
Comparison analysis of major traits among four shell color selective lines of pearl oyster (Pinctada fucata)
-
摘要:
为评估合浦珠母贝(Pinctada fucata)白壳、黑壳、红壳和金壳色选育系F6代的育种性能,对养殖达18个月龄4种壳色合浦珠母贝的形态、质量、闭壳肌拉力和抗才女虫寄生病等12个性状指标进行了方差分析和多重比较。结果显示,4种壳色选育系间壳高、绞合线长、壳宽、体质量、壳质量、软体部质量、闭壳肌质量、闭壳肌拉力、右壳感染率和贝壳感染率均存在显著差异(P < 0.05)。综合相关分析表明,白壳色选育系可作为壳高和低才女虫感染率同时选育的最优育种群体,金壳色选育系可作为壳宽、体质量和闭壳肌拉力复合选育的最优育种群体。该研究结果为开展合浦珠母贝多性状复合选择育种工作提供了科学根据。
Abstract:To evaluate the breeding performance of sixth generations of pearl oysters (Pinctada fucata) (white, black, red and gold shell color selective lines), we analyzed 12 indices of morphological traits, weight, shell-closing strength and resistance to Polydora ciliata for an 18-month culture cycle by ANOVA. Significant difference was found in shell height, hinge length, shell width, body weight, shell weight, tissue weight, adductor weight, shell-closing strength, infestation ratio of right shell and infestation ratio of shell among four shell color selective lines (P < 0.05). The average values of selective breeding traits indicate that white shell color line was the best line for growth of shell height and resistance to P.ciliate. Golden shell color line was the best line for growth of shell width, body weight and shell-closing strength. The results provide references for multi-trait compound breeding of P. fucata.
-
合浦珠母贝(Pinctada fucata) 是中国培育海水珍珠的主要贝类之一,但随着珍珠养殖业的持续发展,养殖环境逐步恶化,近亲繁殖造成种质退化,母贝生长缓慢,个体小,死亡率高,育珠质量差等[1],严重制约了合浦珠母贝养殖产业的发展。因此,开展合浦珠母贝新品种选育工作刻不容缓。壳色作为贝类表型之一,已广泛应用于遗传育种和养殖生产[2-5]。在合浦珠母贝选育过程中发现在野生和养殖群体中均存在壳色的多态性,常见壳色有褐色、红色、黄色、黑色和白色等。WADA等[6]报道了以合浦珠母贝白壳色选育系作为外套膜小片供体时,可培育出高质量的白色珍珠。由此可见,壳色品系的选育为合浦珠母贝的养殖产业提供了新的发展方向。近年来,中国学者先后开展了合浦珠母贝白、黑、黄和红壳色选育系间多个世代的生长和成活对比[7-8]、遗传多样性分析[9-10]、壳棱柱层和珍珠质层金属元素含量[11]、插核育种效果[12]和壳色相关候选AFLP标记[13]等研究工作。这些研究为合浦珠母贝良种选育以及优质珍珠培育奠定了理论基础。
闭壳肌拉力可作为衡量贝类健康程度的一个生理性状指标,最先在日本被发现,并应用于合浦珠母贝遗传育种中。CHIHIRO等[14]研究发现日本珍珠贝闭壳肌拉力和死亡率成负相关。在中国,牛志凯等[15]研究发现合浦珠母贝群体间闭壳肌拉力存在显著差异(P < 0.05),闭壳肌拉力较优的YNSY和YNSZ群体可作为选育材料。多毛类凿贝才女虫(Polydora ciliata)是造成合浦珠母贝死亡率高的主要原因之一[16],用饱和盐水浸泡法和涂料涂覆法等方法防治效果不显著,而且在生产上操作也不便[17]。王爱民等[18]研究发现合浦珠母贝不同地理群体杂交后代多毛类寄生病的感染率低于自交群体后代,表明通过杂交选育可提高合浦珠母贝抗多毛类寄生虫病能力。陈明强等[19]利用合浦珠母贝家系才女虫感染情况数据,筛选出了抗才女虫寄生病能力较强的家系。而有关合浦珠母贝4种壳色选育系形态、闭壳肌拉力和抗才女虫寄生虫病等复合性状的研究尚未见报道。
该研究收集了合浦珠母贝白、黑、红和金4种壳色选育系F6代养殖18个月龄个体的形态、质量、闭壳肌拉力和抗才女虫寄生病等性状数据,利用SAS 9.3软件进行了4种壳色选育系间各性状的方差分析和多重比较,分析了各性状间的相关性,以期为合浦珠母贝多性状复合选择育种工作提供理论参考和基础数据。
1. 材料与方法
1.1 实验材料
2012年4月将课题组经多代壳色纯化选育的合浦珠母贝棱柱层为白、黑、红和金壳色4种壳色F5代个体作为亲本,在中国水产科学研究院南海水产研究所热带水产研究开发中心三亚安游基地采用解剖法收集精卵进行人工授精,依据同种壳色个体自繁的交配方案,培育了4种壳色F6代选育系幼虫。初期以湛江等鞭金藻(Isochrysis zhanjiangensis)为主要饵料,适当混合投喂亚心形扁藻(Platymonas subcordiformis)和活性酵母等,后期则以亚心形扁藻为主要饵料。每天换水1/3,连续充气培养。各壳色选育系贝苗投饵、换水、充气等管理一致。当贝苗长到壳长2 mm以上时,出苗移至陵水试验基地进行海上养殖,采用固定立桩式吊养。在养殖过程中及时进行换笼分笼、清理附着生物、检查死贝和敌害生物等,保持养殖和管理方法一致。
1.2 性状测量
贝苗养殖达到18个月龄后,对4种壳色合浦珠母贝个体的形态、质量、闭壳肌拉力和抗才女虫寄生病等性状数据进行测量。每个壳色选育系随机挑选4组,每组50只个体,共计200只个体。在清除贝体表面上的附着生物后,使用数显游标卡尺测量个体的壳长、壳高、壳宽和绞合线长,使用电子天平测量体质量、壳质量、软体部质量和闭壳肌质量,具体测量方法参照刘志刚等[20]。
依据AOKI等[21]和牛志凯等[15]的方法测量闭壳肌拉力,具体测量方法为把测量贝放在淡水中浸泡约10 min使其完全闭壳后,用毛巾擦干贝壳表面水分,从贝壳腹缘插入开口器,用数显拉力机测量贝开口幅度10 mm时的拉力值,精确到0.1 N。
依照陈明强等[19]进行抗才女虫寄生病的数据收集。统计每只贝才女虫寄生病的感染情况和每吊的感染率,以贝壳内表面有无虫管和珍珠瘤(Blisters)情况把个体分为无才女虫感染和有才女虫感染,其中贝壳内面珍珠层光滑,无虫管症状异常为无才女虫感染个体;而壳内面珍珠层上有小斑点,或闭壳肌等周围有脓包或溃烂性病灶[16]等情况的为有才女虫感染个体。性状指标分左壳感染率、右壳感染率和贝壳感染率,计算公式如下:
$$ \text { 左壳感染率: } P_{l i j}=\frac{\sum n_{l j}}{N} $$ (1) $$ \text { 右壳感染率: } P_{r i j}=\frac{\sum n_{r i j}}{N} $$ (2) $$ \text { 贝壳感染率: } P_{t i j}=\frac{\sum n_{l i j}+\sum n_{r i j}-\sum n_{i j}}{N} $$ (3) 式中Plij表示第i个壳色选育系第j组的左壳感染率,nlij表示第i个壳色选育系第j组的左壳感染个体,Prij表示第i个壳色选育系第j组的右壳感染率,nrij表示第i个壳色选育系第j组的右壳感染个体,Ptij表示第i个壳色选育系第j组的贝壳感染率,nij表示第i个壳色选育系第j组左右壳同时感染的个体,N为每组的总个体数(N=50)。
1.3 数据分析
使用统计软件Excel 2013进行4种壳色选育系形态、质量、闭壳肌拉力和抗才女虫寄生病等性状的数据整理,使用SAS软件ANOVA过程对4种壳色选育系各性状进行单因素方差分析(One-Way ANOVA)和Tukey多重比较分析,感染率在进行方差分析时进行反正弦转换[22]。使用SAS 9.3软件CORR过程进行各性状间的相关分析。
2. 结果与分析
2.1 壳色选育系间形态性状比较
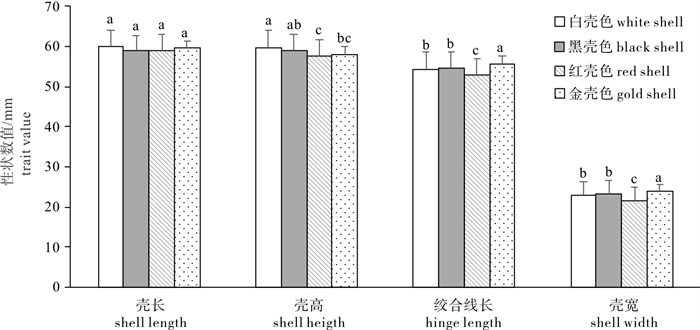
合浦珠母贝4种壳色选育系的4个形态性状比较分析见图 1。方差分析显示4种壳色选育系间壳高、绞合线长和壳宽存在显著差异(P < 0.05)。多重比较表明: 白壳色选育系与黑壳色选育系的壳高不存在显著差异(P>0.05),但与红壳色选育系间存在显著差异(P < 0.05);绞合线长和壳宽较优的金壳色选育系与其他3种壳色选育系间均存在显著差异(P < 0.05)。综合各壳色选育系间性状均值的大小和差异是否显著,分析得出以壳长和壳高为选育目标,白壳色选育系为最优育种群体;以壳宽为选育目标,金壳色选育系则为最优育种群体。
![]() 图 1 合浦珠母贝壳色选育系形态性状比较字母不同者为差异显著(P < 0.05),字母相同者为差异不显著(P>0.05),后图同此Figure 1. Comparison of four morphological traits among four shell color selective lines of P. fucataValues with different letters are significantly different (P < 0.05), while those with same letters are not significantly different (P>0.05). The same case in the following figure.
图 1 合浦珠母贝壳色选育系形态性状比较字母不同者为差异显著(P < 0.05),字母相同者为差异不显著(P>0.05),后图同此Figure 1. Comparison of four morphological traits among four shell color selective lines of P. fucataValues with different letters are significantly different (P < 0.05), while those with same letters are not significantly different (P>0.05). The same case in the following figure.2.2 壳色选育系间质量和闭壳肌拉力性状比较
合浦珠母贝4种壳色选育系质量和闭壳肌拉力性状比较分析见表 1。方差分析显示4种壳色选育系间体质量、壳质量、软体部质量、闭壳肌质量和闭壳肌拉力均存在显著差异(P < 0.05)。多重比较表明:体质量、壳质量和软体部质量较优的白壳色选育系、黑壳色选育系和金壳色选育系间不存在显著差异(P>0.05),但均与红壳色选育系存在显著差异(P < 0.05);闭壳肌质量较优的金壳色选育系和黑壳色选育系间不存在显著差异(P>0.05),均与白壳色选育系和红壳色选育系存在显著差异(P < 0.05);闭壳肌拉力较优的金壳色选育系和白壳色选育系间不存在显著差异(P>0.05),但均与黑壳色选育系和红壳色选育系存在显著差异(P < 0.05)。综合各壳色选育系间性状均值的大小和差异是否显著,分析可知以质量性状和闭壳肌拉力为选育目标,金壳色选育系为最优选育群体。
表 1 合浦珠母贝壳色选育系质量和闭壳肌拉力性状比较Table 1. Comparison of four weight and shell-closing strength traits among four shell color selective lines of P. fucata性状
trait白壳色
white shell黑壳色
black shell红壳色
red shell金壳色
gold shell体质量/g body weight 36.05±5.46a 35.69±6.80a 33.67±7.63b 37.27±5.61a 壳质量/g shell weight 18.99±3.27ab 19.09±3.73ab 18.63±4.28c 19.71±3.46a 软体部质量/g tissue weight 15.16±2.39a 15.18±3.04a 14.08±3.57b 15.19±2.58a 闭壳肌质量/g adductor weight 1.91±0.36b 2.22±0.57a 1.73±0.53c 2.28±0.54a 闭壳肌拉力/N shell-closing strength 75.76±16.95a 67.52±14.24b 67.43±13.34b 76.17±14.36a 注:同一列不同小写字母表示差异显著(P < 0.05)
Note:Values with different superscript letters within the same column are significantly different at 0.05 level.2.3 壳色选育系间抗才女虫寄生病性状比较
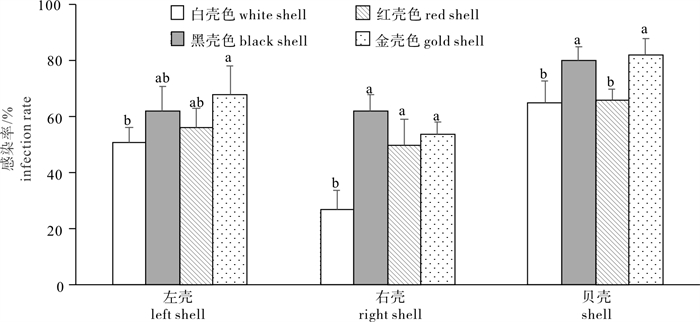
合浦珠母贝壳色选育系抗才女虫寄生病性状比较分析见图 2。方差分析显示4种壳色选育系间右壳感染率和贝壳感染率存在显著差异(P < 0.05)。多重比较表明, 右壳感染率较低的白壳色选育系与其他3种壳色选育系间存在显著差异(P < 0.05);贝壳感染率较低的白壳色选育系和红壳色选育系间不存在显著差异(P>0.05),均与黑壳色选育系和金壳色选育系间存在显著差异(P < 0.05)。综合各壳色选育系间性状均值的大小和差异是否显著,分析可知以低才女虫感染率为选育目标时,白壳色选育系为最优选育群体。
![]() 图 2 合浦珠母贝壳色选育系抗才女虫寄生病性状比较Figure 2. Comparison of three resistance to P. ciliata traits among four shell color selective lines of P. fucata
图 2 合浦珠母贝壳色选育系抗才女虫寄生病性状比较Figure 2. Comparison of three resistance to P. ciliata traits among four shell color selective lines of P. fucata3. 讨论
该研究表明,4种合浦珠母贝壳色选育系间壳高、绞合线长、壳宽、体质量、壳质量、软体部质量和闭壳肌质量均存在显著差异(P < 0.05),结合各壳色选育系形态和质量性状指标均值的大小和差异是否显著,可初步认为白壳色选育系是进行壳长或壳高选育的最优育种群体,金壳色选育系是进行壳宽或质量性状选育的最优育种群体。该研究结果与朱晓闻等[10]报道的4种合浦珠母贝壳色选育系中白壳色选育系生长速度最快的结果基本一致,但与王庆恒等[7]、DENG等[8]和陈静等[9]研究结果不同,他们的研究发现黑壳色选育系生长性状数值大于其他3种壳色选育系。上述研究结果的差异可能是由于选育基础群体亲本来源、选育技术路线和养殖环境等不同造成。
闭壳肌拉力可作为合浦珠母贝遗传育种工作的选育指标,它与合浦珠母贝的生长、成活、生理、营养成分和珍珠层分泌能力等性状具有相关性[23-24],牛志凯等[15]和OKAMOT等[25]分别研究发现合浦珠母贝群体间和家系间闭壳肌拉力存在显著差异(P < 0.05),闭壳肌拉力较优的群体(家系)可作为选育材料。ISHIKAWA等[26]运用回归法估计了18月龄日本合浦珠母贝闭壳肌拉力的现实遗传力,其数值为0.29。该研究表明4种壳色选育系间闭壳肌拉力也存在显著差异(P < 0.05),综合各壳色选育系间闭壳肌拉力均值的大小和差异是否显著,表明金壳色选育系的闭壳肌拉力最优,可以进一步作为闭壳肌拉力性状遗传改良的育种群体。
该研究进行了4种合浦珠母贝壳色选育系左壳、右壳和贝壳才女虫感染率的比较,结合各壳色选育系3个感染率指标均值的大小和差异是否显著,得出白壳色才女虫感染率最小,表明白壳色选育系最具抗病选育潜力,可以作为抗才女虫寄生病品系的育种材料。王爱民等[18]对合浦珠母贝不同地理群体自交和杂交后代感染多毛类寄生病进行比较分析,发现杂交群体后代多毛类寄生病的感染率低于自交群体的后代,表明通过杂交选育可提高合浦珠母贝抗多毛类寄生虫能力。陈明强等[19]利用合浦珠母贝家系才女虫感染情况数据,筛选出了抗才女虫寄生病能力较强的家系,这些研究推动了通过遗传改良技术培育抗才女虫寄生病珍珠贝新品种的育种工作进展。
该研究综合4种壳色选育系各性状均值的大小和差异是否显著的分析结果,表明白壳色选育系可作为壳高和低才女虫感染率同时选育的最优育种群体,金壳色选育系可作为壳宽、体质量和闭壳肌拉力复合选育的最优育种群体。该研究全面评估了合浦珠母贝4种壳色选育系的育种性能,筛选出了多性状育种材料,为开展合浦珠母贝多性状复合选择育种工作提供了科学依据。
-
![]()
图 1 合浦珠母贝壳色选育系形态性状比较
字母不同者为差异显著(P < 0.05),字母相同者为差异不显著(P>0.05),后图同此
Figure 1. Comparison of four morphological traits among four shell color selective lines of P. fucata
Values with different letters are significantly different (P < 0.05), while those with same letters are not significantly different (P>0.05). The same case in the following figure.
![]()
图 2 合浦珠母贝壳色选育系抗才女虫寄生病性状比较
Figure 2. Comparison of three resistance to P. ciliata traits among four shell color selective lines of P. fucata
表 1 合浦珠母贝壳色选育系质量和闭壳肌拉力性状比较
Table 1 Comparison of four weight and shell-closing strength traits among four shell color selective lines of P. fucata
性状
trait白壳色
white shell黑壳色
black shell红壳色
red shell金壳色
gold shell体质量/g body weight 36.05±5.46a 35.69±6.80a 33.67±7.63b 37.27±5.61a 壳质量/g shell weight 18.99±3.27ab 19.09±3.73ab 18.63±4.28c 19.71±3.46a 软体部质量/g tissue weight 15.16±2.39a 15.18±3.04a 14.08±3.57b 15.19±2.58a 闭壳肌质量/g adductor weight 1.91±0.36b 2.22±0.57a 1.73±0.53c 2.28±0.54a 闭壳肌拉力/N shell-closing strength 75.76±16.95a 67.52±14.24b 67.43±13.34b 76.17±14.36a 注:同一列不同小写字母表示差异显著(P < 0.05)
Note:Values with different superscript letters within the same column are significantly different at 0.05 level. 下载: 导出CSV
下载: 导出CSV
-
[1] 黎火金, 刘宝锁, 罗会, 等. 合浦珠母贝幼贝生长性状的遗传参数估计[J]. 中国水产科学, 2013, 20(6): 1182-1187. doi: 10.3724/SP.J.1118.2013.01182 [2] 丛日浩, 李琪, 葛建龙, 等. 长牡蛎4种壳色家系子代的表型性状比较[J]. 中国水产科学, 2014, 21(3): 494-502. [3] 葛建龙, 李琪, 于红, 等. 长牡蛎3种壳色家系间杂交子代生长和存活比较[J]. 水产学报, 2015, 39(3): 345-352. doi: 10.11964/jfc.20140809436 [4] EVANS S, CAMARA M D, LANGDON C J. Heritability of shell pigmentation in the Pacific oyster, Crassostrea gigas[J]. Aquaculture, 2009, 286(3/4): 211-216. https://www.sciencedirect.com/science/article/pii/S0044848608006881
[5] 王照旗, 韩学凯, 白志毅, 等. 三角帆蚌紫色选育系1龄阶段内壳色及生长性状的遗传参数估计[J]. 水产学报, 2014, 38(5): 644-650. doi: 10.3724/SP.J.1231.2014.49071 [6] WADA K T. Perspective of genetic improvement in the culture of pearl oysters[J]. Fish Sci, 2002, 68: 726-729. doi: 10.2331/fishsci.68.sup1_726
[7] 王庆恒, 邓岳文, 杜晓东, 等. 马氏珠母贝4个壳色选系F1幼虫的生长比较[J]. 中国水产科学, 2008, 15(3): 488-492. doi: 10.3321/j.issn:1005-8737.2008.03.016 [8] DENG Y W, FU S, LIANG F L, et al. Fertilization, hatching, survival, and growth of third-generation colored pearl oyster (Pinctada martensii) stocks[J]. J Appl Aquac, 2013, 25(2): 113-120. doi: 10.1080/10454438.2013.788311
[9] 陈静, 刘志刚, 孙小真, 等. 马氏珠母贝4种壳色选育系F3的生长特性和遗传多样性比较[J]. 上海海洋大学学报, 2010, 19(5): 588-595. https://www.cnki.com.cn/Article/CJFDTotal-SSDB201005003.htm [10] 朱晓闻, 刘志刚, 王辉, 等. 马氏珠母贝(Pinctada martensii)四种壳色选育系F5的生长及遗传多样性分析[J]. 海洋与湖沼, 2012, 43(1): 160-165. doi: 10.11693/hyhz201201026026 [11] 邹柯姝, 张殿昌, 郭华阳, 等. 4种壳色合浦珠母贝贝壳棱柱层和珍珠质层7种金属元素质量分数的比较分析[J]. 南方水产科学, 2015, 11(3): 74-79. doi: 10.3969/j.issn.2095-0780.2015.03.012 [12] 符韶, 谢绍河, 邓岳文, 等. 利用马氏珠母贝4个壳色系F3培育厚层优质珍珠[J]. 水产学报, 2012, 36(9): 1418-1424. doi: 10.3724/SP.J.1231.2012.27905 [13] ZOU K, ZHANG D, GUO H, et al. A preliminary study for identification of candidate AFLP markers under artificial selection for shell color in pearl oyster Pinctada fucata[J]. Gene, 2014, 542(1): 8-15. doi: 10.1016/j.gene.2014.03.029
[14] CHIHIRO O, AKIRA K, MASAHIRO H, et al. Shell-closing strength of pearl oyster, Pinctada fucata martensii and its relation-ship to mortality and various parts weight of oyster[J]. Aquac Sci, 2006, 54 (3) : 293 -299. https://www.jstage.jst.go.jp/article/aquaculturesci1953/54/3/54_3_293/_article
[15] 牛志凯, 刘宝锁, 张东玲, 等. 合浦珠母贝3个地理群体杂交后代生长性状和闭壳肌拉力的比较分析[J]. 南方水产科学, 2015, 11(1): 26-32. doi: 10.3969/j.issn.2095-0780.2015.01.004 [16] 石耀华, 王爱民, 吴星. 中国养殖马氏珠母贝多毛类寄生虫病的调查[J]. 海洋科学, 2004, 28(7): 13-18. doi: 10.3969/j.issn.1000-3096.2004.07.004 [17] 王爱民, 石耀华, 吴星. 4种防治马氏珠母贝多毛类寄生病方法的效果比较[J]. 海洋水产研究, 2004, 25(2): 41-46. [18] 王爱民, 阎冰, 叶力, 等. 马氏珠母贝不同地理种群内自繁和种群间杂交子一代感染多毛类寄生病的分析[J]. 海洋水产研究, 2003, 24(3): 38-44. doi: 10.3969/j.issn.1000-7075.2003.03.007 [19] 陈明强, 刘宝锁, 谭才钢, 等. 合浦珠母贝家系间生长性状和感染多毛类寄生病的比较分析[J]. 广东农业科学, 2015, 42(5): 87-91. doi: 10.3969/j.issn.1004-874X.2015.05.018 [20] 刘志刚, 王辉, 孙小真, 等. 马氏珠母贝经济性状对体重决定效应分析[J]. 广东海洋大学学报, 2007, 27(4): 15-20. doi: 10.3969/j.issn.1673-9159.2007.04.004 [21] AOKI H, ISHIKAWA T, FUJIWARA T, et al. Utility of shell-closing strength as the indicator of good health in breeding and culture management of Japanese pearl oyster Pinctada fucata[J]. Aquaculture, 2010, 308(S1): S115-S118. https://www.sciencedirect.com/science/article/pii/S0044848610004692
[22] 明道绪. 生物统计附试验设计[M]. 北京: 中国农业出版社, 2008: 127-129. [23] OKAMOTO C, KOMARU A, HAYASH M, et al. Shell-closing strength of pearl oyster, Pincata fucata martensii and its relationship to mortality and various parts weight of oyster[J]. Aquac Sci, 2006, 54(3): 293-299. https://www.jstage.jst.go.jp/article/aquaculturesci1953/54/3/54_3_293/_article
[24] AOKI H, KOMARU A. Studies on the breeding and culture management of pearl oysters using shell-closing strength as an indicator[J]. Fish Genet Breed Sci, 2010, 40: 85-96.
[25] OKAMOTO C, KOMARU A, HAYASH M, et al. Variation of shell-closing strength among several families in pearl oyster, Pinctada fucata martensii[J]. Aquac Sci, 2006, 54(4): 525-529. doi: 10.1002/2211-5463.12502
[26] ISHIKAWA T, OKAMOTO C, HAYASH M, et al. The heritability of the shell-closing strength in the Japanese pearl oyster, Pinctada fucata martensii[J]. Aquac Sci, 2009, 57(1): 77-82. https://www.jstage.jst.go.jp/article/aquaculturesci/57/1/57_77/_article
-
期刊类型引用(8)
1. 邹杰,彭慧婧,郑德斌,张守都. 马氏珠母贝杂交家系生长性状与遗传参数分析. 南方农业学报. 2021(12): 3221-3227 .  百度学术
百度学术
2. 魏海军,邓正华,陈明强,邓林琪,谭春明,李有宁,王雨,王爱民. 不同壳色合浦珠母贝生产性能评估及其雌雄群体间的差异. 南方农业学报. 2020(02): 412-420 . 百度学术
3. 陈明强,魏海军,李有宁,邓正华,王雨,马振华,于刚,孙静. 企鹅珍珠贝F_3选育系的珍珠层颜色及生长性状的比较分析. 南方农业学报. 2020(03): 659-668 . 百度学术
4. 魏海军,陈明强,邓正华,李有宁,王雨,马振华,于刚. 4个企鹅珍珠贝F_3选育系的生长存活差异及其生长规律. 中国水产科学. 2020(06): 624-636 . 百度学术
5. 孙静,陈明强,魏海军,邓正华,李有宁,金朝焕,王继金,王雨,石承开,黄超旬. 不同地理群体合浦珠母贝双列杂交子代数量性状比较及其相关和通径分析. 南方农业学报. 2020(06): 1451-1461 . 百度学术
6. 陈健,白丽蓉,罗会,喻达辉. 2种贝龄合浦珠母贝数量性状的相关与通径分析. 南方农业学报. 2020(10): 2557-2564 . 百度学术
7. 邓正华,陈明强,李有宁,赵旺,郑兴,刘芊,王继金,王雨. 3种野生珍珠贝表型性状比较及通径分析. 广东农业科学. 2019(01): 122-132 . 百度学术
8. 王祎哲,邓正华,王雨,李有宁,于刚,黄桂菊,陈明强. 合浦珠母贝不同壳色选育系F6数量性状的相关性和通径分析. 中国水产科学. 2018(05): 988-997 . 百度学术
其他类型引用(5)
计量
- 文章访问数: 3345
- HTML全文浏览量: 201
- PDF下载量: 1182
- 被引次数: 13
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
