输送的重要流体机械,研究中,实验泵最高活鱼输送能力达到了918 kg·h-1,水力功率仅2.83 kW,优于真空式鱼泵和气力式鱼泵,且随着射流式鱼泵结构尺寸的增大,其输送能力会有更为明显的提升;实验中没有实验鱼死亡,鱼损伤小且静养之后各项生理指标能恢复,优于离心式鱼泵。因此,相比于其他类型的鱼泵,射流式鱼泵在综合性能上有明显的优势。射流式鱼泵内部没有旋转部件,在输送过程中对鱼造成的损伤小。但是,该研究采用数值模拟的方法研究射流式鱼泵内部流动时发现,泵内流场结构复杂,存在如压力梯度、剪切层和内流偏转等特殊流动状态,仍可能给被输送鱼造成损伤。
由于射流式鱼泵的本质是一类环形射流泵,其工作原理是高速工作流体在喷嘴位置处卷吸低速被吸流体,而2种不同流速的流体在掺混的过程中存在明显的剪切层,该研究所示速度云图也明显地指示出剪切层的存在。NEITZEL等[26]通过将不同类型的幼鱼放置于速度0~21.3 m·s-1的水下射流中以研究剪切层的影响,发现在相似的环境中逆流鱼类损伤及死亡率高于顺流鱼类。而从文章的损伤结果中可以发现,并未有实验鱼死亡,但有较多的鳞片脱落现象。这是因为剪切层对鱼造成的损伤程度与其作用在鱼体表的时间有着密切的关系,而实验鱼经过剪切层的时间极短,因此损伤体现在鳞片脱落上,而不致死亡。由于泵内流体流速变化较快,实验鱼体表的鳞片很容易与其体表间产生空隙,与体表连接不紧密的鳞片在剪应力的作用下脱落,因此剪切层是造成鳞片脱落的主要原因。此外,实验鱼在暂养期间与网箱剐蹭以及实验操作过程中的人为因素都可能造成实验鱼鳞片脱落。
剪切层在射流式鱼泵内不可避免,但是,若实验鱼保持轴线运动则在更下游的位置接触到剪切层,其受到的剪应力较小。而数值模拟发现,泵内存在明显的内流偏转现象,在内流偏转的作用下,实验鱼较早地接触到了剪切层,以致鳞片损伤。由于无法实现结构完全轴对称,内流偏转现象普遍存在于各种水力机械中,而该现象却很少被研究者重视。文章通过高速摄影技术发现实验鱼在被输送的过程中受到内流偏转的影响,在泵内偏向其中一侧,以致实验鱼接触泵内壁面而导致撞击伤及鳞片脱落等现象,故内流偏转是鱼在被射流式鱼泵输送时出现撞击伤的主要原因。
鱼周围环境压力迅速降低时,鱼鳔内的气体膨胀,可能导致鱼鳔破裂,鱼自身失去平衡,同时鱼眼可能受到压力作用而突出甚至损伤[15-25]。TYTLER和BLAXTER[15]发现鱼在经过压力迅速降低后,表现出浮力上升的现象且失去平衡,通常不能再回到它们原来生活的水深。RUMMER等[36]通过X光成像技术和解剖的方法,发现红鲷在压力以10.1 kPa·s-1速率降低时,其鱼鳔出现了过度膨胀的现象。笔者发现,射流式鱼泵收缩管和喉管位置存在非常明显的压力梯度,达到了约300 kPa·s-1,远高于10.1 kPa·s-1,但对实验鱼外观观察及解剖后并未发现鱼眼突出和内脏受损等情况,这是因为实验鱼经过该位置时间非常短暂,约0.05 s,内脏等器官尚未对压力变化做出应激反应便已脱离该刺激。
在损伤结果方面,没有实验鱼在输送后死亡或表现出游泳异常,且对实验鱼解剖后也未发现组织或器官损伤,即射流式鱼泵输送鱼类的过程是非致命的。同时通过检测发现,实验鱼在过泵后其大部分血液指标与未过泵的对照组相比会有明显的变化,但这种变化在经过24 h的静养之后都可以恢复到未过泵时的水平,表明实验鱼在过泵过程中产生了应激反应,但不会出现过度应激而致死的情况,且该应激反应在静养一段时间之后可以得到恢复。根据数值模拟和高速摄影结果,实验鱼产生该应激的原因可能有以下几类:1)剪切层等强烈水力作用;2)内流偏转诱导其撞击泵内壁面;3)泵内压力梯度的刺激。
之前的研究很少对射流式鱼泵输送能力、水功率以及输送过程中鱼类损伤情况进行分析,该研究是射流式鱼泵输送草鱼性能及其损伤的探索性研究,为其他类似研究提供了参考。但是鉴于实验条件有限,可能无法全面反映不同型号尺寸的射流式鱼泵输送各种鱼类的能力以及水功率,因此,射流式鱼泵鱼类输送性能仍需要更加全面的研究。
-
![]()
图 1 射流式鱼泵结构示意图
1.工作流体输送管;2.被吸流体输送管;3.环形喷嘴;4.收缩室;5.喉管;6.测压孔;7.扩散管
Figure 1. Sketch of jet fish pump structure
1. primary flow tube; 2.secondary flow tube; 3.annular nozzle; 4. suction chamber; 5. throat tube; 6. pressure measuring hole; 7. diffuser
表 1 射流式鱼泵性能参数
Table 1 Jet fish pump performance parameters
性能参数
performance parameter面积比m
area ratio流量比q
flow ratio压力比h
pressure ratio泵效率η
pump efficiency输送能力ηf
conveyance capacity表达式 expression m=At/Aj q=Qs/Qj h=(Pc-Ps)/(Pj-Pc) η=qh ηf=mf/t  下载: 导出CSV
下载: 导出CSV
表 2 射流式鱼泵输送参数
Table 2 Jet fish pump transport parameters
组别
group工作流体流量/m3·h-1
primary flow rate流量比
flow ratio工作流体压力/kPa
primary flow pressuret/s 鱼水比
fish to water ratio1 69.81 0.33 72.85 52 1:46 2 69.81 0.33 72.85 55 1:48 3 80.20 0.42 96.20 49 1:63 4 80.20 0.42 96.20 34 1:44 5 90.51 0.45 112.48 28 1:44 6 90.51 0.45 112.48 34 1:54
下载: 导出CSV
表 3 射流式鱼泵输送计算
Table 3 Jet fish pump transport calculation
组别
group输送能力/kg·h-1
transport efficiency平均输送能力/kg·h-1
average transport efficiency工作流体的水功率/kW
water power输送单位质量鱼能耗/kW·h·t-1
transport energy consumption per unit mass of grass carp平均输送单位质量鱼能耗/kW·h·t-1
average transport energy consumption per unit mass of grass carp1 494.38 480.90 1.410 2.86 2.94 2 467.410 1.410 3.02 3 524.57 640.29 2.143 4.09 3.46 4 756.00 2.143 2.83 5 918.00 837.00 2.830 3.08 3.41 6 756.00 2.830 3.74
下载: 导出CSV
-
[1] SUMMERFELT S T, DAVIDSON J, WILSON G, et al. Advances in fish harvest technologies for circular tanks [J].Aquacultl Engin, 2009, 40:62-71. doi: 10.1016/j.aquaeng.2008.12.001
[2] VALDEMARSEN J W. Technological trends in capture fisheries [J].Ocean & Coastal Manag, 2001, 44:635-651. https://www.sciencedirect.com/science/article/pii/S0964569101000734
[3] 刘健, 钱晨荣, 黄洪亮.国内外吸鱼泵研究进展[J].渔业现代化, 2013, 40(1):57-62. http://www.cqvip.com/QK/90617A/201301/45032673.html [4] 许钟, 曲敏.吸鱼泵在日本的应用[J].渔业机械仪器, 1996, 3:31-33. http://kns.cnki.net/KCMS/detail/detail.aspx?filename=hdxy199603011&dbname=CJFD&dbcode=CJFQ [5] 叶燮明, 徐君卓.国内外吸鱼泵研制现状[J].现代渔业信息, 2005, 20(9):7-8. http://www.cqvip.com/QK/90549X/200509/20209698.html [6] 徐英士.真空式吸鱼泵阀件设计[J].渔业现代化, 2007, 6:39. http://www.cnki.com.cn/Article/CJFDTOTAL-CANB201304015.htm [7] 袁军亭, 汤威, 刘忠斌, 等.排水辅助减压型真空式鱼泵研究[J].上海水产大学学报, 2007, 16(1):54-59. http://kns.cnki.net/KCMS/detail/detail.aspx?filename=ssdb200701010&dbname=CJFD&dbcode=CJFQ [8] 董又新.鱼泵及其分类[J].河北渔业, 2005(3):16. http://kns.cnki.net/KCMS/detail/detail.aspx?filename=hbyu199303004&dbname=CJFD&dbcode=CJFQ [9] LONG X P, XU M S, LYU Q, et al.Impact of the internal flow in a jet fish pump on the fish[J].Ocean Engin, 2016, 126:313-320 doi: 10.1016/j.oceaneng.2016.09.027
[10] 郭喜庚, 谭细畅.吸鱼泵的主要结构型式及其应用前景分析[J].渔业信息与战略, 2013, 28(3):214-218. http://d.wanfangdata.com.cn/Periodical/xdyyxx201303007 [11] 黄滨, 关长涛, 林德芳.网箱真空吸鱼泵试验中的技术问题研究[J].渔业现代化, 2005, 32(6):39-41. http://www.doc88.com/p-473422330461.html [12] 黄小华, 郭根喜, 陶启友.射流式吸鱼泵关键技术研究及设计[J].南方水产科学, 2007, 3(3):41-46. http://www.schinafish.cn/CN/abstract/abstract8376.shtml [13] 叶燮明.文丘里吸鱼泵研究[J].渔业现代化, 2006, 33(4):37-38. http://www.wenkuxiazai.com/doc/2505468b6bec0975f465e263.html [14] XIAO L Z, LONG X P, LI L, et al. Movement characteristics of fish in a jet fish pump [J].Ocean Engin, 2015, 108:480-492. doi: 10.1016/j.oceaneng.2015.08.039
[15] TYTLER P, BLAXTER J. The effect of swimbladder deflation on pressure sensitivity in the saithe Pollachius virens [J].J Mar Biol Assoc UK, 1977, 57:1057-1064. doi: 10.1017/S0025315400026126
[16] 赵希坤, 薛桢锷.鱼类克服流速能力的试验[J].水产学报, 1980, 4(1):31-37. http://www.cqvip.com/QK/90183X/198001/15118995.html [17] PICKERING A. Environmental stress and the survival of brown trout Salmo trutta[J].Freshw Biol, 1989, 21:47-55. doi: 10.1111/fwb.1989.21.issue-1
[18] CADA G F. A review of studies relating to the effects of propeller-type turbine passage on fish early life stages [J].N Am J Fish Manag, 1990, 10:418-426. doi: 10.1577/1548-8675(1990)010<0418:AROSRT>2.3.CO;2
[19] PARRISH F A, MOFFITT R B. Subsurface fish handling to limit decompression effects on deepwater species [J].Mar Fish Rev, 1992, 54:29-32. http://aquaticcommons.org/9893/
[20] CADA G F. The development of advanced hydroelectric turbines to improve fish passage survival [J].Fisheries, 2001, 26:14-23.
[21] GARRISON L A, FISHER J R K, SALE M J, et al. Application of biological design criteria and computational fluid dynamics to investigate fish survival in Kaplan turbines [J].Proc Hydro Vision, 2002:1-12. https://www.researchgate.net/publication/239589108_Application_of...
[22] JARVIS E T, LOWE C G. The effects of barotrauma on the catch-and-release survival of southern California nearshore and shelf rockfish (Scorpaenidae, Sebastes spp.) [J].Can J Fish Aquat Sci, 2008, 65:1286-1296. doi: 10.1139/F08-071
[23] BROWN R S, CARLSON T J, WELCH A E, et al. Assessment of barotrauma from rapid decompression of depth-acclimated juvenile Chinook salmon bearing radiotelemetry transmitters [J].Trans Am Fish Soc, 2009, 138:1285-1301. doi: 10.1577/T08-122.1
[24] RICHMOND M C, SERKOWSKI J A, EBONER L L, et al. Quantifying barotrauma risk to juvenile fish during hydro-turbine passage [J].Fish Res, 2014, 154:152-164. doi: 10.1016/j.fishres.2014.01.007
[25] TRUMBO B A, AHMANN M L, RENHLODS J F, et al. Improving hydroturbine pressures to enhance salmon passage survival and recovery [J].Rev Fish Biol Fish, 2014, 24:955-965. doi: 10.1007/s11160-013-9340-8
[26] NEITZEL D A, DAUBLE D D, CADA G F, et al. Survival estimates for juvenile fish subjected to a laboratory-generated shear environment [J].Trans Am Fish Soc, 2004, 133:447-454. doi: 10.1577/02-021
[27] 刘平.基于ANSYS CFX的吸鱼泵的内部流场分析[J].流体机械, 2014, 42(11):43-46. doi: 10.3969/j.issn.1005-0329.2014.11.009 [28] LONG X P, ZENG Q L, YANG X L, et al. Structure optimization of an annular jet pump using design of experiment method and CFD[C].26th IAHR Symposiumon Hydraulic Machinery and Systems, 2012:1-7. http://adsabs.harvard.edu/abs/2012E%26ES...15e2020L
[29] 曾庆龙, 龙新平, 肖龙洲, 等.环形射流泵结构优化设计[J].排灌机械工程学报, 2014, 32(2):98-102. doi: 10.3969/j.issn.1674-8530.12.1217 [30] 杨雪龙, 龙新平, 肖龙洲, 等.不同湍流模型对射流泵内部流场模拟的影响[J].排灌机械工程学报, 2013, 31(2):98-102. http://www.wenkuxiazai.com/doc/3a2e9b6067ec102de2bd89cd.html [31] 涂志英. 雅砻江流域典型鱼类游泳特性研究[D]. 武汉: 武汉大学, 2012: 3. [32] CHAN D K, WOO N Y. Effect of cortisol on the metabolism of the eel, Anguilla japonica[J].Gen Comp Endocrinol, 1978, 35:205-215. doi: 10.1016/0016-6480(78)90064-3
[33] MOMMSEN T P, VIJAYAN M M, MOON T W. Cortisol in teleosts:dynamics, mechanisms of action, and metabolic regulation [J].Rev Fish Biol Fish, 1999, 9:211-268. doi: 10.1023/A:1008924418720
[34] 王波, 李继强, 曹志海, 等.犬齿牙鲆幼鱼呼吸耗氧率的研究[J].海洋科学进展, 2003, 21:325-330. doi: 10.3969/j.issn.1671-6647.2003.03.011 [35] 李黎. 力竭性运动后鲇鱼幼鱼生理生化指标的变动[D]. 重庆: 重庆师范大学, 2006: 2. [36] RUMMER J L, BENNETT W A. Physiological effects of swim bladder overexpansion and catastrophic decompression on red snapper [J].Trans Am Fish Soc, 2005, 134(6):1457-1470. doi: 10.1577/T04-235.1
-
期刊类型引用(2)
1. 叶梓茵,欧慧慧,丁月霞. 罗非鱼无乳链球菌毒力检测及药敏试验. 中国兽医杂志. 2021(07): 47-51 .  百度学术
百度学术
2. 李战军,张建柏,曹亚男,陈坤,刘蓬,徐惠章,王鹤. 鱼类副乳房链球菌病研究进展. 大连海洋大学学报. 2021(06): 1059-1070 . 百度学术
其他类型引用(5)


计量
- 文章访问数: 3304
- HTML全文浏览量: 991
- PDF下载量: 35
- 被引次数: 7
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
