
Ecological basis assessment of artificial reef site selection in Liuniu coastal waters of Zhelin Bay, Eastern Guangdong
-
摘要:
采用层次分析法,构建了基于生态系统效应的资源保护型人工鱼礁建设选址评价方法,通过选择水文、水质、底质和浮游生物、鱼卵仔鱼、大型底栖动物、游泳动物、初级生产力、潮间带大型底栖动物等30个海洋环境与海洋生物因子,建立了人工鱼礁建设选址生态评估模型;以柘林湾溜牛人工鱼礁区选址区域为例,对所选因子进行了春、夏、秋、冬4个季度的跟踪监测,探讨了该海区进行人工鱼礁建设的适宜性,根据该模型计算得出各季度人工鱼礁建设选址生态基础评价指数分别为0.65、0.71、0.63、0.64,平均为0.66,计算结果表明,该区域建礁生态基础良好,适宜选址开展人工鱼礁建设,其中生态基础最优的季节为夏季。
Abstract:The AHP was employed to evaluate the stock protection artificial reef site selection based on ecosystem effect. We selected 30 marine environment and marine biological factors (hydrology, water quality, sediment, macrobenthos, swimming animals, primary productivity, intertidal macrobenthos, etc.) to establish an ecological evaluation model for artificial reef site selection. This paper tracked and monitored the selected factors at artificial reef site in the Liuniu coastal waters of Zhelin Bay in spring, summer, autumn and winter. The suitability of this area for building artificial reefs was discussed. According to the calculation results by this model, indices of ecological basis assessment for artificial reef site selection in each season were 0.65, 0.71, 0.63 and 0.64 with an average value of 0.66. The results show that there is a good ecological foundation for building artificial reef, and it is a suitable site to construct artificial reefs. The optimal ecological foundation is in summer.
-
Keywords:
- artificial reef /
- site selection /
- ecological basis assessment /
- Zhelin Bay
-
生长是水产养殖经济动物的重要性状。在一些经济鱼类中,肌肉占到鱼体质量的50%~70%[1]。肌球蛋白是肌肉细胞的重要组成部分,在肌肉收缩、细胞分裂及细胞内物质运输等方面有着重要功能[2-3]。肌球蛋白由2条肌球蛋白重链 (Myosin heavy chain) 和4条肌球蛋白轻链 (Myosin light chain) 共同形成“Y”形不对称结构。肌球蛋白重链具有运动和肌纤维形成双重生物学功能,在肌肉生长、发育和收缩上扮演着重要角色[4],这与其球状氨基末端头部结构域和细长α-螺旋卷曲羧基末端棒状结构域密不可分[5]。肌球蛋白重链基因属多基因家族,在鳜 (Siniperca chuatsi)[6]、青鳉 (Oryzias latipes)[7]、斑马鱼 (Danio rerio)[8]、红鳍东方鲀 (Takifugu rubripers)[9]等多种鱼类中发现了不同基因类型。该基因编码区较长,普遍在5 000 bp以上,约可编码1 900个氨基酸。
肌球蛋白重链基因可通过表达量的差异以及核苷酸序列上存在的单核苷酸多态性 (Single nucleotide polymorphisms, SNPs) 位点影响物种生长[10-13]。已有文献报道其表达量与石斑鱼仔鱼期肌纤维的新生、仔鱼后期原有肌纤维的增生相关[14]。在红鳍东方鲀中,肌球蛋白重链基因表达量的差异体现在快慢肌中,影响其生长速率[15]。在虹鳟 (Oncorhynchus mykiss) 和玻璃梭吻鲈 (Sander vitreus) 中也同样发现肌球蛋白重链基因的表达量会影响生长速度[16-17]。此外,在鳜中发现肌球蛋白重链基因5' 转录调控区存在与生长表型性状 (如体长、体高等) 相关的SNPs位点,这些位点位于启动子区域,可通过影响转录调控因子结合进而改变该基因的表达量,最终实现对生长性状的调控[4]。
石斑鱼为名贵水产经济动物,属鲈形目、鮨科、石斑鱼亚科。因其低脂肪、高蛋白、肉质细腻等特点,备受消费者喜爱。在众多石斑鱼养殖种类中,杉虎杂交斑 (棕点石斑鱼Epinephelus fuscoguttatus ♀ × 清水石斑鱼E. polyphekadion ♂) 因生长速度快、抗病性强、肉质鲜嫩等特点,成为我国沿海地区重要的养殖鱼类[18-19]。有学者基于生长重要基因,如生长激素 (Growth hormone)[20]、生长激素受体 (Growth hormone receptors)[19]、肌肉抑制素 (Myostatin)[21]来研究杉虎杂交斑的生长性状。本研究团队在杉虎杂交斑及其亲本的转录组数据比较分析中发现,肌球蛋白重链基因在杉虎杂交斑生长性状调控中具有重要作用。但该基因在杉虎杂交斑中尚未见相关报道。获取杉虎杂交斑肌球蛋白重链基因的序列全长,分析其蛋白结构是解析该基因功能的重要前提。
分子标记辅助育种借助于目标性状分子标记对个体进行选择,进而获得含有目标基因的优良个体。该技术在水产经济动物育种中应用较为成熟,其在育种中呈现的高效性和精准性是单一依赖表型性状进行传统选育所无法比拟的[22-23]。筛选与性状紧密相关的分子标记是开展辅助育种的重要前提和基础。基于生长重要调控基因的SNPs位点挖掘生长分子标记是一种非常有效的手段,在植物和动物育种中应用较为广泛[24-26]。在水产经济动物如鳜[6]、大口黑鲈 (Micropterus salmoides)[13]、马氏珠母贝 (Pinctada fucata martensii)[12]中,均在肌球蛋白重链基因核苷酸序列中成功筛选到与生长相关的SNPs位点。因此,本研究克隆了杉虎杂交斑的肌球蛋白重链基因核苷酸序列,预测其蛋白结构,并挖掘与生长相关的SNPs位点,可为杉虎杂交斑生长分子标记开发提供候选位点,为筛选鉴定具有生长优势的杉虎杂交斑提供技术支撑。
1. 材料与方法
1.1 实验材料
杉虎杂交斑采集自陵水德林诚信水产养殖有限公司 (海南省陵水县)。杉虎杂交斑在鱼苗时期养殖在4 m×4 m×1.2 m的椭圆形水泥池中,养殖密度为1 200尾·m−3。根据鱼苗生长情况逐渐调整养殖密度,最后转移至6 m×5 m×1.5 m的长方形水泥池进行养殖。至12月龄时其养殖密度约为50尾·m−3,每日投喂1~2次。利用游标卡尺测量养殖在同一水泥池中12月龄杉虎杂交斑表型生长数据 (包括全长、体长、头长、体高和尾柄高)。利用电子秤称量体质量,精确到0.01 kg。在体质量数据符合正态分布的群体中,挑选位于两端极值的杉虎杂交斑个体 (即生长较快个体和生长较慢个体) 组成2个群体,每个群体各30尾。生长较快的群体平均体质量为 (0.59±0.05) kg,生长较慢的群体平均体质量为 (0.15±0.03) kg。表型数据测定前利用3-氨基苯甲酸乙酯甲基磺酸盐 (MS-222) 对鱼体进行麻醉,数据测量同时取尾鳍组织。按照动物基因组 DNA 快速抽提试剂盒 [生工生物工程 (上海) 有限公司] 的操作步骤提取基因组DNA。本研究操作严格遵守实验动物伦理要求。
1.2 肌球蛋白重链基因全长序列扩增及SNPs位点筛选
根据研究团队测得的棕点石斑鱼基因组,获得肌球蛋白重链基因参考序列并进行引物设计 (表1)。以杉虎杂交斑DNA为模板,通过PCR扩增获取肌球蛋白重链基因序列全长。扩增体系为:DNA模板1 μL,Taq 5 μL,正、反向引物各0.5 μL,最后用水补齐至10 μL。扩增程序:55~60 °C退火30 s,72 °C延伸30 s,循环数为30。将扩增获得的产物送至广州生工进行测序。采用BioEdit和Clustal W对序列进行多重比对分析并筛选SNPs位点。根据SNPs位点前后150 bp序列进行引物设计,再次进行PCR扩增以验证SNPs位点 (表1)。
表 1 用于PCR扩增引物表Table 1 Primers used for PCR引物名称
Primer序列 (5'—3')
Sequence (5'−3')用途
Usage引物1
Primer 1F: GAACTTTTTATCATCTCTAC 全长序列扩增 R: ACTTGTTATTTTCTTTTTGT 引物2
Primer 2F: GTATCTAAAACATGAAGCAC 全长序列扩增 R: AACCTACAAATTATCGACCA 引物3
Primer 3F: GGTCCCAAATCAAACTCCTG 全长序列扩增 R: GCCTGACATCTGTTGCCATC 引物4
Primer 4F: TGTCAGGCTCTCACTCATTC 全长序列扩增 R: GTAAGTGTCTCTTCTGATTG 引物5
Primer 5F: TACTCAAGAGGAAAAGAACT 全长序列扩增 R: TTGTTCTTCACCAGCCAGTT 引物6
Primer 6F: AGCTTCTGATACCACCTTTA 全长序列扩增 R: ACAAATCACAAACCAATCAT 引物7
Primer 7F: CTATTCAAGCCAGGTCCCGT 全长序列扩增 R: CATCCTCAAGTCTGCCATTT 引物 8
Primer 8F: GACCTTGAGAGAGCTAAGAG 全长序列扩增 R: CAAGTGAAAAGAGTAATGCT 引物 9
Primer 9F: GTCAGCATTACTCTTTTCAC 全长序列扩增 R: TCTCTCTCTTCAGGGTCTCC 引物 10
Primer 10F: GGAAACAAAAATATGAGGAG 全长序列扩增 R: ATACAATGTACGAAAAAGCA 引物 11
Primer 11F: CAGATTGAAGTAGAGGAAGC 全长序列扩增 R: GGGAAAGAAAGAAGAAGAGA 引物12
Primer 12F: AGAACCAATCAGAAGAGACA SNP位点验证 R: TGTAATACCAAGCAGTGAAA 引物13
Primer 13F: ACTTTCATAGATTTTGGGAT SNP位点验证 R: TAGACTTCTGGTAGAGCGCA 引物14
Primer 14F: ACTCATATTTTTGTCCCCTC SNP位点验证 R: TTTGCCTTCAGTAACTTCGG 引物15
Primer 15F: ATAAAATATTCTGTCCCTCC SNP位点验证 R: TGTCTTGAAAGTTCATCTAG 引物16
Primer 16F: ATTATCATTTGCCAAAATGT SNP位点验证 R: TCCTTCACCAAGTTGCTCAG 引物17
Primer 17F: TGGAGCAGGAAAAGAGCGAG SNP位点验证 R: GGTTACAAAGATGCCAGAGT 引物18
Primer 18F: CCTGAAGGTAAAAAAGAGGA SNP位点验证 R: ACAATGTACGAAAAAGCAAC 1.3 肌球蛋白重链基因序列分析
利用Open Reading Frame (ORF) (https://www.ncbi.nlm.nih.gov/orffinder/) 预测肌球蛋白重链基因序列阅读框。通过SMART (https://smart.embl.de/) 预测蛋白结构域。MEGA 7.0软件用于进化分析。SWISS-MODEL (SWISS-MODEL Interactive Workspace (expasy.org) 用于三级蛋白结构预测。
1.4 肌球蛋白重链基因SNPs分析
计算SNPs位点基因频率和基因型频率[27]。选取最小等位基因频率大于0.05的位点用于后续分析。利用PLINK 软件计算预期杂合度 (Expected heterozygosity, He)、观测杂合度 (Observed heterozygosity, Ho) 和Hardy-Weinberg平衡。采用Popgene v. 1.31[28]分析有效等位基因数 (Effective number of alleles, Ne)。多态性信息 (Polymorphic information content, PIC) 参考Bostein等[29]提出的方法进行计算。SNPs位点间的连锁不平衡分析通过Haploview 4.2计算,连锁程度大小用r2度量[30]。采用PLINK一般线性模型进行SNPs位点与生长性状关联分析。同一位点不同基因型之间的性状表现差异由SPSS 17.0软件的t-test检测。显著性水平α设为0.05。
2. 结果
2.1 肌球蛋白重链基因序列分析
经测定发现杉虎杂交斑肌球蛋白重链基因序列长度为12 129 bp,包含36个外显子和35个内含子 (图1),完整的基因序列已经提交到NCBI (PQ826489)。外显子长度介于28~390 bp,可编码1 934个氨基酸。推测其蛋白分子质量约为223 kD,等电点为5.532。蛋白结构域分析结果显示存在4种结构类型,即Myosin_N (第34—第73位氨基酸)、MYSc (第79—第782位氨基酸)、IQ (第783—第805位氨基酸) 和Myosin_tail_1 (第846—第
1927 位氨基酸)。2.2 肌球蛋白重链基因蛋白高级结构预测
将杉虎杂交斑肌球蛋白重链基因氨基酸序列与棕点石斑鱼 (XP_049453941.1)、鞍带石斑鱼 (E. lanceolatus, XP_033506503.1)、金钱鱼 (Scatophagus argus, XP_046263685.1)、红鳍东方鲀 (XP_011614688.1)、黄鳍棘鲷 (Acanthopagrus latus, XP_036941235.1)、狭鳞庸鲽 (Hippoglossus stenolepis, XP_035027113.1)、射水鱼 (Toxotes jaculatrix, XP_040910911.1) 以及大口黑鲈 (XP_038574763.1) 进行同源性比对。结果表明,杉虎杂交斑肌球蛋白重链基因氨基酸序列与其他鱼类的相似度均高于90%;其中与棕点石斑鱼和鞍带石斑鱼的相似度最高,分别为98.9%和98.7%;与狭鳞庸鲽的相似度最低 (91.6%)。以上结果表明肌球蛋白重链基因在不同物种间具有较高的保守性 (表2)。
表 2 肌球蛋白重链基因氨基酸序列在不同物种间的相似度比较Table 2 Similarity comparison of amino acid sequence in MYH gene between different species% 棕点石斑鱼
E. fuscoguttatus鞍带石斑鱼
E. lanceolatus金钱鱼
S. argus红鳍东方鲀
T. rubripes黄鳍棘鲷
A. latus狭鳞庸鲽
H. stenolepis射水鱼
T. jaculatrix大口黑鲈
M. salmoides杉虎杂交斑
E. fuscoguttatus × E. polyphekadion98.9 98.7 94.5 92.4 94.3 91.6 94.2 94.0 棕点石斑鱼
E. fuscoguttatus99.4 95.1 93.0 95.0 92.2 94.8 94.6 鞍带石斑鱼
E. lanceolatus95.0 92.9 94.8 92.2 94.9 94.5 金钱鱼
S. argus94.7 96.7 93.4 96.3 95.9 红鳍东方鲀
T. rubripes94.9 91.8 93.8 93.6 黄鳍棘鲷
A. latus93.1 95.9 96.1 狭鳞庸鲽
H. stenolepis93.0 92.3 射水鱼
T. jaculatrix95.2 对上述物种的Myosin_N、MYSc、IQ和Myosin_tail_1蛋白结构域比较分析发现,在不同物种间,结构域有较高的氨基酸序列相似性,其中IQ区域保守性最高。此外,在Myosin_N、MYSc和Myosin_tail_1这3个区域中存在石斑鱼类保守性氨基酸位点,如3种石斑鱼在Myosin_N区域第37位均为缬氨酸、MYSc区域第245位均为天冬酰胺。而这些位点与其他鱼类的氨基酸组成不同 (图2,红色星号所示)。在MYSc和Myosin_tail_1区域还存在杉虎杂交斑特异性氨基酸位点,如MYSc区域第187—第189位分别为苏氨酸、脯氨酸和色氨酸,Myosin_tail_1区域第951位为脯氨酸。这些位点与其他鱼类均不同 (图2,黑色星号所示)。
![]() 图 2 不同物种中肌球蛋白重链基因蛋白结构域氨基酸序列比较分析Fig. 2 Amino acid sequence alignment in Myosin_N, MYSc, IQ and Myosin_tail_1 domain of MYH gene among different species
图 2 不同物种中肌球蛋白重链基因蛋白结构域氨基酸序列比较分析Fig. 2 Amino acid sequence alignment in Myosin_N, MYSc, IQ and Myosin_tail_1 domain of MYH gene among different species对杉虎杂交斑、棕点石斑鱼及狭鳞庸鲽的高级蛋白结构进行预测,发现三者蛋白结构具有极高的相似性 (图3),表明肌球蛋白重链基因在不同物种间具有相似的蛋白功能。
![]() 图 3 预测肌球蛋白重链基因的蛋白结构注:a. 杉虎杂交斑;b. 棕点石斑鱼;c. 狭鳞庸鲽。Fig. 3 Predicted protein structure of MYHNote: a. E. fuscoguttatus×E. polyphekadion; b. E. fuscoguttatus;c. H. stenolepis.
图 3 预测肌球蛋白重链基因的蛋白结构注:a. 杉虎杂交斑;b. 棕点石斑鱼;c. 狭鳞庸鲽。Fig. 3 Predicted protein structure of MYHNote: a. E. fuscoguttatus×E. polyphekadion; b. E. fuscoguttatus;c. H. stenolepis.2.3 肌球蛋白重链基因遗传进化分析
基于肌球蛋白重链基因氨基酸序列构建进化树 (图4)。结果表明,3种石斑鱼先紧密聚集形成一小支,再与其他鲈形目鱼类聚集。红鳍东方鲀 (鲀形目) 相较于狭鳞庸鲽 (鲽形目),与鲈形目鱼类的遗传关系更近。
![]() 图 4 在不同物种间基于邻接法构建肌球蛋白重链基因进化树Fig. 4 Phylogenetic tree of MYH among different species based on neighbor-joining method
图 4 在不同物种间基于邻接法构建肌球蛋白重链基因进化树Fig. 4 Phylogenetic tree of MYH among different species based on neighbor-joining method2.4 肌球蛋白重链基因SNPs位点分析
根据肌球蛋白重链基因全长,筛选获得62个SNPs位点 (表3)。各位点He介于0.066 7~0.750 0,Ho介于0.152 8~0.497 8。在62个SNPs位点中,除4个为低水平遗传多态性外,其余均属于中等水平遗传多态性,未发现有位点属于高水平遗传多态性。有8个SNPs位点符合Hardy-Weinberg平衡 (p>0.05),即g960 C>T,g988 G>A,g1442 C>T,g3740 A>G,g5196 T>C,g5253 C>A,g8598 A>G,g10120 C>T。对这些位点进行连锁不平衡分析,发现位点g5196 T>C和g8598 A>G之间显示出强连锁 (r2=0.81)。
表 3 肌球蛋白重链基因上62个SNPs的遗传参数Table 3 Genetic parameters of 62 SNPs in MYH单核苷酸多态性位点
SNPs碱基类型
Type of base有效等位
基因数Ne期望杂合度
He观测杂合度
Ho遗传多态性
PICHardy-Weinberg平衡 (p值)
HWE p valueg960 T/C 1.342 3 0.300 0 0.255 0 0.255 0 0.332 4 g979 A/T 1.973 1 0.750 0 0.468 8 0.469 0 9.27×10−7* g988 A/G 1.280 0 0.250 0 0.218 8 0.219 0 0.579 5 g1011 T/C 1.882 4 0.750 0 0.468 8 0.469 0 9.27×10−7* g1017 C/T 1.800 0 0.666 7 0.444 4 0.444 0 6.59×10−5* g1027 A/G 1.800 0 0.666 7 0.444 4 0.444 0 6.59×10−5* g1038 A/G 1.800 0 0.666 7 0.444 4 0.444 0 6.59×10−5* g1113 T/A 1.935 0 0.316 7 0.483 2 0.483 0 0.008 008* g1166 T/C 1.724 1 0.550 0 0.398 8 0.399 0 0.002 727* g1187 C/T 1.724 1 0.600 0 0.420 0 0.420 0 0.0005 355*g1283 C/A 1.851 4 0.650 0 0.459 9 0.459 8 0.001 751* g1352 A/G 1.910 3 0.683 3 0.476 5 0.476 5 0.001 054* g1442 T/C 1.219 5 0.200 0 0.180 0 0.180 0 1 g1805 C/T 1.882 4 0.750 0 0.468 8 0.468 8 9.27×10−7* g2796 C/T 1.763 0 0.200 0 0.432 8 0.432 8 4.39×10−5* g2975 G/A 1.896 7 0.300 0 0.472 8 0.472 8 0.005796 *g3006 A/C 1.991 2 0.200 0 0.497 8 0.497 8 2.38×10−6* g3007 A/G 1.763 0 0.166 7 0.432 8 0.432 8 2.80×10−6* g3015 C/T 1.663 2 0.116 7 0.398 8 0.398 8 1.16×10−7* g3073 T/C 1.882 4 0.183 3 0.468 8 0.468 8 2.95×10−6* g3176 A/T 1.882 4 0.183 3 0.468 8 0.468 8 2.95×10−6* g3428 G/C 1.834 9 0.133 3 0.455 0 0.455 0 3.49×10−8* g3740 G/A 1.406 0 0.250 0 0.288 8 0.288 8 0.36 g4301 G/A 1.882 4 0.750 0 0.468 8 0.468 8 9.27×10−7* g4482 G/A 1.882 4 0.750 0 0.468 8 0.468 8 9.27×10−7* g4778 T/C 1.882 4 0.750 0 0.468 8 0.468 8 9.27×10−7* g5196 C/T 1.973 1 0.616 7 0.493 2 0.493 2 0.071 36 g5217 T/C 1.743 8 0.616 7 0.426 5 0.426 5 0.000 456 2* g5253 A/C 1.683 8 0.300 0 0.406 1 0.406 1 0.054 04 g5265 C/T 1.743 8 0.616 7 0.426 5 0.426 5 0.000 456 2* g5322 C/T 1.867 2 0.733 3 0.464 4 0.464 5 2.56×10−6* g5391 G/A 1.867 2 0.733 3 0.464 4 0.464 5 2.56×10−6* g5417 G/C 1.882 4 0.750 0 0.468 8 0.468 8 9.27×10−7* g6413 T/C 1.956 0 0.183 3 0.488 8 0.488 8 9.08×10−7* g7071 C/T 1.683 0 0.066 7 0.406 1 0.406 1 1.36×10−7* g7588 G/T 1.743 8 0.616 7 0.426 5 0.426 5 0.000 456 2* g7606 T/C 1.743 8 0.616 7 0.426 5 0.426 5 0.000 456 2* g7619 T/C 1.743 8 0.616 7 0.426 5 0.426 5 0.000 456 2* g8320 T/C 1.743 8 0.616 7 0.426 5 0.426 5 0.000 456 2* g8328 G/A 1.743 8 0.616 7 0.426 5 0.426 5 0.000 456 2* g8406 C/A 1.743 8 0.616 7 0.426 5 0.426 5 0.000 456 2* g8528 A/G 1.743 8 0.616 7 0.426 5 0.426 5 0.000 456 2* g8548 A/G 1.743 8 0.616 7 0.426 5 0.426 5 0.000 456 2* g8598 G/A 1.973 1 0.583 3 0.476 5 0.476 5 0.110 2 g8715 T/C 1.683 8 0.066 7 0.406 1 0.406 1 1.36×10−10* g9646 T/C 1.882 4 0.750 0 0.468 8 0.468 8 9.27×10−7* g9649 T/C 1.882 4 0.750 0 0.468 8 0.468 8 9.27×10−7* g9657 A/G 1.882 4 0.750 0 0.468 8 0.468 8 9.27×10−7* g9846 A/G 1.882 4 0.750 0 0.468 8 0.468 8 9.27×10−7* g9884 C/T 1.882 4 0.750 0 0.468 8 0.468 8 9.27×10−7* g9902 T/C 1.882 4 0.750 0 0.468 8 0.468 8 9.27×10−7* g10120 T/C 1.180 3 0.167 0 0.152 8 0.152 8 1 g10187 C/G 1.882 4 0.750 0 0.468 8 0.468 8 9.27×10−7* g10221 G/A 1.882 4 0.750 0 0.468 8 0.468 8 9.27×10−7* g10368 C/T 1.882 4 0.750 0 0.468 8 0.468 8 9.27×10−7* g10444 C/T 1.896 7 0.733 3 0.472 8 0.472 8 1.88×10−5* g10545 T/C 1.867 2 0.733 3 0.464 4 0.464 5 2.56×10−6* g10559 T/G 1.882 4 0.750 0 0.468 8 0.468 8 9.27×10−7* g10575 T/G 1.882 4 0.750 0 0.468 8 0.468 8 9.27×10−7* g10658 A/C 1.882 4 0.750 0 0.468 8 0.468 8 9.27×10−7* g10661 C/T 1.239 5 0.083 3 0.193 2 0.193 1 0.000 693 7* g10994 A/G 1.600 0 0.066 7 0.375 0 0.375 0 8.73×10−10* 注:*表示偏离Hardy-Weinberg定律 (p<0.05)。 Note: *. Devating from Hardy-Weinberg equilibrium (p<0.05). 2.5 SNPs与生长性状关联分析
在62个SNPs位点中有12个位点位于外显子区域,将它们与生长性状进行关联分析。结果表明,所有SNPs均与生长性状表现出显著相关 (p<0.05) (表4)。在12个SNPs中,除g5196 T>C、g5253 C>A和g8598 A>G外,其余位点均发生了基因型丢失现象。例如在g4482 A>G位点处丢失GG型。但所有位于外显子区域的SNPs仅有g5417 C>G为错义突变 (第606个氨基酸),导致丙氨酸变成甘氨酸;其余均属同义突变。位点g10444 T>C虽存在3种基因型,但CC型个体仅1个 (体质量0.16 kg,全长16.50 cm,体长14.40 cm,头长5.30 cm,体高4.27 cm,尾柄高1.40 cm)。通过对12个SNPs位点各基因型生长性状的比较分析发现,杂合基因型均不属于优势基因型。
表 4 肌球蛋白重链基因SNPs位点与各生长性状相关性Table 4 Correlations between different genotypes at SNP loci of MYH and various growth traits单核苷酸
多态性位
点SNPs位置
Location基因型
Genotype样本数
Number体质量
Body
mass/kg全长
Total
length/cm体长
Body
length/cm头长
Head
length/cm体高
Body
height/cm尾柄高
Caudal peduncle
height/cmg4482 外显子11 AA 15 0.58±0.06a 32.48±1.00a 27.30±0.88a 10.31±0.53a 9.65±0.63a 3.35±0.29a AG 45 0.30±0.21b 21.36±7.52b 18.17±6.35b 6.75±2.51b 6.07±2.65b 2.15±0.88b g5196 外显子14 CC 8 0.27±0.19a 20.18±6.65a 17.08±5.53a 6.29±2.08a 5.52±2.48a 2.01±0.84a CT 37 0.31±0.22a 21.62±7.68a 18.41±6.49a 6.84±2.58a 6.19±2.68a 2.17±0.88a TT 15 0.58±0.06b 32.48±1.00b 27.30±0.88b 10.31±0.53b 9.65±0.63b 3.35±0.29b g5253 外显子14 AA 8 0.27±0.19a 20.18±6.65a 17.08±5.53a 6.29±2.08 5.52±2.48a 2.01±0.84a AC 18 0.17±0.11b 16.88±3.51b 14.40±3.02b 5.35±1.37a 4.57±1.31b 1.67±0.51b CC 34 0.50±0.18a 28.92±6.70a 24.45±5.59a 9.16±2.26b 8.57±2.30a 2.96±0.77a g5322 外显子14 TT 16 0.56±0.12a 31.41±4.25a 26.41±3.56a 9.96±1.43a 9.29±1.53a 3.23±0.54a TC 44 0.30±0.22b 21.50±7.56b 18.29±6.38b 6.79±2.52b 6.12±2.66b 2.16±0.88b g5391 外显子14 AA 16 0.56±0.12a 31.41±4.25a 26.41±3.56a 9.96±1.43a 9.29±1.53a 3.23±0.54a AG 44 0.30±0.22b 21.50±7.56b 18.29±6.38b 6.79±2.52b 6.12±2.66b 2.16±0.88b g5417 外显子14 CC 15 0.58±0.06a 32.48±1.00a 27.30±0.88a 10.31±0.53a 9.65±0.63a 3.35±0.29a CG 45 0.30±0.21b 21.36±7.52b 18.17±6.35b 6.75±2.51b 6.07±2.65b 2.15±0.88b g8598 外显子25 AA 10 0.49±0.19a 29.11±6.31a 24.36±5.10a 9.27±2.13a 8.17±2.01a 2.87±0.69a AG 47 0.35±0.22b 23.24±8.09b 19.74±6.82b 7.32±2.63b 6.79±2.85b 2.38±0.96b GG 3 0.31±0.21 21.77±7.73 18.57±6.60 7.17±2.99 5.73±2.89 2.05±0.78 g9846 外显子29 GG 15 0.58±0.06a 32.48±1.00a 27.30±0.88a 10.31±0.53a 9.65±0.63a 3.35±0.29a AG 45 0.30±0.21b 21.36±7.52b 18.17±6.35b 6.75±2.51b 6.07±2.65b 2.15±0.88b g10221 外显子31 AA 15 0.58±0.06a 32.48±1.00a 27.30±0.88a 10.31±0.53a 9.65±0.63a 3.35±0.29a AG 45 0.30±0.21b 21.36±7.52b 18.17±6.35b 6.75±2.51b 6.07±2.65b 2.15±0.88b g10368 外显子31 TT 15 0.58±0.06a 32.48±1.00a 27.30±0.88a 10.31±0.53a 9.65±0.63a 3.35±0.29a TC 45 0.30±0.21b 21.36±7.52b 18.17±6.35b 6.75±2.51b 6.07±2.65b 2.15±0.88b g10444 外显子31 TC 44 0.30±0.22a 21.47±7.57a 18.26±6.40a 2.19±0.43a 6.11±2.67a 2.16±0.88a TT 15 0.58±0.06b 32.48±1.00b 27.30±0.88b 10.31±0.53b 9.65±0.63b 3.35±0.29b g10658 外显子32 CC 15 0.58±0.06a 32.48±1.00 a 27.30±0.88a 10.31±0.53a 9.65±0.63a 3.35±0.29a CA 45 0.30±0.21b 21.36±7.52b 18.17±6.35b 6.75±2.51b 6.07±2.65b 2.15±0.88b 注:均值±标准误。同一列中不同上标字母表示差异显著 (p<0.05)。 Note: Mean±SE. Values with the different superscript letters within same column are significantly different (p<0.05). 3. 讨论
3.1 肌球蛋白重链基因结构特征
肌球蛋白约占鱼类骨骼肌蛋白总质量的60%[31],在肌肉生长和运动过程中具有结构和动力学功能[32]。有研究表明在太阳科鱼类中,肌球蛋白重链基因在游泳和进食过程中对肌肉收缩起重要作用[33]。尽管肌球蛋白重链基因的分子结构已在多种鱼类中被报道,但在石斑鱼类中的研究较少。本研究获得杉虎杂交斑的肌球蛋白重链基因全长序列 (12 129 bp),包含36个外显子和35个内含子。经与其他物种比对分析,发现其在氨基酸组成及蛋白结构上均呈现较高的保守性。这也与其他文献报道的结果一致[12-13]。肌球蛋白重链基因包含4种结构域,即Myosin_N、MYSc、IQ和Myosin_tail_1。Myosin_N区域主要形成球状头部,保守性较强,是肌球蛋白家族的主要功能域。MYSc结构是分子马达,具有ATP酶活性。ATP酶合成和水解的速率与肌肉新陈代谢和生理学特征息息相关,例如收缩速率、肌肉类型等[34]。MYSc结构通过分子间构型变化可以调节肌球蛋白与ATP的结合能力,其与肌动蛋白结合位点可直接参与肌肉运动和能量供应[35]。IQ结构域为保守结构域,一般包含23个氨基酸,常作为EF-hand家族的结合位。在软体动物中,Ca2+ 和EF-hand结构域的结合可引发肌肉收缩[36]。Myosin_tail_1具有决定肌球蛋白与膜结合或与另外肌球蛋白分子相互反应的功能[37],同时也是构成粗肌丝的基础结构[38]。Myosin_N和IQ在本研究中显示出较高的序列保守性。而在MYSc和Myosin_tail_1区域,发现存在杉虎杂交斑特异性氨基酸位点,与其他物种均不同。肌球蛋白重链的蛋白高级结构在杉虎杂交斑与棕点石斑鱼中未见明显差异,表明该基因在杉虎杂交斑中仍具有正常的生物学功能。Nakaya和Watabe[39]提出,不同驯化温度下鲤 (Cyprinus carpio) Myosin_tail_1的α-螺旋结构有所差异,进而影响该基因的表达。相较于其他石斑鱼,杉虎杂交斑的肌球蛋白重链基因在氨基酸组成上发生改变,这种改变对基因的表达是否有影响仍有待进一步研究。除了仅存在于杉虎杂交斑中的特异性氨基酸位点外,还在Myosin_N、MYSc和Myosin_tail_1区域检测到石斑鱼与其他鱼类的差异性位点。例如在Myosin_N第37位氨基酸,石斑鱼为缬氨酸,而其他鱼类为苏氨酸、异亮氨酸。遗传进化分析结果也证明,所有石斑鱼类紧密地聚集成一个分支,与其他鱼类分开。以上结果表明,肌球蛋白重链基因在石斑鱼进化过程中具有高度保守性,因此可作为石斑鱼类与其他鱼类的分子鉴定标记。
3.2 肌球蛋白重链基因编码区SNPs位点分布特征
在杉虎杂交斑肌球蛋白重链基因编码区共检测到62个SNPs位点,其中有12个分布于外显子。这些外显子区域包括11、14、25、29、31以及32。其中位于外显子14的SNPs位点最多,有5个;其次是外显子31,为3个;其余外显子均只含有1个SNPs位点。外显子14编码第526—第628位氨基酸,参与MYSc蛋白结构域。外显子31编码第1 552—第1 654位氨基酸,参与Myosin_tail_1蛋白结构域。以上结果表明这2个区域的遗传变异相对较高。从基因进化的角度分析,功能基因在进化中其蛋白编码区保持高度的保守性以保障基因的正常功能。而通过对本研究中3种石斑鱼的肌球蛋白重链基因在4种蛋白结构域的氨基酸序列比对分析发现,该基因在石斑鱼中有着极高的保守性,但杉虎杂交斑仍存在与其他石斑鱼不同的特异性氨基酸位点。杉虎杂交斑是由棕点石斑鱼和清水石斑鱼种间杂交而来。杂交可以快速改变物种基因型,使子代具备更强的遗传可塑性和更好的生理适应性,为其具备优于亲本的表型性状奠定遗传基 础[40-41]。在2种杂交鲍 (Haliotis rubra × H. laevigata和H. discus hannai × H. fulgens) 中,均有文献报道杂交带来的遗传变异可能是它们呈现杂交优势的潜在原因[42-43]。杉虎杂交斑在生长速率上相较于亲本存在一定的优势[44],其肌球蛋白重链基因的核苷酸位点变异可能与其生长优势存在关联性,但具体作用机制本研究并未涉及,仍需进一步探究。
3.3 SNPs位点与生长性状关联分析
位于肌球蛋白重链基因外显子区域的12个SNPs位点,其Ho和PIC均高于0.4,属中等水平遗传多样性,但不符合高水平遗传多样性标准。本研究的实验样本杉虎杂交斑为棕点石斑鱼与清水石斑鱼杂交获得的子一代,属人工养殖群体。该群体虽未经历选育,但其亲本均为选育群体。而定向选育会导致群体遗传多样性水平呈现一定程度下降[45]。定向选育还可导致不利基因型逐渐丢失,而使有利基因型得到累积[46-47]。在12个SNPs位点中仅有4个位点包含3种基因型,其余位点均发生基因型丢失事件。而这种基因型丢失可能在杉虎杂交斑2个亲本的重要经济性状中属于不利基因型或与隐性致死基因位点连锁,进而在选育过程中遭到淘汰。在马氏珠母贝、鳙 (Aristichthys nobilis) 中也发现与生长性状显著相关的SNPs位点存在基因型丢失现象,认为这是适应选育的一种结果[12,46,48]。现代分子育种策略是通过育种技术使后代富集优势基因或基因型。但实际育种情况非常复杂,需要考虑基因与基因之间的互作关系,以及不同基因型对性状的作用效应大小。因此,某一基因型的丢失对整个性状的影响及其作用机制仍有待进一步探究。
通过重要调控基因筛选与经济性状相关的SNPs位点,获得分子标记,可应用于水产经济动物分子辅助选育工作[49-50]。在鳜MYH-7基因5' 端调控区筛选到与生长显著相关的SNP位点[4]。在大口黑鲈中,肌球蛋白重链基因在位于内含子的C-6811T位点存在与体质量、全长、体高以及尾柄长显著相关的基因型[13]。而本研究中将位于外显子区域的12个SNPs位点与生长性状进行关联分析发现,所有位点均与生长性状 (体质量、体长、全长、头长、体高、尾柄高) 呈显著性相关。在只有2种基因型的位点g4482、g5322、g5391、g5417、g9846、g10221、g10368、g10658,均为纯合基因型 (AA、TT、AA、CC、GG、AA、TT、CC) 的生长表型性状显著高于杂合基因型 (AG、TC、AG、CG、AG、AG、TC、CA),成为优势基因型。具有3种基因型的位点g5196、g5253、g8598和g10444,TT、CC、AA和TT分别为优势基因型,其生长表型数据均高于其他2种基因型。位点g5196 T>C和g8598 A>G显示出强连锁。2个位点的联合作用相较于单个位点其效应可能会更强,进而对性状的影响更大[51]。上述结果显示,杉虎杂交斑的肌球蛋白重链基因编码区存在与生长性状显著相关的SNPs。Chen等[12]在马氏珠母贝的肌球蛋白重链基因编码区同样检测到18个与生长性状显著相关的SNPs,与本研究结果一致。肌球蛋白重链基因氨基酸组成变异可对肌肉特性产生重要影响,进而影响其功能[52]。本研究仅发现位点g5417 C>G表现为错义突变,该位点位于MYSc区域,可能通过影响MYSc结构进而改变ATP酶活性。肌球蛋白重链头部ATP酶活性的差异与肌纤维的收缩速率极其相关[53],而高质量的肌肉收缩对肌肉的生长不可或缺[54]。其余与生长相关的SNPs位点均为同义突变。在豹纹鳃棘鲈 (Plectropomus leopardus) 中同样发现有生长相关SNPs位点属同义突变[55]。同义突变可影响基因mRNA的二级结构,从而改变在翻译、翻译整体速度以及蛋白折叠过程中暂停周期的时间长度[22,56-57]。这种普遍存在的长时间暂停会导致阅读框移码和蛋白质错误折叠[58]。已有研究表明,基因外显子区域的同义突变位点确实会通过影响转录和转录后过程进而影响该基因的表达和生物学功能[59]。本研究基于肌球蛋白重链基因SNPs位点与生长性状之间的关联分析,筛选获得生长候选标记。但由于样本数量有限,可能会带来抽样误差,从而影响研究结果的准确性,后续仍需继续扩大样本数量对候选位点进行进一步验证分析。
4. 结论
本研究成功获得了杉虎杂交斑肌球蛋白重链基因的核苷酸序列全长,识别出完整的蛋白编码区,并分析了其蛋白功能结构域及高级结构,为解析该基因的生物学功能奠定了基础。同时,通过与其他8种鱼类的蛋白结构域氨基酸序列进行比对分析及基于全部氨基酸序列构建遗传进化树,鉴定到能够区别石斑鱼类和其他鱼类的氨基酸位点以及杉虎杂交斑的特异性位点。揭示肌球蛋白重链基因在物种进化中有着极高的保守性,而杂交可为生物带来遗传变异。此外,基于杉虎杂交斑肌球蛋白重链基因上发现的62个SNPs位点,筛选到12个位于外显子且与生长表型性状显著相关的位点。本研究填补了石斑鱼肌球蛋白重链基因的研究空白,同时为杉虎杂交斑生长优势个体的筛选提供了候选分子标记。
-
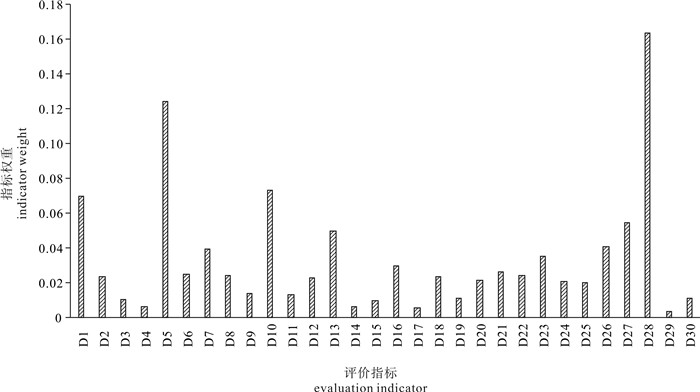
![]()
图 2 各项评价因子权重
D1. 水深;D2. 海流;D3. 透明度;D4. pH;D5. 溶解氧;D6. 活性磷酸盐;D7. 总无机氮;D8. 化学需氧量;D9. 悬浮物;D10. 石油类;D11. 硅酸盐;D12. 沉积物有机碳;D13. 沉积物硫化物;D14. 沉积物粒径;D15. 叶绿素a;D16. 初级生产力;D17. 浮游植物生物量;D18. 浮游植物多样性;D19. 浮游动物生物量;D20. 浮游动物多样性;D21. 鱼卵生物量;D22. 鱼卵多样性;D23. 仔鱼生物量;D24. 仔鱼多样性;D25. 大型底栖动物生物量;D26. 大型底栖动物多样性;D27. 游泳动物生物量;D28. 游泳动物多样性;D29. 潮间带大型底栖动物生物量;D30. 潮间带大型底栖动物多样性
Figure 2. Weight of each indicator
D1. depth; D2. current; D3. transparency; D4. pH; D5. DO; D6. IP; D7. IN; D8. COD; D9. SS; D10. Petroleum; D11. silicate; D12. sedimentary organic carbon; D13. sediment sulfide; D14. sediment grain size; D15. Chl-a; D16. primary productivity; D17. plant plankton biomass; D18. plant plankton biomass biodiversity; D19. zooplankton biomass; D20. zooplankton biomass biodiversity; D21. fish eggs biomass; D22. fish eggs biomass biodiversity; D23. larva fish biomass; D24. larva fish biomass biodiversity; D25. Macrobenthos biomass; D26. Macrobenthos biomass biodiversity; D27. swimming animals biomass; D28. swimming animals biomass biodiversity; D29. intertidal macrobenthos biomass; D30. intertidal macrobenthos biomass biodiversity
表 1 相应指标隶属函数及其确定依据
Table 1 Evaluating standard for quantitative index
指标
index隶属函数
fuzzy membership function确定依据
basis of evaluating standard水深/h
depth$\begin{cases}0 & x<10, x>60 \\ \left(\frac{x-10}{25-10}\right)^{0.53} & 10 \leqslant x \leqslant 25 \\ \left(\frac{60-x}{60-25}\right)^{1.37} & 25 \leqslant x \leqslant 60\end{cases}$ 《人工鱼礁建设技术规范》(SCT 9416—2014),20~30 m水深为投礁合适水深[17] 海流/m·s-1
current$\left\{\begin{array}{lr} 0 & >2 \\ 1 & 0.5 \leqslant x \leqslant 2 \\ \frac{x}{0.5} & 0<x<0.5 \end{array}\right.$ 稳定安全流速为0.5~2 m·s-1[18] 透明度/m
transparency$\left\{\begin{array}{rr} \frac{x}{4} & 0<x \leqslant 4 \\ 1 & x>4 \end{array}\right.$ 透明度基准值设定为4 m[15] 酸碱度
pH$\begin{cases}0 & x<7.0, x>8.5 \\ 1 & 7.0 \leqslant x \leqslant 8.5\end{cases}$ 《海水水质标准》 ρ(溶解氧)/mg·L-1
DO$\left\{\begin{array}{lr} 0 & x<5 \\ \left(\frac{x-5}{6-5}\right)^{0.011} & 5 \leqslant x \leqslant 6 \\ 1 & x>6 \end{array}\right.$ 《海水水质标准》 ρ(活性磷酸盐)/mg·L-1
IP$\left\{\begin{array}{lr} 1 & x<0.03 \\ \left(\frac{0.045-x}{0.045-0.03}\right) & 0.03 \leqslant x \leqslant 0.045 \\ 0 & x>0.45 \end{array}\right.$ 《海水水质标准》 ρ(总无机氮)/mg·L-1
IN$\left\{\begin{array}{lr} 1 & x<0.2 \\ \left(\frac{0.3-x}{0.3-0.2}\right) & 0.2 \leqslant x \leqslant 0.3 \\ 0 & x>0.3 \end{array}\right.$ 《海水水质标准》 ρ(化学需氧量)/mg·L-1
COD$\left\{\begin{array}{lr} 1 & x<2 \\ \left(\frac{3-x}{3-2}\right)^{0.011} & 2 \leqslant x \leqslant 3 \\ 0 & x>3 \end{array}\right.$ 《海水水质标准》 ρ(悬浮物)/mg·L-1
SS$\left\{\begin{array}{lr} 1 & x=0 \\ \left(\frac{10-x}{10-0}\right)^{0.073} & 0 \leqslant x \leqslant 10 \\ 0 & x>10 \end{array}\right.$ 《海水水质标准》 ρ(石油类)/mg·L-1
petroleum$\left\{\begin{array}{lr} 1 & x<0.05 \\ \left(\frac{0.5-x}{0.5-0.05}\right)^{0.57} & 0.5 \leqslant x \leqslant 0.05 \\ 0 & x>0.5 \end{array}\right.$ 《海水水质标准》 c(硅酸盐)/μmol·L-1
silicate$\left\{\begin{array}{lr} 0 & x<2 \\ \left(\frac{x-2}{4.7-2}\right)^{0.63} & 2 \leqslant x \leqslant 4.7 \\ 1 & x>4.7 \end{array}\right.$ 临界浓度为2 μmol·L-1,南海北部的活性硅酸盐年平均值为3.2 μmol·L-1[19] 沉积物有机碳
sedimentary organic carbon$\left\{\begin{array}{lr} 1 & x<2 \\ \left(\frac{4-x}{4-2}\right)^{0.74} & 2 \leqslant x \leqslant 4 \\ 0 & x>4 \end{array}\right.$ 《中华人民共和国海洋沉积物质量》(GB 18668—2002) 沉积物硫化物
sediment sulfide$\left\{\begin{array}{lr} 1 & x<300 \\ \left(\frac{600-x}{600-300}\right)^{0.46} & 600 \leqslant x \leqslant 300 \\ 0 & x>600 \end{array}\right.$ 《中华人民共和国海洋沉积物质量》(GB 18668—2002) 沉积物粒径/mm
sediment grain size$\begin{cases}1 & x>0.5 \\ \frac{x}{0.5} & x \leqslant 0.5\end{cases}$ 《粒度划分标准》 ρ(叶绿素a)/mg·m-3
Chl-a$\left\{\begin{array}{lr} 0 & x=0 \\ \left(\frac{x-0}{7.33-0}\right)^{0.61} & 0 \leqslant x<7.33 \\ 1 & x \geqslant 7.33 \end{array}\right.$ 该研究所获年度调查叶绿素a平均浓度3.17 mg·m-3作为评价值0.6 初级生产力/mg·(m2·d)-1
primary productivity$\left\{\begin{array}{lr} 0 & x<200 \\ \left(\frac{x-200}{600-200}\right)^{0.43} & 200 \leqslant x \leqslant 600 \\ 1 & x>600 \end{array}\right.$ 《饵料分级标准》 浮游植物生物量/104个·m-3
plant plankton biomass$\left\{\begin{array}{lr} 0 & x<20 \\ \left(\frac{x-20}{200-20}\right)^{0.28} & 20 \leqslant x \leqslant 200 \\ 1 & x>200 \end{array}\right.$ 《饵料分级标准》 浮游动物生物量/mg·m-3
zooplankton biomass$\left\{\begin{array}{lr} 0 & x<10 \\ \left(\frac{x-10}{100-20}\right)^{0.34} & 10 \leqslant x \leqslant 100 \\ 1 & x>100 \end{array}\right.$ 《饵料分级标准》 鱼卵生物量/10-3个·m-3
fish eggs biomass$\left\{\begin{array}{lr} 0 & x=0 \\ \left(\frac{x-0}{2\;600-0}\right)^{0.46} & 0 \leqslant x \leqslant 2\;600 \\ 1 & x>2\;600 \end{array}\right.$ 该研究所获年度调查鱼卵平均生物量855×10-3枚·m-3作为评价值0.6 仔鱼生物量/10-3个·m-3
larva fish biomass$\left\{\begin{array}{lr} 0 & x=0 \\ \left(\frac{x-0}{53-0}\right)^{1.38} & 0 \leqslant x \leqslant 53 \\ 1 & x>53 \end{array}\right.$ 该研究所获年度调查仔鱼平均生物量36.5×10-3尾·m-3作为评价值0.6 大型底栖动物生物量/g·m-3
macrobenthos biomass$\left\{\begin{array}{lr} 0 & x<5 \\ \left(\frac{x-5}{100-5}\right)^{0.175} & 5 \leqslant x \leqslant 100 \\ 1 & x>100 \end{array}\right.$ 《饵料分级标准》 游泳动物生物量/kg·h-1
swimming animals biomass$\left\{\begin{array}{lr} 0 & x=0 \\ \left(\frac{x-0}{100-0}\right)^{2.29} & 0 \leqslant x \leqslant 100 \\ 1 & x>100 \end{array}\right.$ 选择南海北部大陆架区年平均渔获量82 kg·h-1作为评价值0.6[20] 潮间带大型底栖动物生物量/g·m-3
intertidal macrobenthos biomass$\left\{\begin{array}{lr} 0 & x<0 \\ \left(\frac{x-0}{400-0}\right)^{4.3} & 0 \leqslant x \leqslant 400 \\ 1 & x>400 \end{array}\right.$ 该研究所获年度调查潮间带大型底栖动物生物量355 g·m-2作为评价值0.6 生物多样性阈值
biodiversity$\left\{\begin{array}{lr} 0 & x<0.6 \\ \left(\frac{x-0.6}{3.5-0.6}\right)^{0.7} & 0.6 \leqslant x \leqslant 3.5 \\ 1 & x>3.5 \end{array}\right.$ 《饵料分级标准》  下载: 导出CSV
下载: 导出CSV
表 2 4个季节监测数据的归一化值
Table 2 Normalization value of monitoring value at four seasons
评价因子
evaluation factor归一化值normalization value 春
spring夏
summer秋
autumn冬
winter平均
mean水深depth 0.295 0.285 0.379 0.262 0.305 海流current 0.580 0.580 0.580 0.580 0.580 透明度transparency 0.225 1.000 0.350 0.375 0.488 酸碱度pH 1.000 1.000 1.000 1.000 1.000 溶解氧DO 1.000 1.000 1.000 1.000 1.000 活性磷酸盐IP 1.000 1.000 1.000 1.000 1.000 总无机氮IN 0 0.354 0 0 0.089 化学需氧量COD 1.000 1.000 1.000 1.000 1.000 悬浮物SS 0 0 0.894 0 0.223 石油类petroleum 1.000 1.000 1.000 1.000 1.000 硅酸盐silicate 0.476 0.515 0.445 0.471 0.476 沉积物有机碳sedimentary organic carbon 1.000 1.000 1.000 1.000 1.000 沉积物硫化物sediment sulfide 1.000 1.000 1.000 1.000 1.000 沉积物粒径sediment grain size 1.000 1.000 1.000 1.000 1.000 叶绿素a Chl-a 0.502 1.000 0.523 0.177 0.551 初级生产力primary productivity 0 0.697 0 0 0.174 浮游植物生物量plant plankton biomass 0 1.000 0 0.220 0.305 浮游植物多样性plant plankton biomass biodiversity 0.764 0.498 0.805 0.464 0.633 浮游动物生物量zooplankton biomass 1.000 1.000 1.000 0.726 0.932 浮游动物多样性zooplankton biomass biodiversity 0.816 0.798 0.677 0.705 0.749 鱼卵生物量fish eggs biomass 0.403 1.000 0.268 0.358 0.507 鱼卵多样性fish eggs biomass biodiversity 0.766 0.752 0.776 0.713 0.752 仔鱼生物量larva fish biomass 1.000 1.000 0.335 0.192 0.632 仔鱼多样性larva fish biomass biodiversity 0.716 0.677 0.332 0.494 0.555 大型底栖动物生物量macrobenthos biomass 1.000 1.000 0.969 0.945 0.978 大型底栖动物多样性macrobenthos biomass biodiversity 0.488 0.481 0.427 0.536 0.483 游泳动物生物量swimming animals biomass 0.004 0.005 0.007 0 0.004 游泳动物多样性swimming animals biomass biodiversity 0.589 0.639 0.627 0.819 0.669 潮间带大型底栖动物生物量intertidal macrobenthos biomass 0.511 0.318 0.695 1.000 0.631 潮间带大型底栖动物多样性intertidal macrobenthos biomass biodiversity 0.406 0.454 0.507 0.507 0.469
下载: 导出CSV
-
[1] SCHROETER S C, REED D C, RAIMONDI P T. Effects of reef physical structure on development of benthic reef community: a large-scale artificial reef experiment[J]. Mar Ecol Prog Ser, 2015, 540: 43-55. doi: 10.3354/meps11483
[2] GRANNEMAN J E, STEELE M A. Effects of reef attributes on fish assemblage similarity between artificial and natural reefs[J]. ICES J Mar Sci, 2015, 72(8): 2385-2397. doi: 10.1093/icesjms/fsv094
[3] LEITAO F, SANTOS M N, ERZINI K, et al. Fish assemblages and rapid colonization after enlargement of an artificial reef off the Algarve coast (Southern Portugal)[J]. Mar Ecol, 2008, 29(4): 435-448. doi: 10.1111/j.1439-0485.2008.00253.x
[4] 李文涛, 张秀梅. 关于人工鱼礁礁址选择的探讨[J]. 现代渔业信息, 2003, 18(5): 3-6. doi: 10.3969/j.issn.1004-8340.2003.05.001 [5] MOUSAVI S H, DANEHKAR A, SHOKRI M R, et al. Identification of effective criteria for artificial reefs site selection using analytical hierarchy process (AHP) methodology (a case study: coral reefs in the Kish Island)[J]. Oceanography, 2011, 2(5): 7.
[6] KRAMER S H, HAMILTON C D, SPENCER G C, et al. Evaluating the potential for marine and hydrokinetic devices to act as artificial reefs or fish aggregating devices. based on analysis of surrogates in tropical, subtropical, and temperate US west coast and hawaiian coastal waters[R]. Honolulu: H.T. Harvey & Associates, 2015.
[7] SU D T, LIU T L, OU C H. Numerical investigation into effects of seabed topography on flows in and around artificial reefs[J]. Fish Sci, 2008, 74(2): 236-254. doi: 10.1111/j.1444-2906.2008.01518.x
[8] MOUSAVI S H, DANEHKAR A, SHOKRI M R, et al. Site selection for artificial reefs using a new combine Multi-Criteria Decision-Making (MCDM) tools for coral reefs in the Kish Island - Persian Gulf[J]. Ocean Coast Manag, 2015, 111: 92-102. doi: 10.1016/j.ocecoaman.2015.03.004
[9] 李梦杰. 组合式人工鱼礁的PIV二维流场效应与物理稳定性研究[D]. 上海: 上海海洋大学, 2015: 47.https://www.doc88.com/p-9774963162102.html [10] 林军, 吴辉, 章守宇. 非结构网格海洋模式在洞头人工渔礁区选址中的应用[J]. 浙江海洋学院学报(自然科学版), 2009, 28(1): 57-62. doi: 10.3969/j.issn.1008-830X.2009.01.011 [11] 许强, 章守宇. 基于层次分析法的舟山市海洋牧场选址评价[J]. 上海海洋大学学报, 2013, 22(1): 128-133. http://shhydxxb.ijournals.cn/shhy/article/abstract/20120300368 [12] 周艳波, 陈丕茂, 李辉权. 广东省柘林湾海域溜牛礁区建礁可行性研究[J]. 广东农业科学, 2011, 38(23): 10-14. doi: 10.3969/j.issn.1004-874X.2011.23.003 [13] XU F L, ZHAO Z Y, ZHAN W, et al. An ecosystem health index methodology (EHIM) for lake ecosystem health assessment[J]. Ecol Modell, 2005, 188(2/3/4): 327-339. https://www.sciencedirect.com/science/article/abs/pii/S0304380005001419
[14] 佟飞, 张秀梅, 吴忠鑫, 等. 荣成俚岛人工鱼礁区生态系统健康的评价[J]. 中国海洋大学学报(自然科学版), 2014, 44(4): 29-36. doi: 10.16441/j.cnki.hdxb.2014.04.005 [15] 李利. 廉州湾海域生态系统健康评价[D]. 青岛: 中国海洋大学, 2011: 39.https://www.doc88.com/p-9049332245159.html [16] 尹增强, 章守宇. 东海区资源保护型人工鱼礁生态效果评价体系的初步研究[J]. 海洋渔业, 2012, 34(1): 23-31. doi: 10.3969/j.issn.1004-2490.2012.01.004 [17] 贾后磊, 谢健, 彭昆仑. 人工鱼礁选址合理性分析[J]. 海洋开发与管理, 2009, 26(4): 72-75. https://www.doc88.com/p-6939870584788.html [18] 钟术求, 孙满昌, 章守宇, 等. 钢制四方台型人工鱼礁礁体设计及稳定性研究[J]. 海洋渔业, 2006, 28(3): 234-240. doi: 10.3969/j.issn.1004-2490.2006.03.011 [19] 袁梁英. 南海北部营养盐结构特征[D]. 厦门: 厦门大学, 2005: 23.https://www.zhangqiaokeyan.com/academic-degree-domestic_mphd_thesis/020311542750.html [20] 江艳娥, 林昭进, 黄梓荣. 南海北部大陆架区渔业生物多样性研究[J]. 南方水产, 2009, 5(5): 32-37. https://www.schinafish.cn/article/exportPdf?id=2a3e6d43-6ad6-4e54-839a-30edc4149a21 [21] 许强. 海洋牧场选址问题的研究[D]. 上海: 上海海洋大学, 2012: 78.https://www.zhangqiaokeyan.com/academic-degree-domestic_mphd_thesis/020314222076.html [22] JAXION-HARM J, SZEDLMAYER S T. Depth and artificial reef type effects on size and distribution of red snapper in the Northern Gulf of Mexico[J]. N Am J Fish Manag, 2015, 35(1): 86-96. doi: 10.1080/02755947.2014.982332
[23] 彭璇, 马胜伟, 陈海刚, 等. 夏季柘林湾-南澳岛海洋牧场营养盐的空间分布及其评价[J]. 南方水产科学, 2014, 10(6): 27-35. doi: 10.3969/j.issn.2095-0780.2014.06.004 [24] SCOTT M E, SMITH J A, LOWRY M B, et al. The influence of an offshore artificial reef on the abundance of fish in the surrounding pelagic environment[J]. Mar Freshw Res, 2015, 66(5): 429-437. doi: 10.1071/MF14064
[25] 朱小山, 杨炼锋. 广东柘林湾海水增养殖区环境质量评价[J]. 海洋通报, 2005, 24(1): 87-91. doi: 10.3969/j.issn.1001-6392.2005.01.015 [26] 朱四喜, 杨红丽, 朱臣, 等. 广东闸坡潮间带大型底栖动物的群落特征[J]. 浙江海洋学院学报(自然科学版), 2008, 27(1): 22-26. https://www.doc88.com/p-8592244344891.html [27] 佟文天, 李捷, 王海艳, 等. 海南三亚风景区潮间带软体动物的资源现状研究[J]. 海洋科学, 2013, 37(8): 11-15. http://qdhys.cnjournals.com/hykx/ch/reader/create_pdf.aspx?file_no=20130803&flag=1&journal_id=hykx&year_id=2013 [28] 龚志军, 谢平, 唐汇涓, 等. 水体富营养化对大型底栖动物群落结构及多样性的影响[J]. 水生生物学报, 2001, 25(3): 210-216. http://ssswxb.ihb.ac.cn/cn/article/id/5bbef668-6d5b-41ba-a2e8-61e254ee0fcd [29] CRESSON P, RUITTON S, OURGAUD M, et al. Contrasting perception of fish trophic level from stomach content and stable isotope analyses: a Mediterranean artificial reef experience[J]. J Exp Mar Bio Ecol, 2014, 452: 54-62. https://www.sciencedirect.com/science/article/abs/pii/S0022098113004061
[30] 王飞, 张硕, 丁天明. 舟山海域人工鱼礁选址基于AHP的权重因子评价[J]. 海洋学研究, 2008, 26(1): 65-71. http://qikan.cqvip.com/Qikan/Article/Detail?id=26904027 [31] 潘本锋, 宫正宇, 王帅, 等. 环境空气质量指数在应用中存在的问题及建议[J]. 中国环境监测, 2015, 31(1): 64-67. doi: 10.3969/j.issn.1002-6002.2015.01.013 [32] WALKER S J, SCHLACHER T A. Limited habitat and conservation value of a young artificial reef[J]. Biodivers Conserv, 2014, 23(2): 433-447. doi: 10.1007/s10531-013-0611-4
[33] AJEMIAN M J, WETZ J J, SHIPLEY-LOZANO B, et al. An analysis of artificial reef fish community structure along the Northwestern Gulf of Mexico Shelf: potential impacts of "Rigs-to-Reefs" programs[J]. PLoS ONE, 2015, 10(5): e0126354. https://pmc.ncbi.nlm.nih.gov/articles/PMC4425496/
[34] HENDERSON M J, FABRIZIO M C, LUCY J A. Movement patterns of summer flounder near an artificial reef: effects of fish size and environmental cues[J]. Fish Res, 2014, 153: 1-8. doi: 10.1016/J.FISHRES.2014.01.001






计量
- 文章访问数: 3467
- HTML全文浏览量: 228
- PDF下载量: 1114
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
