
Effect of nutrient enrichment on phytoplankton growth in the adjacent waters of the Xisha Yongxing Islands in late fall
-
摘要:
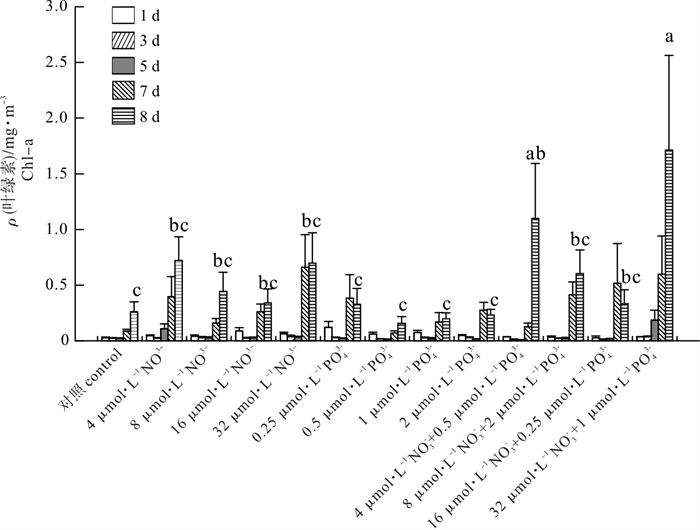
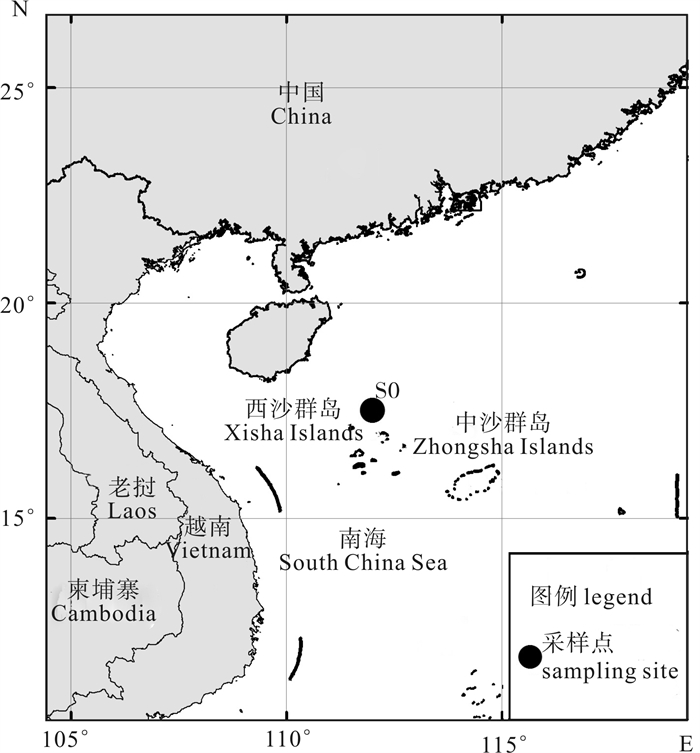
为探讨西沙海域氮(N)和磷(P)对浮游植物生长限制的影响,2014年秋末在永兴岛附近海域S0站(111°59.843′E,17°30.364′N)进行了N、P营养盐的现场添加实验。添加氮的浓度依次为4 μmol · L-1、8 μmol · L-1、16 μmol · L-1、32 μmol · L-1,添加磷的浓度依次为0.25 μmol · L-1、0.5 μmol · L-1、1 μmol · L-1、2 μmol · L-1,添加氮和磷的浓度组合依次为4 μmol · L-1+0.5 μmol · L-1 (N : P=8 : 1)、8 μmol · L-1 +2 μmol · L-1 (N : P=4 : 1)、16 μmol · L-1+0.25 μmol · L-1 (N : P=64 : 1)、32 μmol · L-1+1 μmol · L-1 (N : P=32 : 1)。结果显示:加富N+P后,叶绿素a浓度显著增长(P < 0.05),其中,32 μmol · L-1NO3--N+1 μmol · L-1PO43--P添加组和4 μmol ·L-1NO3--N+0.5 μmol · L-1 PO43--P添加组叶绿素质量浓度从初始的0.06 mg · m-3分别达到1.71 mg · m-3和1.10 mg ·m-3,说明添加N/P范围在8~32之间可以促进浮游植物生长。在单独加富N和P后,叶绿素a浓度均无显著的增加(P>0.05),说明单独添加N和P不能促进浮游物生长。限制因子分析表明,S0站点的浮游植物生长为N+P共同限制。
Abstract:To investigate the effect of nitrate and phosphate on phytoplankton growth, we carried out field experiments and enrichment incubation experiments in the adjacent waters of the Xisha Yongxing Islands (S0 station: 17°30.364′N, 111°59.843′E) in late fall of 2014. The added nitrogen concentrations were 4 μmol · L-1, 8 μmol · L-1, 16 μmol · L-1 and 32 μmol · L-1; the added phosphorus concentrations were 0.25 μmol · L-1, 0.5 μmol · L-1, 1 μmol · L-1 and 2 μmol · L-1; the added mixed nitrogen and phosphorus concentrations were 4 μmol · L-1+0.5 μmol · L-1, 8 μmol · L-1+2 μmol · L-1, 16 μmol · L-1+0.25 μmol · L-1 and 32 μmol · L-1+1 μmol · L-1. The results show that the chlorophyll a increased significantly after adding mixed nitrate and phosphate (P < 0.05). The chlorophyll a concentration increased from 0.06 mg · m-3 to 1.71 mg · m-3 and 1.10 mg · m-3 at S0 station after adding 32 μmol · L-1NO3--N+1 μmol · L-1PO43--P and 4 μmol · L-1NO3--N+0.5 μmol · L-1 PO43--P mixed nitrate and phosphate, respectively. It is suggested that enrichment of mixed nitrate and phosphate (N/P: 8~32) could promote the growth of phytoplankton. However, enrichment of single nitrate and single phosphate did not promote the phytoplankton growth because the chlorophyll a concentration showed no significant increase by adding single nitrate and single phosphate at S0 station. Consequently, the restriction factors of phytoplankton growth at S0 station were both nitrate and phosphate concentrations.
-
鲍隶属于软体动物门,腹足纲,前鳃亚纲,原始腹足目,鲍科鲍属[1],具有很高的营养和药用价值,自古以来就有“一口鲍一口金”的说法,在中国的饮食文化中占有很重要的地位[2-3]。中国是世界第一养鲍大国[4-5],主要养殖种类是分布在大连、山东等北方沿海的皱纹盘鲍(Haliotis.dicus hannai Ino)和广东、福建等南方沿海的杂色鲍(H.diversicolor Reeve)[6],近年来由于鲍养殖产业的高速发展而出现近交严重、种质退化以及环境恶化致使鲍病害频发,皱纹盘鲍[7-8]和杂色鲍[9-11]相继出现了重大病害,严重影响了鲍养殖业的发展[12]。针对种质退化问题,科研工作者正在致力于培育出具有更高抗性的新品种,如“大连1号”杂交鲍[13]和“东优1号”杂色鲍[14]新品种都具有强于亲本的优良性状。
科研工作者通过选育、杂交、多倍体诱导、雌核发育等技术试图培育出性状更好的新品种[15-21]。关于耐高温和抗病新品种的培育,国内外学者已经取得了一些进展[22-24],其中对大菱鲆(Scophthalmus maximus)、刺参(Apostichopus japonicus Selenka)、褐牙鲆(Paralichthys olivaceus)和近江牡蛎(Crassostrea ariakensis Gould)等耐高温品系的选育研究较多[25-30],此外,在水生植物中也有对坛紫菜(Porphyra haitanensis)[31]耐高温品系的培育,然而在鲍中还未见到耐高温选育的系统研究报道。此研究所选试验动物是青岛选育的2个不同自然群体皱纹盘鲍的杂交鲍,这也是首次将该种杂交鲍引进南方进行试养,为进一步研究高温胁迫对鲍体内热休克蛋白基因的表达情况和鲍的耐高温机制奠定基础,以期为培育适合南方海区养殖的抗逆性强的新品种提供参考。
1. 材料与方法
1.1 试验材料
试验时间为2013年11月中旬至2013年12月末,选取生长健康、壳长接近10 mm的皱纹盘鲍450只,平均放置于15个尺寸为50 cm×30 cm×40 cm的玻璃水缸中,玻璃缸内放置鲍砖以供鲍附着并有一定的遮光作用。试验共分5组,每组3个平行,每个平行30个重复。试验开始时选取壳长相近的幼鲍进行试验,经测量并进行数据分析后各组鲍的平均壳长约11 mm,平均壳宽约8 mm,湿质量约0.2 g,均值、标准差和组间的差异性详见表 1。
表 1 各组鲍试验开始时的规格Table 1 Size of each group at the beginning of research温度/℃ temperature 壳长/mm shell length 壳宽/mm shell width 湿质量/g wet weight 自然温度 natural temperature 11.08±0.25a 7.91±0.13a 0.23±0.02a 25 11.25±0.06a 8.12±0.09a 0.24±0.02a 28 10.83±0.24a 7.79±0.08a 0.21±0.01a 31 10.95±0.11a 7.90±0.10a 0.22±0.01a 32 11.29±0.56a 8.26±0.43a 0.23±0.01a 注:同列相同上标字母表示差异不显著,反之差异显著,下表同此
Note:The same superscripts within the column indicate no significant difference,while different ones indicate significant difference. The same case in the following figure.1.2 试验方法
将上述15个玻璃水缸平均分为5组,每组3个平行,其中1组为对照组,温度维持自然水温(试验开始时温度为22 ℃,在20~23 ℃间变化),其他4组利用加热棒(JEBO佳宝)采用温度渐变(30 ℃以下每3 d升温1 ℃,30 ℃以上每周升温1 ℃)的方式分别升温至25 ℃、28 ℃、31 ℃和32 ℃。试验开始前和结束后对其形态指标进行测定,试验期间记录死亡情况。
1.3 日常管理
每个水缸接通一个充气石保证溶解氧不低于5 mg · L-1,试验期间每天下午投喂适量鲍配合饲料,次日早晨采用虹吸法从底部清除残饵和粪便并添加经过滤并预热至所需温度的新鲜海水,依据试验设计记录死亡情况和控制加热棒的温度来调节试验组温度,温差范围控制在±0.5 ℃以内。
1.4 形态特征测量
用电子数显游标卡尺测量每只鲍试验前后的壳长和壳宽,电子天平测量每缸中所有鲍的总湿质量,计算平均湿质量以减少水分的影响。按照LGR=(L2-L1) · t-1,WGR=(W2-W1) · t-1的公式计算壳长、壳宽和湿质量的日增长率,其中L1和W1为初始所有个体数据,L2和W2为终末剩余个体数据,其中控温组死亡个体数据以初始数据即增长率为0计算在内求得平均值,t为试验持续时间。
1.5 数据处理
应用SPSS 19.0软件进行单因素方差分析,分析不同温度下5组鲍的壳长、壳宽和湿质量的日增长率。
2. 结果
2.1 温度对皱纹盘鲍幼鲍生长速率的影响
经过45 d的控温试验后,各组鲍的平均壳长、壳宽和湿质量均有所增长,并有随着温度的升高增长幅度减小的趋势,自然温度组的鲍壳长增加了6 mm,壳宽增长接近5 mm,湿质量在试验结束时增长接近约3.0 g,增长显著,其他各组的壳长和壳宽增长相近且均小于3 mm,湿质量随着温度的升高增长幅度减小。表 2为试验结束时成活鲍的规格。将各组鲍试验前后的壳长、壳宽和湿质量数据代入1.4的特定增长率公式,死亡鲍的增长率以0%计算在内,温度由低到高的壳长增长率分别为1.2%、0.5%、0.3%、0.1%和0.1%,壳宽增长率分别为1.2%、0.6%、0.3%、0.1%和0.1%,湿质量增长率分别为6.6%、3.3%、1.7%、1.1%和1.2%(图 1)。
表 2 各组鲍试验结束时的规格Table 2 Size of each group at the end of the experiment温度/℃ temperature 壳长/mm shell length 壳宽/mm shell width 湿质量/g wet weight 自然温度 natural temperature 11.62±1.22a 8.44±0.89a 3.21±0.11a 25 11.49±0.26b 8.39±0.25b 1.73±0.04b 28 10.95±0.34c 7.93±0.12c 0.96±0.02c 31 10.99±0.23c 7.95±0.27c 0.73±0.19c 32 11.31±0.64c 8.28±0.46c 0.75±0.23c ![]() 图 1 温度对皱纹盘鲍幼鲍壳长、壳宽和湿质量的影响Fig. 1 Effect of temperature on shell length, width and wet weight of juvenile H.discus hannai Ino
图 1 温度对皱纹盘鲍幼鲍壳长、壳宽和湿质量的影响Fig. 1 Effect of temperature on shell length, width and wet weight of juvenile H.discus hannai Ino2.2 皱纹盘鲍幼鲍对不同温度的耐受性
在日常管理期间观察到自然温度组的鲍吸附力和活力明显高于高温组,高温组鲍的软体部分所占比例随温度的升高也减小;试验结束后统计各组鲍的死亡率分别为2.2%、11.1%、43.3%、81.1%和86.7%。随着温度的升高死亡率显著升高(图 2)。
![]() 图 2 温度对皱纹盘鲍幼鲍死亡率的影响Fig. 2 Effect of temperature on mortality of juvenile H.discus hannai Ino
图 2 温度对皱纹盘鲍幼鲍死亡率的影响Fig. 2 Effect of temperature on mortality of juvenile H.discus hannai Ino3. 讨论
温度是影响鲍健康生长和生长快慢的重要决定因素。随着夏季气温的升高,提高鲍的抗逆性以使鲍安全度过较高水温是最直接且不易突破的技术手段,学者们针对这一问题采取过杂交育种的措施,“东优1号”杂色鲍和“大连1号”杂交鲍的培育成功在一定程度上提高了鲍的生长性能。游伟伟等[14]在对比温度对“东优1号”和其他3种地理群体杂色鲍的生长研究中发现,“东优1号”杂色鲍在各个温度下的生长情况高于其他3个地理群体,“大连1号”杂交鲍也比非杂交鲍的适温上限提高了4~5 ℃,生长率和成活率都高于父母本[13, 32],充分体现了杂种优势,这在一定程度上缓解了夏季死亡率高的局面,但仍不能满足市场需求。此外,前人从培育成功的杂交鲍中再继续筛选性状优良的鲍进行研究,该研究中5组鲍在试验开始时的壳长、壳宽和湿质量不存在显著性差异(表 1),而在试验结束时,壳长、壳宽和湿质量在对照组、25 ℃和28 ℃组间存在显著性差异(表 2),在28 ℃、31 ℃和32 ℃ 3组间无显著性差异,单因素方差分析表明,温度对鲍的生长影响很大,在高于其最适生长温度22 ℃后都有生长,但随温度的升高生长率降低,如湿质量的增长率在22 ℃时为6.6%,至25 ℃时为3.3%,28 ℃时只有1.7%,30 ℃以后下降到约1%。高绪生等[33]的研究结果显示,22 ℃时皱纹盘鲍幼鲍湿质量增长率只有2.6%,温度升高到26 ℃时其湿质量日增重率下降到1.7%,而该研究中皱纹盘鲍幼鲍在22 ℃时的湿质量增长率为6.6%,鲍湿质量日增重率为1.7%时的温度达到28 ℃,说明该试验选取的皱纹盘鲍幼鲍与高绪生等[34]选取的皱纹盘鲍稚鲍相比,在耐高温方面有所提高。
整体来看,杂交鲍随着温度的升高,死亡率明显增加。对照组温度随着气温变化在20~23 ℃间自然波动,其他4个控温组温度均设置为高于皱纹盘鲍最适生长温度的范围,温度达到28 ℃时该品系杂交鲍的成活率在50%以上,温度达到31 ℃时仍有接近20%的成活率,在此高温的基础上继续升温探索该杂交鲍的耐温性能,温度升至32 ℃维持1周后,该温度组成活鲍数为10只,占总数的11.1%,说明该杂交鲍经驯化后部分鲍能耐受的上限温度可达到32 ℃,这一温度高出其最适生长温度10 ℃。
综合上述两方面内容评价该品系杂交鲍的耐温性能,该杂交鲍在25 ℃时的日增长率可以达到传统皱纹盘鲍在22 ℃时的日增长率,且死亡率只有11.1%,适宜生长的上限温度升高了约3 ℃[33];温度升高至32 ℃的高温时仍有10%的个体成活,说明部分杂交鲍的耐受上限能够达到32 ℃。此结果为进行下一步的筛选工作奠定了良好的基础,笔者将从28 ℃以上高温中仍然成活下来的杂交鲍继续进行选育,选出更具耐高温潜力的品系。
在后续的试验中将选取不同地区选育的皱纹盘鲍进行对比,结合其在高温条件下的生长、死亡情况、体内的生理生化反应和温度骤变条件下热休克蛋白的表达情况[34-36],对比分析不同来源鲍对不良环境的耐受性,运用微卫星标记技术研究耐温群体和不耐温群体的基因序列差异[37-38]来辅助选取优良品种,为提高种群质量新品种的研究,对该杂交鲍的抗逆性进行评估。
-
![]()
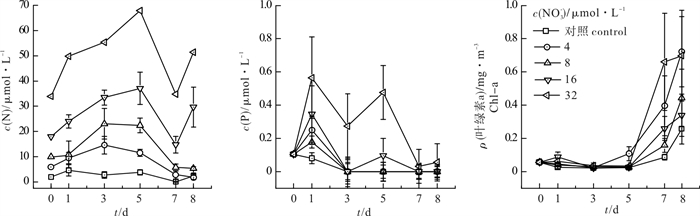
图 2 加N组叶绿素a和N、P浓度变化情况
Figure 2. Response of chlorophyll a to nutrients in N-added groups
![]()
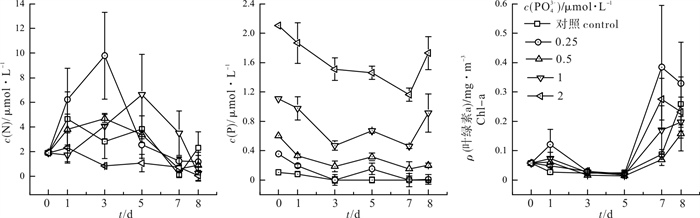
图 3 加P组叶绿素a和N、P浓度变化情况
Figure 3. Response of chlorophyll a to nutrients in P-added groups
![]()
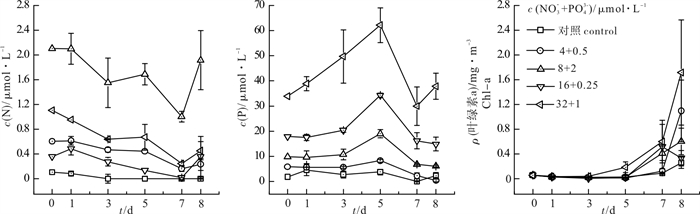
图 4 加N+P组叶绿素a和N、P浓度变化情况
Figure 4. Response of chlorophyll a to nutrients in N+P added groups
![]()
图 5 实验组叶绿素a质量浓度变化
不同字母表明有显著性差异(P < 0.05);a, b, c是用Duncan′s法排序
Figure 5. Temporal variation of chlorophyll a in experimental groups
Different letters indicate significant difference (P < 0.05);a, b and c are sorted by Duncan′s method.
表 1 培养站位理化环境因子初始值
Table 1 Initial physicochemical and environment factors values at S0 station
环境因子 environmental factor S0 水深/m depth -1 381 采样水层/m sampling water layer -5 透明度/m transparency 11.97 温度/℃ temperature 26.85 盐度 salinity 33.57 pH 8.21 ρ(溶解氧)/mg·L-1 DO 6.473 ρ(叶绿素a)/mg·m-3 Chl-a 0.06 c(NO2--N)/μmol·L-1 0.08 c(NO3--N)/μmol·L-1 1.81 c(NH4+-N)/μmol·L-1 0.74 c(PO43--P)/μmol·L-1 0.10 c(SiO32--Si)/μmol·L-1 1.34  下载: 导出CSV
下载: 导出CSV
表 2 营养盐加富实验方案
Table 2 Experiment design for nutrient enrichment
实验组
group No.添加元素
additional nutrientc(NO3--NaNO3)/μmol·L-1 c(PO43--NaH2PO4)/μmol·L-1 0 - - - 1 N 4 - 2 N 8 - 3 N 16 - 4 N 32 - 5 P - 0.25 6 P - 0.5 7 P - 1 8 P - 2 9 N,P 4 0.5 10 N,P 8 2 11 N,P 16 0.25 12 N,P 32 1 注:“-”表示不添加
Note:“-” means no addition.
下载: 导出CSV
-
[1] NEUNDORFER J V, KEMP W M. Nitrogen versus phosphorus enrichment of brackish waters: responses of the submersed plant Potamogeton perfoliatus and its associated algal community[J]. Mar Ecol Prog Ser, 1993, 94(1): 71-82.
[2] 唐松. 印度洋海水营养盐添加模拟实验中浮游植物生长的营养盐限制作用[J]. 海洋科学, 2010, 34(7): 34-40. [3] DUFOUR P, BERLAND B. Nutrient control of phytoplanktonic biomass in atoll lagoons and Pacific ocean waters: studies with factorial enrichment bioassays[J]. J Exp Mar Biol Ecol, 1999, 234(2): 147-166. doi: 10.1016/S0022-0981(98)00135-X
[4] THOMAS W H. Effect of ammonium and nitrate concentration on chlorophyll increases in natural tropical Pacific phytoplankton populations[J]. Limnol Oceanogr, 1970: 386-394.
[5] 陈纯, 李思嘉, 肖利娟, 等. 营养盐加富和鱼类添加对浮游植物群落演替和多样性的影响[J]. 生态学报, 2013, 33(18): 5777-5784. [6] 孙晓庆, 董树刚, 汤志宏. 营养盐和光照对浮游植物群落结构的影响[J]. 南方水产, 2008, 4(1): 1-9. doi: 10.3969/j.issn.2095-0780.2008.01.001 [7] 金杰, 刘素美. 海洋浮游植物对磷的响应研究进展[J]. 地球科学进展, 2013, 28(2): 253-261. [8] THINGSTAD T F, ZWEIFEL U L, RASSOULZADEGAN F. P limitation of heterotrophic bacteria and phytoplankton in the northwest Mediterranean[J]. Limnol Oceanogr, 1998, 43(1): 88-94. doi: 10.4319/lo.1998.43.1.0088
[9] 胡明辉, 杨逸萍, 徐春林, 等. 长江口浮游植物生长的磷酸盐限制[J]. 海洋学报(中文版), 1989, 11(4): 439-443. [10] 刘媛, 曹振锐, 黄邦钦, 等. 东、黄海典型海区浮游植物对营养盐添加的响应[J]. 厦门大学学报(自然科学版), 2004, 43(8): 147-152. [11] 王晓伟, 李纯厚, 戴明. 大亚湾湾口海域冬季浮游植物生长限制性元素研究[J]. 南方水产, 2007, 3(4): 26-31. doi: 10.3969/j.issn.2095-0780.2007.04.005 [12] 王晓伟, 李纯厚, 李占东, 等. 大亚湾海域春季浮游植物生长的限制性营养元素[J]. 中国水产科学, 2007, 14(5): 836-842. doi: 10.3321/j.issn:1005-8737.2007.05.019 [13] 王勇, 焦念志. 胶州湾浮游植物对营养盐添加的响应关系[J]. 海洋科学, 2002, 26(4): 8-13. doi: 10.3969/j.issn.1000-3096.2002.04.003 [14] 王玉珏, 洪华生, 王大志, 等. 台湾海峡上升流区浮游植物对营养盐添加的响应[J]. 生态学报, 2008, 28(3): 1321-1327. doi: 10.3321/j.issn:1000-0933.2008.03.049 [15] 朱艾嘉, 黄良民, 许战洲. 氮、磷对大亚湾大鹏澳海区浮游植物群落的影响Ⅰ. 叶绿素a与初级生产力[J]. 热带海洋学报, 2008, 27(1): 38-45. doi: 10.3969/j.issn.1009-5470.2008.01.008 [16] 晏宏, 孙立广, 刘晓东, 等. 近50年来南海西沙群岛海域气候异常的ENSO效应[J]. 热带海洋学报, 2010, 29(5): 29-35. doi: 10.3969/j.issn.1009-5470.2010.05.005 [17] 林洪瑛, 韩舞鹰. 南沙群岛海域营养盐对浮游植物生长的限制研究[C]//第七届全国海洋湖沼青年学者学术研讨会论文摘要集. [出版地不详]: 中国海洋湖沼学会, 2000: 14. [18] CHEN Y L, CHEN H, KARL D M, et al. Nitrogen modulates phytoplankton growth in spring in the South China Sea[J]. ContShelf Res, 2004, 24(4): 527-541.
[19] 车宏, 冉祥滨, 臧家业, 等. 南海南部海域夏季分粒级叶绿素a分布及影响因素[J]. 水生态学杂志, 2012, 33(4): 63-72. [20] LIU H, LI G, TAN Y, et al. Latitudinal changes (6° S-20° N) of summer ciliate abundance and species compositions in surface waters from the Java Sea to the South China Sea[J]. Acta Oceanologica Sinica, 2013, 32(4): 66-70. doi: 10.1007/s13131-013-0299-z
[21] REDFIELD A C. The biological control of chemical factors in the environment[J]. Am Sci, 1958, 46(3): 205-221.
[22] 李北兴, 王伟昌, 张大侠, 等. 正交设计和均匀设计在优化噻虫胺悬浮剂物理稳定性上的应用[J]. 中国农业科学, 2015, 48(2): 280-292. [23] 彭欣, 宁修仁, 孙军, 等. 南海北部浮游植物生长对营养盐的响应[J]. 生态学报, 2006, 26(12): 3959-3968. doi: 10.3321/j.issn:1000-0933.2006.12.006 [24] 唐松. 印度洋海水营养盐添加模拟实验中浮游植物生长的营养盐限制作用[J]. 海洋科学, 2010, 34(7): 34-40. [25] 徐燕青, 陈建芳, 高生泉, 等. 太平洋中西部海域浮游植物营养盐的潜在限制[J]. 生态学报, 2012, 32(2): 394-401. [26] MALLIN M A, PARSONS D C, JOHNSON V L, et al. Nutrient limitation and algal blooms in urbanizing tidal creeks[J]. J Exp Mar Biol Ecol, 2004, 298(2): 211-231. doi: 10.1016/S0022-0981(03)00360-5
[27] VUORIO K, LAGUS A, LEHTIMÄKI J M, et al. Phytoplankton community responses to nutrient and iron enrichment under different nitrogen to phosphorus ratios in the northern Baltic Sea[J]. J Exp Mar Biol Ecol, 2005, 322(1): 39-52. doi: 10.1016/j.jembe.2005.02.006
[28] 陈露, 戴明, 肖雅元, 等. 南沙群岛海域夏季营养盐对浮游植物生长的限制[J]. 生态学杂志, 2015, 34(5): 1342-1350. [29] 叶然, 刘艳云, 崔永平, 等. 东海营养盐结构的时空分布及其对浮游植物的限制[J]. 海洋与湖沼, 2015, 46(2): 311-320. [30] KIRKKALA T, HELMINEN H, ERKKILÄ A. Variability of nutrient limitation in the Archipelago Sea, SW Finland[M]//Eutrophication in planktonic ecosystems: food web dynamics and elemental cycling. Netherlands: Springer, 1998: 117-126.
[31] 王勇, 焦念志. 胶州湾浮游植物对营养盐添加的响应关系[J]. 海洋科学, 2002, 26(4): 8-13. doi: 10.3969/j.issn.1000-3096.2002.04.003 [32] 刘媛, 曹振锐, 黄邦钦, 等. 东、黄海典型海区浮游植物对营养盐添加的响应[J]. 厦门大学学报(自然科学版), 2004, 43(Z1): 147-152. doi: 10.3321/j.issn:0438-0479.2004.z1.035 [33] GRUBER N. The dynamics of the marine nitrogen cycle and its influence on atmospheric CO2 variations[M]//The ocean carbon cycle and climate. Netherlands: Springer, 2004: 97-148.
[34] 王保栋, 战闰, 藏家业. 黄海、东海浮游植物生长的营养盐限制性因素初探[J]. 海洋学报, 2003(S2): 190-195. [35] HECKY R E, KILHAM P. Nutrient limitation of phytoplankton in freshwater and marine environments: a review of recent evidence on the effects of enrichment. [J]. Limnol Oceanogr, 1988, 33(4): 796-822.
[36] 贾守伟, 史洁, 高会旺. 胶州湾营养盐限制作用空间差异和长期变化的数值研究[J]. 中国海洋大学学报(自然科学版), 2015, 45(5): 1-10.




计量
- 文章访问数: 2972
- HTML全文浏览量: 128
- PDF下载量: 1225
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
