
Early development of squamation for Oplegnathus fasciatus
-
摘要:
采用茜素红染色法对条石鲷(Oplegnathus fasciatus)早期发育阶段鳞片的形成和发育过程进行了观察和测量。结果显示条石鲷鳞片的起始中心有3个,分别是鳃盖后缘、侧线上方和尾柄处。在水温为24~27 ℃的人工培育条件下,20日龄在这3个起始中心最早出现鳞片,鳃盖后缘至胸鳍基部的鳞被向上下前后扩展,胸鳍后缘的侧线上方鳞被向下和向前后扩展,尾柄处的鳞被前后扩展,头顶部的鳞被向四周扩展,到40日龄时鱼体全身被鳞进入幼鱼期。通过观察还发现,鳞片向下方发育的速度快于上方,上下方向的发育速度快于前后方向。向前扩展到鳃盖后暂停,向后发育到达尾柄末端,最后出现鳞片的部位是鳃盖。条石鲷鳞片为栉鳞,较细小。在鱼体的同一时期同一部位,鳞片的大小存在较大的差异。初生鳞鳞嵴(环片)数目与发育阶段的关系可用二次方程表示。
Abstract:This paper studies the scale formation at early developmental stages of Oplegnathus fasciatus by using Alizarin red staining method. The fish have three scale starting centers, including the posterior margin of gill cover, above side of lateral line and the caudal peduncle. At rearing water temperatures of 24~27 ℃, primary scales appeared first in these three starting centers at 20th day. Scales originated from back edge of operculum to pectoral fin base toward upward and downward as well as before and after expansion; scales originated above lateral line on back edge of pectoral fin expanded downward and forward and backward, and those originated from caudal peduncle in front and rear extension. The scales on the top of head extended to periphery. At 40th day, the whole body surface was covered by scales, coming into young fish stage. The development of scales extending to lower area of the body surface was faster than that in the upper area surface, and extension in upper and lower direction was faster than that to direction before and after. Expansion paused after extending to the operculum, backward to the end of caudal peduncle, and the last part of scales coverage was operculum. Scales of O. fasciatus belonged to ctenoid scale type with smaller size. The sizes of scales were very different in the same area at the same developmental stage. Ridge number on primary scale in relation to developmental stages of O. fasciatus could be expressed by quadratic equation.
-
Keywords:
- Oplegnathus fasciatus /
- squamation /
- primary scale /
- development
-
鳞片是由鱼类皮肤原始干细胞定向分化产生[1],是鱼体与外界接触的边缘组织。其基本功能除减少游泳产生的摩擦力、维持体型骨架之外,还起到保护鱼体免遭病原损害的作用。同时,鳞片的形态结构也是研究鱼类分类学特征、资源分布、栖息环境以及生长规律的重要依据[2]。根据鳞片的外形、构造和发生特点,可将鳞片划分为3种基本类型:盾鳞、硬鳞和骨鳞。骨鳞是最常见的一种鳞片,为真骨鱼类所有,呈覆瓦状排列。根据鳞片露出部的表面边缘的构造不同可将骨鳞分为两类:圆鳞和栉鳞。
条石鲷(Oplegnathus fasciatus)隶属于辐鳍鱼纲、鲈形目、鲈亚目、石鲷科、石鲷属,自然分布范围在东海、黄海、台湾海峡和太平洋[3]。俗称日本鹦鹉鱼,栖息于温热带近海碎石、岩礁或珊瑚礁底质的底层水域。条石鲷形态优美,色泽艳丽,肉质细嫩,营养价值高,是一种极具养殖前景和观赏价值的海产名贵鱼类[4]。近年来对条石鲷的早期发育已有许多研究[5-9],但关于鳞片的形成在中国未见有报道,同时由于南北方水域环境的差异,鳞片的发育在时间和速度上也可能会存在一些差异。该文采用茜素红染色法初步分析了条石鲷鳞片最初出现的时间和位置,以及鳞片形成的时间及各个发育阶段的特点,旨在丰富条石鲷早期发育的生物学文库,同时通过对鳞片发育过程的观察,判断鱼类种苗的培育效果,及时调整种苗生产管理措施。
1. 材料与方法
1.1 材料
观察样本为采自广东省饶平县中国水产科学研究院南海水产研究所海水鱼类试验基地人工孵化和培育的0~40日龄的条石鲷仔稚鱼。
1.2 方法
将人工催产的受精卵置于容积为0.5 m3的玻璃纤维孵化桶内进行孵化,连续充气,仔鱼孵出后在面积为1/15~2/15 hm2,水深为1.5~2 m的池塘内进行人工育苗,育苗期间水温为24~27 ℃(其中前期培育水温为22~23 ℃,后期为24~27 ℃),盐度29~30,pH 7.4~8.2。
实验方法采用茜素红染色法[10]。从2008年5月2日到6月11日育苗期间每天用手抄网随机采集10~20尾样本,用5%甲醛溶液保存,将要观察鳞片的鱼苗样本放入清水中浸泡3 h,然后放入2%茜素红溶液中染色,染色后的样本用滤纸吸干体表水分,在体视显微镜下观察鳞片出现的部位及其分布情况、鳞片上的鳞嵴数、鳞焦的中心以及水平鳞列数。用解剖针小心将鱼体的鳞片掀起,用解剖刀轻轻刮下,在显微镜下观察、拍照。
1.3 实验数据处理
采用SPSS 18.0软件,对鳞嵴数与发育阶段的相关关系分别用直线方程、幂函数方程以及二次方程进行拟合,对相关系数做显著性检验,依最小剩余平方和来确定最适回归方程。
2. 结果
2.1 早期鳞被的形成
2.1.1 条石鲷鳞片起始中心及鳞被形成
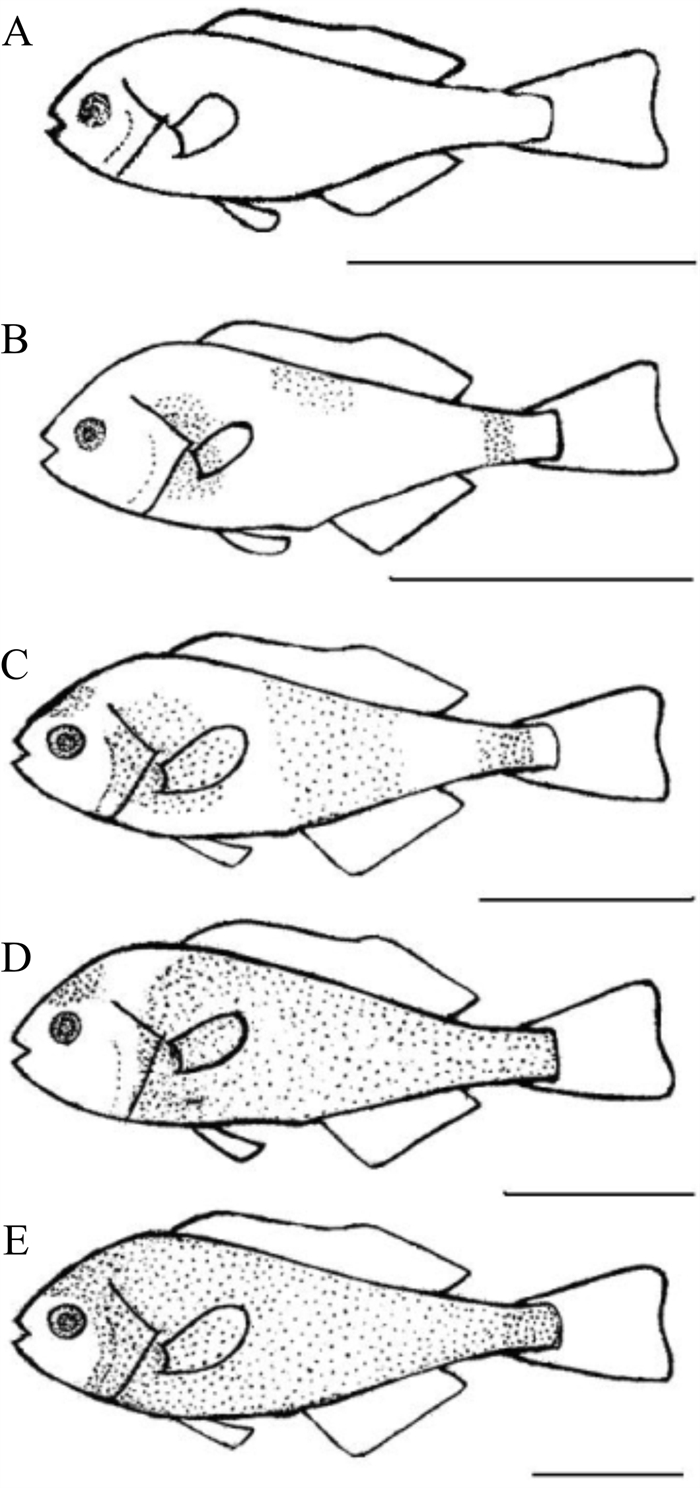
条石鲷鳞片最初出现的部位在鳃盖后缘至胸鳍基部、胸鳍后缘的侧线上方和尾柄处,共有3个鳞被起始中心。鳃盖后缘至胸鳍基部的鳞被向上下前后扩展,胸鳍后缘的侧线上方鳞被向下和向前后扩展,尾柄处的鳞被前后扩展,头顶部的鳞被向四周扩展,最后连成一片,鳞被覆盖全身。根据条石鲷鳞被在鱼体体表的形成过程将其划分为5个阶段(图 1)。
阶段A:鳞片未出现,全身无鳞。
阶段B:鳃盖后缘至胸鳍基部、胸鳍后缘的侧线上方和尾柄处开始出现鳞片,水平鳞列数1~2。
阶段C:鳞被向各方扩展,在侧线的上下方均出现鳞被,尾柄处的鳞被发育较快,水平鳞列数5~11。头部开始出现鳞片。
阶段D:鳞被向全身扩展,鳃盖后缘至胸鳍基部、胸鳍后缘的侧线上方和尾柄处的鳞被连成一片,并覆盖大部分鱼体。鱼体腹部出现鳞被,鳞被发育较均匀,尾柄全部覆盖鳞被,鳃盖上也出现鳞片,鳞被发育较晚。
阶段E:体表鳞被与头部鳞被连合,鳞被发育完全,鱼体全身具鳞。
2.1.2 鳞被形成特点
条石鲷鳞被向下方扩展的速度快于上方,上下扩展速度快于前后扩展,向前扩展到鳃盖后暂停,向后发育到达尾柄末端,最后出现鳞片的部位是鳃盖。同一个部位鳞被的发育速度并不完全相同。
2.1.3 条石鲷鳞被形成的体长和日龄
条石鲷仔鱼最初出现鳞片的最小个体为全长19 mm、体长15 mm,最大个体为全长26 mm、体长20 mm,为孵化后18~20日龄(表 1);鳞被发育完全时的最小个体为全长36 mm、体长29.9 mm,最大个体为全长66 mm、体长51 mm,为孵化后35~40日龄。最初出现鳞片和鳞被发育完全时个体的大小差异最大为全长47 mm、体长36 mm,22日龄。
表 1 条石鲷鳞被发育阶段与个体生长的关系(n=220)Table 1 Relationship between early developmental stage of squamation and body length and total length of O. fasciatus鳞被发育阶段
developmental stage全长/mm total length 体长/mm body length 日龄
day after hatching最大maximum 最小minimum 最大maximum 最小minimum 阶段A Stage A 22 15 18 11 15~20 阶段B Stage B 26 19 20 15 18~20 阶段C Stage C 39 26 31 20 25~32 阶段D Stage D 35 32 27 25 32~35 阶段E Stage E 66 36 51 29.9 35~40 2.2 鳞片形态结构和初生鳞的发育
2.2.1 鳞片的形态结构
条石鲷鳞片为细栉鳞,长方形,前区较平截,后区边缘和鳞嵴具细栉齿(图 2);具放射状辐射沟2~6条,向基区(前区)辐射,起始处不一致,有的始于鳞焦,有的始于中途,成鱼辐射沟界限不太清晰;鳞焦偏于后区。
![]() 图 2 条石鲷成鱼鳞片的形态特征(标尺=0.5 mm)Fig. 2 Morphological characteristics of scale in adult O. fasciatus(scale=0.5 mm)
图 2 条石鲷成鱼鳞片的形态特征(标尺=0.5 mm)Fig. 2 Morphological characteristics of scale in adult O. fasciatus(scale=0.5 mm)2.2.2 初生鳞的发育
条石鲷初生鳞为圆形或椭圆形,表面光滑、透明,鳞嵴(环片)、栉齿、鳞焦和辐射沟等均无。鳞片在发育过程中,其外形、鳞嵴、栉齿、鳞焦和辐射沟均随个体大小和日龄而发生变化。条石鲷初生鳞的发育特征与鳞被形成阶段的关系为:
阶段A:鳞片未出现。
阶段B:鳃盖后缘初生鳞的形态为较原始的圆鳞,鳞嵴为1~3,胸鳍基部鳞嵴为4~6;侧线上方鳞片的鳞嵴为3~6,鳞片极薄易碎;尾部鳞片鳞嵴为3~6(图 3-B-a,b,c)。
![]() 图 3 条石鲷各个鳞片形成阶段的鳞片B-a. 侧线上鳞片;B-b. 尾柄鳞片;B-c. 鳃盖后缘鳞片;C-a. 侧线上鳞片;C-b. 胸鳍基鳞片;C-c. 头部鳞片;C-d. 鳃盖鳞片;C-e. 尾柄鳞片;C-f. 腹部鳞片;D-a. 侧线上鳞片;D-b. 侧线下鳞片;D-c. 胸鳍基鳞片;D-d. 头部鳞片;D-e. 鳃盖鳞片;D-f. 尾柄鳞片;E-a. 侧线上鳞片;E-b. 鳃盖鳞片;E-c. 尾柄鳞片;E-d. 胸鳍基鳞片;标尺=0.25 mmFig. 3 Scales at different developmental stages in O. fasciatusB-a. scale up lateral line; B-b. scale of caudal peduncle; B-c. scale of posterior margin of gill cover; C-a. scale up lateral line; C-b. scale of pectoral fin base; C-c. scale of head; C-d. scale of gill cover; C-e. scale of caudal peduncle; C-f. scale of ventral; D-a. scale up lateral line; D-b. scale down ateral line; D-c. scale of pectoral fin base; D-d. scale of head; D-e. scale of gill cover; D-f. scale of caudal peduncle; E-a. scale up the lateral line; E-b. scale of gill cover; E-c. scale of caudal peduncle; E-d. scale of pectoral fin base; scale=0.25 mm
图 3 条石鲷各个鳞片形成阶段的鳞片B-a. 侧线上鳞片;B-b. 尾柄鳞片;B-c. 鳃盖后缘鳞片;C-a. 侧线上鳞片;C-b. 胸鳍基鳞片;C-c. 头部鳞片;C-d. 鳃盖鳞片;C-e. 尾柄鳞片;C-f. 腹部鳞片;D-a. 侧线上鳞片;D-b. 侧线下鳞片;D-c. 胸鳍基鳞片;D-d. 头部鳞片;D-e. 鳃盖鳞片;D-f. 尾柄鳞片;E-a. 侧线上鳞片;E-b. 鳃盖鳞片;E-c. 尾柄鳞片;E-d. 胸鳍基鳞片;标尺=0.25 mmFig. 3 Scales at different developmental stages in O. fasciatusB-a. scale up lateral line; B-b. scale of caudal peduncle; B-c. scale of posterior margin of gill cover; C-a. scale up lateral line; C-b. scale of pectoral fin base; C-c. scale of head; C-d. scale of gill cover; C-e. scale of caudal peduncle; C-f. scale of ventral; D-a. scale up lateral line; D-b. scale down ateral line; D-c. scale of pectoral fin base; D-d. scale of head; D-e. scale of gill cover; D-f. scale of caudal peduncle; E-a. scale up the lateral line; E-b. scale of gill cover; E-c. scale of caudal peduncle; E-d. scale of pectoral fin base; scale=0.25 mm阶段C:圆鳞逐渐发育成栉鳞,胸鳍基鳞嵴为4~6,侧线上方鳞片的鳞嵴为2~6,侧线下方鳞片的鳞嵴为3~8,尾柄处鳞片的鳞嵴为4~7,头部鳞片鳞嵴为2~5(图 3-C-a,b,c,d,e,f)。
阶段D:侧线上方鳞片鳞嵴为2~9,侧线下方鳞片的鳞嵴为12~16,腹部鳞片鳞嵴为5~9,胸鳍基鳞片鳞嵴为3~11,尾柄部鳞嵴为4~8,头部的鳞片鳞嵴为3~6,鳃盖上的鳞片鳞嵴为3~8(图 3-D-a,b,c,d,e,f)。
阶段E:侧线上方鳞片鳞嵴为8~13,下方为11~16,尾柄处鳞嵴为13~15,胸鳍基鳞片鳞嵴为8~12,鳃盖后缘鳞片鳞嵴为15~18(图 3-E-a,b,c,d)。
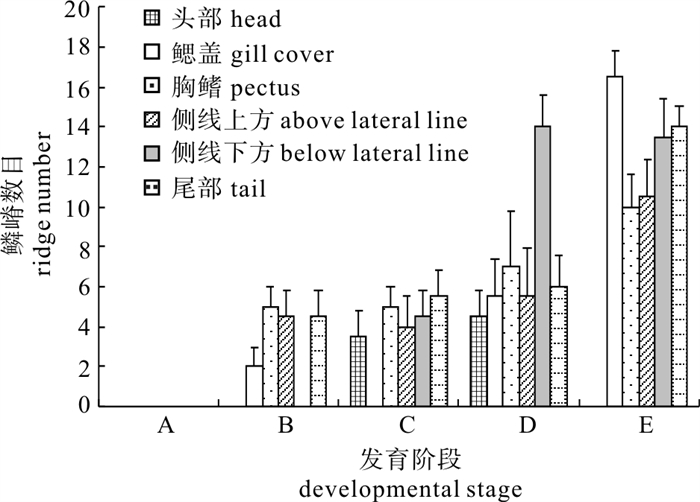
条石鲷初生鳞鳞嵴(环片)数目与发育阶段的关系为条石鲷初生鳞在发育过程中,鱼体不同部位鳞嵴(环片)数目随个体大小和日龄而发生变化(图 4)。从总体趋势看,随着发育进程,鱼体不同部位鳞片的鳞嵴数逐渐增加。经统计学拟合,鳞嵴数与发育阶段的相关关系可分别用直线方程y=2.88x-2.96(n=220,R2=0.918 1)、幂函数方程y=6.802 4ln(x)-0.833 3(n=220,R2=0.827 4)以及二次方程y=0.414 3x2 +0.394 3x-0.06(n=220,R2=0.944 6)表示。式中y为鳞嵴数,x为发育阶段。统计结果显示3种方程的适合度均较高,表现出较好的拟合性,因而三者都可以较好地描述条石鲷初生鳞鳞嵴(环片)数目与发育阶段的关系,依最小剩余平方和来选择回归方程,在3种相关方程中,以二次方程的描述更为吻合。
![]() 图 4 条石鲷鱼体不同部位初生鳞鳞嵴数目与发育阶段的关系(n=220)Fig. 4 Ridge number on primary scale in relation to developmental stages of O. fasciatus
图 4 条石鲷鱼体不同部位初生鳞鳞嵴数目与发育阶段的关系(n=220)Fig. 4 Ridge number on primary scale in relation to developmental stages of O. fasciatus3. 讨论
3.1 鱼类鳞片最先出现部位和起始位置数量
此研究观察显示,条石鲷具有3个鳞片起始中心。徐恭昭等[11]把鱼类的鳞片起始中心分成两大类型:1)鳞片发育只具有1个起始中心,2)具有2个或2个以上起始中心。鲷科的黑鲷(Sparus macroeephlus)[12]、真鲷(Pagrus major)[13],科的黑边石斑鱼(Epinephelus fasciatus)[14],鳗鲡科的奥克兰鳗鲡(Anguilla australis)和大鳗鲡(A.dieffenbachii)[15]等,只具有1个起始中心。具有2个或2个以上起始中心的种类,如鲻科的梭鱼(Mugil soiuy)[11]、溪鳉科的花斑溪鳉(Rivulus marmoratus)[16]、鲱科的大鳞油鲱(Brevooritia patronus)[17]有2个;齿鳉科杂色鳉(Cyprinodon variegatus variegatus)[18]、太阳鱼科的暗斑棘盖太阳鱼(Pomoxis nigromaeulatus)[19]有3个;鳢科的乌鳢(Channa agrus)[20]、底鳉科的沼泽底鳉(Fundulus confluentus)、底鳉(F.heteroclitus)等种类[18]有4个。而鲤科的齐口裂腹鱼(Schizothorax prenanti)[21]和重口裂腹鱼(S.davidi)[22]稚鱼鳞片发生起始位置有6个,以此为中心向四周扩展延伸直到覆盖全鱼体。据SIRE和ARNULF[23]报道,4种硬骨鱼类的鳞片发生起始位置有7个。从现有这些文献来看,不同鱼类物种之间鳞片发生起始点的数目及其覆盖方式存在着差异,是否与物种进化以及栖息环境具有一定的相关性尚待深入探讨。
3.2 鱼类鳞被覆盖过程
此研究显示,条石鲷稚鱼体表开始长出形态较为原始的圆鳞,而后圆鳞逐渐发育成栉鳞,其发生过程与圆鳞鱼类大致类似[24]。在鳞片开始形成的时间以及鳞片发育完全的时间,条石鲷和黑鲷等种类相似,而且鳞片发育的过程也很相近。关于鱼类鳞片最初出现的时间和位置的问题,KUBO和YASHIWARA[25]把鱼体最早出现鳞片的区域分为两大类型:1)在躯体的前部,如鲤(Cyprinus carpio)和鳟(Salmo trutta)等。草鱼(Ctenopharyngodon idella)、鲢(Hypophthalmichthys molitrix)、鳙(Aristrichthys nobilis)以及重口裂腹鱼等种类,其鳞片最早均出现在躯干前部的侧线处,鲤和草鱼的鳞片以1个起始点为中心,等速纵向和横向伸展,最后覆盖腹部中线;鳙和鲢鳞片向腹部和尾柄部扩展的速度快于向背部方向,当腹部与尾柄部基本覆盖时鳞片才从侧线上方向背部和由尾柄上部向前伸展,头后背部最后覆盖[26]。2)在鱼体后部尾柄末端的侧线上,如白刺盖太阳鱼(P. annuaris)及淡水鼓鱼(Aptodiotus grunnieus)等种类。根据PENAZ[27]、ABLE和LAMOMACA[28]的报道,许多淡水鱼类和鲆鲽鱼类的鳞片发育起始于尾柄。软骨鱼类澳大利亚虎鲨(Heterodontus portusjacksoni)和眶嵴虎鲨(H. galeatus) 的鳞片发育起始于尾鳍[29];斯氏罗非鱼(Tilapia sparrmani)的鳞片最先出现在尾柄中部,沿侧线处向前覆盖的速度快于向后,并以等速向腹部和背部方向扩展,最后覆盖头后背部[25]。从鳞片形成的整个过程来看,条石鲷早期鳞片的形成在整个鱼群中呈不同步性,鳞片出现的最初时间,鳞片发育的速度,鳞片发育完全的时间,在鱼体大小以及日龄方面均有很大变化。同样是20日龄的仔鱼有的没有一片鳞,有的已开始形成鳞片。所以笔者认为,决定鱼类鳞片发育的因素是多方面的,它并不仅受到水温或是饵料的影响,还可能受到其他一些因素的影响。这有待于以后进一步的研究。
-
![]()
图 2 条石鲷成鱼鳞片的形态特征(标尺=0.5 mm)
Figure 2. Morphological characteristics of scale in adult O. fasciatus(scale=0.5 mm)
![]()
图 3 条石鲷各个鳞片形成阶段的鳞片
B-a. 侧线上鳞片;B-b. 尾柄鳞片;B-c. 鳃盖后缘鳞片;C-a. 侧线上鳞片;C-b. 胸鳍基鳞片;C-c. 头部鳞片;C-d. 鳃盖鳞片;C-e. 尾柄鳞片;C-f. 腹部鳞片;D-a. 侧线上鳞片;D-b. 侧线下鳞片;D-c. 胸鳍基鳞片;D-d. 头部鳞片;D-e. 鳃盖鳞片;D-f. 尾柄鳞片;E-a. 侧线上鳞片;E-b. 鳃盖鳞片;E-c. 尾柄鳞片;E-d. 胸鳍基鳞片;标尺=0.25 mm
Figure 3. Scales at different developmental stages in O. fasciatus
B-a. scale up lateral line; B-b. scale of caudal peduncle; B-c. scale of posterior margin of gill cover; C-a. scale up lateral line; C-b. scale of pectoral fin base; C-c. scale of head; C-d. scale of gill cover; C-e. scale of caudal peduncle; C-f. scale of ventral; D-a. scale up lateral line; D-b. scale down ateral line; D-c. scale of pectoral fin base; D-d. scale of head; D-e. scale of gill cover; D-f. scale of caudal peduncle; E-a. scale up the lateral line; E-b. scale of gill cover; E-c. scale of caudal peduncle; E-d. scale of pectoral fin base; scale=0.25 mm
![]()
图 4 条石鲷鱼体不同部位初生鳞鳞嵴数目与发育阶段的关系(n=220)
Figure 4. Ridge number on primary scale in relation to developmental stages of O. fasciatus
表 1 条石鲷鳞被发育阶段与个体生长的关系(n=220)
Table 1 Relationship between early developmental stage of squamation and body length and total length of O. fasciatus
鳞被发育阶段
developmental stage全长/mm total length 体长/mm body length 日龄
day after hatching最大maximum 最小minimum 最大maximum 最小minimum 阶段A Stage A 22 15 18 11 15~20 阶段B Stage B 26 19 20 15 18~20 阶段C Stage C 39 26 31 20 25~32 阶段D Stage D 35 32 27 25 32~35 阶段E Stage E 66 36 51 29.9 35~40  下载: 导出CSV
下载: 导出CSV
-
[1] SHARPE P T. Fish scale development: hair today, teeth and scales yesterday?[J] Curr Biol, 2001, 11(18): R751-752. doi: 10.1016/S0960-9822(01)00438-9
[2] SIRE J Y, AKIMEENKO M A. Scale development in fish: a review, with description of sonic hedgehog (shh) expression in the zebrafish (Danio rerio)[J]. Int J Dev Biol, 2004, 48(2/3): 233-247. https://pubmed.ncbi.nlm.nih.gov/15272389/
[3] 刘敏, 陈驍, 杨圣云. 中国福建南部海洋鱼类图鉴(第一卷)[M]. 北京: 海洋出版社, 2013: 224. [4] 蔡文超, 区又君, 李加儿. 南海区养殖条石鲷的胚胎发育[J]. 南方水产, 2009, 5(4): 31-35. doi: 10.3969/j.issn.1673-2227.2009.04.006 [5] 区又君, 李加儿, 林锋. 南海区驯养条石鲷亲鱼的初次性成熟和产卵[J]. 南方水产科学, 2014, 10(5): 82-86. doi: 10.3969/j.issn.2095-0780.2014.05.012 [6] 区又君, 李加儿, 艾丽, 等. 广东池塘培育条石鲷仔、稚、幼鱼的早期发育和生长[J]. 南方水产科学, 2014, 10(6): 66-71. doi: 10.3969/j.issn.2095-0780.2014.06.009 [7] 艾丽, 区又君, 李加儿, 等. 条石鲷早期发育阶段鳍的分化[J]. 福建农林大学学报(自然科学版), 2010, 39(5): 517-522. [8] 区又君, 李加儿, 艾丽. 条石鲷早期发育阶段体表色素的变化和色素带的形成[J]. 水产学报, 2014, 38(10): 1722-1730. https://www.cnki.com.cn/Article/CJFDTotal-SCKX201410007.htm [9] 区又君, 李加儿, 艾丽. 条石鲷早期发育阶段消化系统组织学研究[J]. 华南农业大学学报, 2015, 36(1): 23-27. [10] 孟国林. 骨科实验技术[M]. 北京: 人民卫生出版社, 2012: 49. [11] 徐恭昭, 郑澄伟, 杨纯武, 等. 梭鱼早期鳞被的形成[G]//鱼类学论文集: 第四辑. 北京: 科学出版社, 1985: 127-133. [12] 丁彦文, 李加儿, 区又君. 黑鲷早期鳞被的形成[J]. 热带海洋, 1991, 10(3): 16-20. https://www.cnki.com.cn/Article/CJFDTotal-RDHY199103002.htm [13] 福原修. マタィ稚仔鱼形態学的研究Ⅰ. 初生鳞の発生与生長[J]. 南西海区水産研究所研究报告, 1976 (9): 13-18. [14] 川辺勝俊. アカハタ仔稚魚の鰭と鱗の発達[J]. 水産増殖, 2000, 48(1): 39-46. [15] JELLYMAN D J. Scale development and age determination in New Zealand freshwater eels (Anguilla spp.)[J]. NZ J Mar Freshw Res, 1979, 13(1): 23-30. doi: 10.1080/00288330.1979.9515777
[16] PARK E, LEE S. Scale growth and squamation chronology for the laboratory-reared hermaphroditic fish Rivulus marmoratus (Cyprinodontidae)[J]. Jpn J Ichthyol, 1988, 34(4): 476-482. doi: 10.1007/BF02905653
[17] CHAPOTON R B. Scale development in the gulf menhaden, Brevoortia patronus[J]. Trans Am Fish Soc, 1967, 96(1): 60-62.
[18] ABLE K W, SAKOWICZ G P, LAMOMACA J C. Scale formation in selected fundulid and cyprinodontid fishes[J]. Ichthyol Res, 2009, 56(1): 1-9. https://www.researchgate.net/publication/226580143_Scale_formation_in_selected_fundulid_and_cyprinodontid_fishes
[19] COOPR J A. Scale development as related to growth of juvenile black crappie, Pomoxis nigromaculatus Lesueur[J]. Trans Am Fish Soc, 1971, 100(3): 570-572. doi: 10.1577/1548-8659%281971%29100%3C570%3ASDARTG%3E2.0.CO%3B2
[20] 谢从新, 周洁, 熊传喜, 等. 乌鳢早期鳞被的形成及鳞片形态特征[J]. 华中农业大学学报, 1996, 15(1): 68-72. https://www.cqvip.com/QK/90088X/19961/2191510.html [21] 严太明, 唐仁军, 刘小帅, 等. 齐口裂腹鱼鳞片发生及覆盖过程研究[J]. 水生生物学报, 2014, 38(2): 298-303. http://qikan.cqvip.com/Qikan/Article/Detail?id=49301965 [22] 严太明, 刘小帅, 杨淞, 等. 重口裂腹鱼鳞片发育和覆盖过程[J]. 动物学杂志, 2014, 49(3): 391-398. https://www.cqvip.com/QK/94741X/201403/49841067.html [23] SIRE J Y, ARNULF I. The development of squamation in four teleostean fishes with a survey of the literature[J]. Jpn J Ichthyol, 1990, 37(2): 133-143. https://www.researchgate.net/publication/225584844_The_development_of_squamation_in_four_teleostean_fishes_with_a_survey_of_the_literature
[24] ZHU J, ZHANG X M, GAO T X. Morphological studies on the development of melanophores and scales in malpigmented Paralichthys olivaceus[J]. Acta Hydrobiologica Sinica, 2004, 28(6): 653-658.
[25] KUBO I, YOSHIWARA T. Early development of the squamation in Tilapia sparrmani[J]. Jpn J Ichthyol, 1971, 18(2): 90-93.
[26] 李思发. 五种鱼的鳞被复盖过程及其同生长的关系[J]. 水产学报, 1983, 7(4): 343-351. https://www.cnki.com.cn/Article/CJFDTotal-SCKX198304005.htm [27] PENAZ M. A general framework of fish ontogeny: a review of the ongoing debate[J]. Folia Zool, 2001, 50(4): 241-256. https://www.researchgate.net/publication/296638001_A_general_framework_of_fish_ontogeny_A_review_of_the_ongoing_debate
[28] ABLE K W, LAMOMACA J C. Scale formation in selected western North Atlantic flatfishes[J]. J Fish Biol, 2006, 68(6): 1679-1692. https://www.researchgate.net/publication/229441256_Scale_formation_in_selected_western_North_Atlantic_flatfishes
[29] JOHNSON Z, SMITH M M, JOSS J M P, et al. Early scale development in Heterodontus (Heterodontiformes; Chondrichthyes): a novel chondrichthyan scale pattern[J]. Acta Zoologica, 2007, 88(30): 249-256. https://www.academia.edu/925530/Early_scale_development_in_Heterodontus_Heterodontiformes_Chondrichthyes_a_novel_chondrichthyan_scale_pattern
-
期刊类型引用(3)
1. 唐杰,周硕,王亚军,胡佳宝,王想兵,王冠林,蒋欢,严小军. 银鲳早期鳞被的覆盖和初生鳞的发育. 水生生物学报. 2023(12): 1948-1953 .  百度学术
百度学术
2. 周胜杰,胡静,杨蕊,杨其彬,马振华. 三斑海马早期发育观察. 南方农业学报. 2021(08): 2302-2310 . 百度学术
3. 王志强,刘寒,耿瑞静,王卫民. 团头鲂和斑马鱼鳞片的发生发育过程比较研究. 淡水渔业. 2019(01): 20-26 . 百度学术
其他类型引用(1)




计量
- 文章访问数: 3189
- HTML全文浏览量: 143
- PDF下载量: 2101
- 被引次数: 4
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
