
Preliminary analysis of relationship between mesh size of gillnet and body characteristics of target species in the South China Sea
-
摘要:
利用南海北部流刺网调查的相关资料对3种流刺网网目尺寸与主捕对象体型特征的关系进行了分析。结果表明,康氏马鲛(Scomberomorus commersoni)、中国鲳(Pampus chinensis)和龙头鱼(Harpodon nehereus)的体型特征为流刺网捕捞提供了有利的条件;相对体围值对应的渔获率变化曲线形状与刺网钟形选择性曲线相似;454尾主捕对象中仅1尾(占0.002%)相对体围值小于0.8;41尾相对体围值大于1.20,占0.09%;当相对体围值为1.0~1.1时,渔获率最高;白鲳刺网、龙头鱼刺网和马鲛刺网的最小网目尺寸分别为144 mm、35 mm和72 mm。建议结合选择性试验与体型特征研究,为制定南海区流刺网最小网目尺寸提供参考。
Abstract:We analyzed the relationship between mesh size of gillnet and body characteristics of target species for three kinds of gillnet fisheries. The results reveal that the morphological characteristics of Scomberomorus commersoni, Pampus chinensis and Harpodon nehereus are beneficial for gillnet fisheries. The catch rate curve of relative girth was similar to the selective curve of normal gillnet. Only 1 fish of 454 target species whose relative girth was less than 0.8, amounting to 0.002% of the total catch, and the relative girth of 41 fish was higher than 1.2, accounting for 0.09% of the total catch. The capture rate reached the maximum when the relative girth was 1.0~1.1. The minimal mesh sizes of three gillnet were 144 mm, 35 mm and 72 mm, respectively. It is suggested that the selectivity research should be combined with this study to provide references for promulgation of the minimal mesh size of gillnet in the South China Sea.
-
Keywords:
- gillnet /
- mesh size /
- target species /
- relationship analysis
-
刺网是一种被动型渔具,其捕捞原理是依靠网目的刺挂或网衣的缠络[1-2]。刺网捕捞是中国海洋渔业的一种重要作业方式。2012年,刺网的捕捞产量为2 728 031 t,仅次于拖网,占海洋捕捞总产量的21.5%[3]。漂流单片刺网(简称流刺网)是南海区分布最广、数量最多的刺网种类之一,其作业水层灵活、捕捞对象广泛,还有能耗少、成本低,且全年不休渔等特点[1]。近年来,随着燃油价格的飞涨,流刺网作业的优势更加突显,发展潜力较大。然而,南海流刺网最突出的问题是网目尺寸不断减小、作业中所使用的网片总数大为增加[4],这增加了近海渔业资源的捕捞压力,阻碍了流刺网渔业的可持续发展。
为了避免南海区流刺网的盲目发展,需出台相关的管理制度和标准。对于流刺网而言,规定最小网目尺寸(minimal mesh size)是最有效的管理措施。因为刺网是一种选择性极强的渔具,仅捕捞一定体长范围的种类[5]。迄今,已有许多学者致力于刺网与捕捞对象体型尺寸的相关研究。其中最为著名的有BARANOV的几何相似原理[6]和HAMLEY[7]的“20%理论”。REIS和PAWSON[8]对刺网网目与4个捕捞种类(体型相似)的体型特征进行了分析研究。然而,针对几种体型差异较大的种类与刺网网目尺寸关系的研究尚未见报道,而这对于制定刺网最小网目尺寸至关重要。目前,南海区对金线鱼刺网、马鲛流刺网和龙头鱼流刺网渔获组成、选择性、网目尺寸与主捕对象的体型特征关系已有相关报道[4-5, 9-12]。但将多种刺网网目尺寸与主捕对象体型特征进行比较和综合的研究还未见报道。文章在渔获方式为刺挂的基础上,对3种流刺网网目内径周长与主捕对象体周长的关系进行分析和比较,探讨南海流刺网的一些共性特点,为制定最小网目尺寸积累基础数据。
1. 材料与方法
1.1 数据的来源
该研究所用数据来自2012年~2013年南海流刺网渔获组成调查(表 1)。调查海域均为南海北部近海,流刺网为白鲳刺网、马鲛刺网和龙头鱼刺网,均为漂流单片刺网。3种流刺网的目标种类分别为中国鲳(Pampus chinensis)、康氏马鲛(Scomberomorus commersoni)和龙头鱼(Harpodon nehereus)。作业渔船分别来自沙扒港、博贺港和东平港。马鲛流刺网和龙头鱼流刺网的具体技术参数见杨炳忠等[11-12]。白鲳流刺网横向网目数为800目,纵向为22目,横目使用,网线材料为白胶丝。
表 1 南海流刺网渔获组成调查情况表Table 1. Gillnet survey in the South China Sea网具类型
gillnet调查时间
survey time渔船
fishing vessel渔场
fishing ground目标种类
target species网具主尺度
main dimension网目尺寸
mesh size白鲳刺网
P.chinensis gillnet2013年1月、2013年4月 粤西24313
(10.4 m,20.2 kW)阳江市沙扒镇近海
(111°30′ E~111°33′ E,21°24′ N~20°26′ N)中国鲳
P.chinensis72.00 m×4.27 m 190 mm 马鲛刺网
S.commersoni gillnet2012年4月、2012年11月 粤电渔42073
(33 m,132 kW)海南岛东北部
(111°08′~111°26′E,20°18′N~20°39′N)康氏马鲛
S.commersoni43.60 m×45.16 m 142 mm 龙头鱼刺网
H.nehereus gillnet2012年9月 粤阳东18111
(21 m,95 kW)珠海市三灶镇近海
(113°22′E~113°25′E,21°51′N~21°56′N)龙头鱼
H.nehereus68.08 m×5.33 m 43 mm 该研究仅针对3种流刺网与其目标种类的体型特征进行分析。样本量为中国鲳31尾、康氏马鲛123尾、龙头鱼300尾。调查中对目标种类进行生物学测量,包括渔获方式、渔获的体长(叉长)、体周长和体质量。
1.2 研究的基本假设
刺网对捕捞对象的渔获方式是研究网目尺寸和捕捞对象体型特征的基础。SPARRE等[13]把刺网的渔获方式分成4种:搁绊、刺入、契入和缠绕[14-15]。由于搁绊一般发生于一些特殊的捕捞对象(如头部有颚骨或棘的鱼类);刺入和契入并无本质区别,都是由于鱼体被网目卡住后捕获,只是挂刺部位有所差异。因此,该研究将流刺网的渔获方式分为刺挂和缠绕。在缠绕的渔获方式中,鱼体被网衣缠住而捕获,鱼体本身并不需要进入网目中[14]。该研究仅针对渔获方式为刺挂的渔获物进行。在此基础上,为了便于研究作如下基本假设:1)假设渔场中所有鱼类接触网具的概率相同;2)对于渔获方式为挂刺的种类,假设一尾鱼能否被捕获完全由其体型特征和网目尺寸的关系决定;3)假设流刺网对捕捞对象的最适捕捞体周长为网目内径周长。
1.3 研究方法
该研究以渔获方式为挂刺的中国鲳、龙头鱼和康氏马鲛为研究对象,分析网目尺寸和主捕对象体型特征的关系。重点分析主捕对象的最大体周长(体围)和网目内径周长的比例关系。统计并绘制相对体围所对应的渔获率变化曲线,分析相对体围值的上限值、下限值和峰值情况。统计相对体围值与HAMLEY[7]的“20%理论”吻合的比率,即相对体围值在0.8~1.2范围内所占的比例。相对体围的计算公式如下:
$$ G_{\mathrm{R}}=\frac{G_{\max }}{P_{\mathrm{mesh}}} $$ 式中GR为相对体围;Gmax为体围(mm);Pmesh为网目内径周长(mm)。
最小网目尺寸的探讨方法:利用目标种类体长与体周的回归关系,在渔获率曲线中把相对体围的峰值点视为选择性曲线的众数体长点(modal length);最后,利用目标种类的开捕规格,探讨3种流刺网的最小网目尺寸。
2. 结果
2.1 刺网的渔获种类组成
调查中发现,马鲛流刺网的渔获种类有20种,产量较大的有康氏马鲛、白卜鲔(Euthynnus yaito)和大眼金枪鱼(Thunnus obesus);龙头鱼刺网的渔获种类有龙头鱼、杜氏叫姑鱼(Johnius dussumieri)、金色小沙丁鱼(Sardinella aurita)等,共39种;白鲳刺网的渔获种类较少,仅见中国龙虾(Panulirus stimpsoni)、中国鲳和黄姑鱼(Nibea albiflora)。
2.2 目标种类的生物学特征
2.2.1 生物学参数
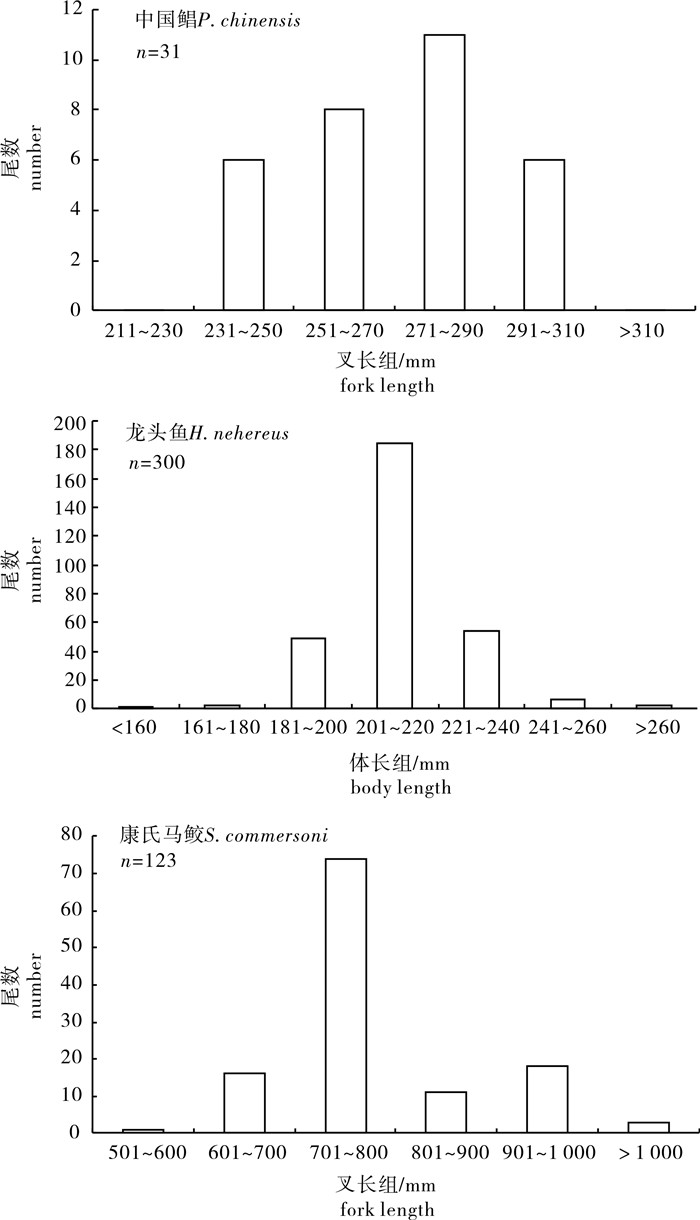
中国鲳生物学特征为:叉长232~301 mm,平均叉长(270.52±19.82)mm,最大体周长(体围)346~430 mm,平均体围(389.65±21.94)mm,体质量500~980 g,平均体质量(744.84±151.17)g;龙头鱼生物学特征为:体长110~289 mm,平均体长(211.62±15.28)mm,体围66~146 mm,平均体围(90.65±8.38)mm,体质量40~280 g,平均体质量(88.82±23.04)g;康氏马鲛的生物学特征为:叉长572~1 374 mm,平均叉长(786.25 ±106.54)mm,体质量1.55~15.48 kg,平均体质量(3.896 ±1.835)kg,体围270~504 mm,平均体围(332.95±45.11)mm。目标种类的体长(叉长)分布见图 1。
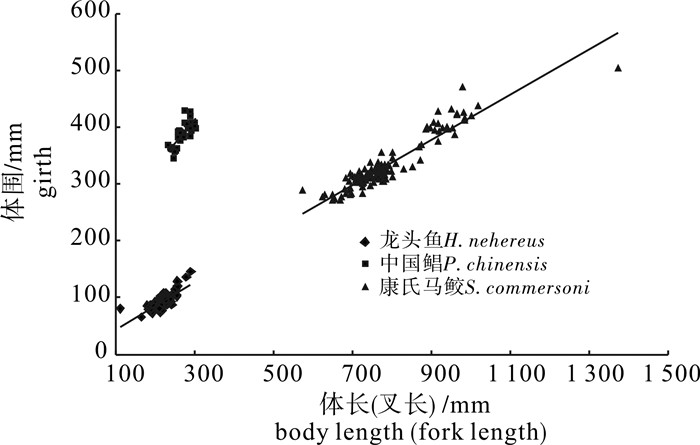
目标种类的体长和体围线性回归关系见图 2。回归方程如下:中国鲳G=0.840 1L+162.4(R2=0.575 6);龙头鱼G=0.409L+4.105 4(R2=0.556 9);康氏马鲛G=0.395 1L+22.285(R2=0.870 9);式中G为体围,L为体长(叉长)。
![]() 图 2 目标种类体长(叉长)和体围线性回归Figure 2. Relationship of body length (fork length) and girth of target species
图 2 目标种类体长(叉长)和体围线性回归Figure 2. Relationship of body length (fork length) and girth of target species2.2.2 体型特征与捕获部位
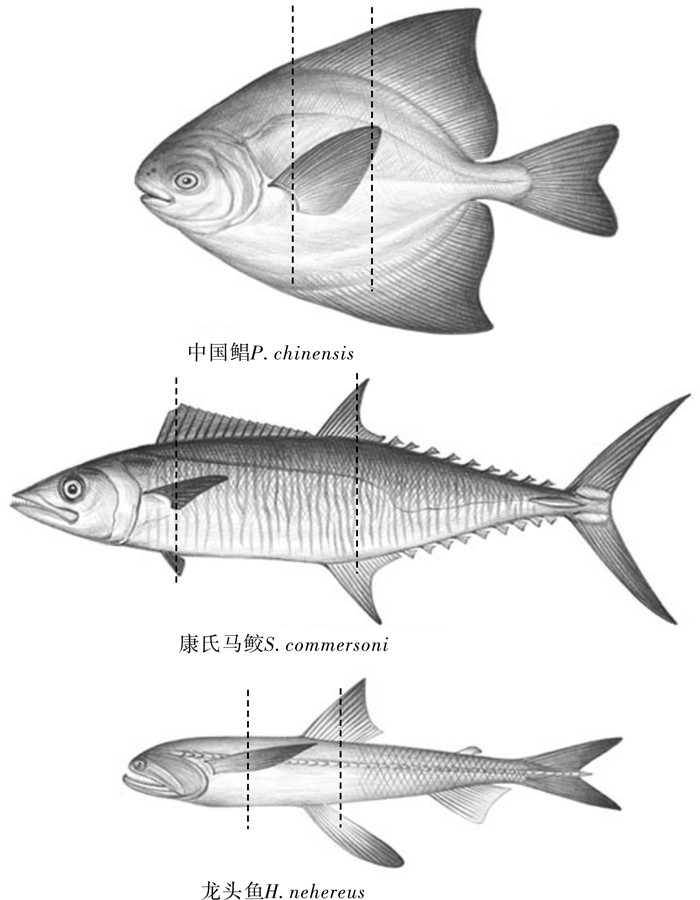
中国鲳、康氏马鲛和龙头鱼的体型见图 3。中国鲳体呈菱形,极侧扁,背鳍和臀鳍相对应;康氏马鲛体延长呈纺锤形,稍侧扁,背鳍2个,稍分离,第二背鳍与臀鳍同形;龙头鱼体柔软,体型接近纺锤型,略侧扁,背鳍起点与腹鳍起点相对。观察发现,中国鲳的捕捞“痕迹”主要位于胸鳍起点后,背鳍和臀鳍前的部位;康氏马鲛的捕捞“痕迹”分布的区域较大,从鳃盖骨后缘到第二背鳍起点处;龙头鱼的捕捞“痕迹”不明显。
2.3 体型特征与网目尺寸的关系
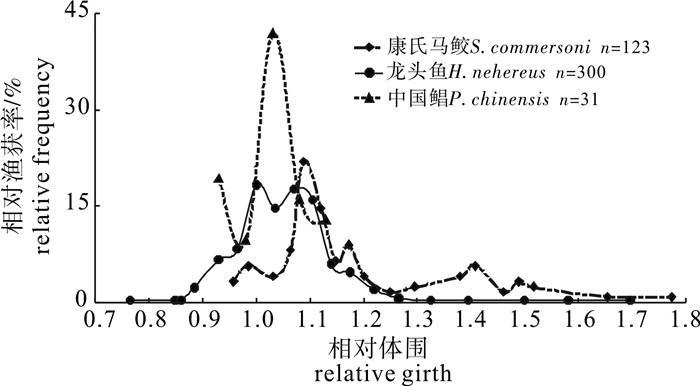
3种流刺网对目标种类的相对渔获率见图 4。从形状上看,3条曲线都呈现渔获率两边低、中间高的趋势,类似于典型的钟形选择性曲线。但龙头鱼和康氏马鲛的渔获率曲线右侧部分较长。
从渔获率曲线看,绝大多数的渔获相对体围分布于0.8~1.2:中国鲳的相对体围100%分布于0.8~1.2;龙头鱼的相对体围89.33%分布于0.8~1.2;康氏马鲛的相对体围77.24%分布于0.8~1.2。但在峰值上3条渔获率曲线有较大差异,中国鲳仅有1个峰值(占41.94%);龙头鱼存在2个峰值,分别占总渔获的18.33%和17.67%;康氏马鲛有多个峰值,但最高峰值仅1个,占总渔获的21.95%。
2.4 开捕规格与最小网目尺寸计算
3种流刺网的开捕规格和最小网目尺寸见表 2。目前,中国鲳尚无开捕规格的文献记录,文章暂时将其开捕规格定为140 mm。把最小网尺寸的参考点定于3种刺网对目标种类的峰值点(modal length)上,根据体周长与网目内径的关系,推导出白鲳刺网、龙头鱼刺网和马鲛刺网的最小网目尺寸分别为144 mm、35 mm和72 mm。
表 2 流刺网最小网目尺寸Table 2. Minimal mesh sizes of three gillnets种类
species体长(L)与体周(G)的关系
relationship of length and girth开捕规格/mm
minimal landing size相对体围峰值点
relative length体周长/mm
relative girth最小网目尺寸/mm
minimal mesh size中国鲳P.chinensis G=0.840 1L+162.4 140 1.03 288 144 龙头鱼H.nehereus G=0.409L+4.1054 160[21] 1.00 70 35 康氏马鲛S.commersoni G=0.395 1L+22.285 280[4] 1.09 144 72 3. 讨论
3.1 流刺网的作业性能比较
由于3种流刺网的主尺度、网目尺寸和作业渔场的水深区别较大,其渔获种类和数量存在较大差异。龙头鱼流刺网和白鲳流刺网网衣材料相同,均为白胶丝,均作业于水深小于20 m的近海渔场,但龙头鱼流刺网的渔获种类比白鲳流刺网的多,主要原因在于网目尺寸的差异。龙头鱼流刺网的网目尺寸仅为43 mm,远小于白鲳流刺网的190 mm。马鲛流刺网的渔获都是一些个体大、游泳能力强的种类,如康氏马鲛、大眼金枪鱼和扁舵鲣(Auxis thazard)等。一方面由于其网具主尺度大,另一方面与网衣的网线材料有关。马鲛流刺网的网衣材料是聚乙烯(8×3),网线粗、韧性强,能承受大型中上层种类试图穿越网目时产生的巨大冲击力。
3.2 流刺网网目尺寸与主捕对象体型的关系分析和探讨
目前,已有文献分别对马鲛流刺网和龙头鱼流刺网的渔获组成、网目与主捕对象的关系进行了初步分析[4, 11-12, 20]。文章以相对渔获率为依据,对南海3种流刺网网目与主捕对象体型的关系进行综合比较和分析,所得结论更加系统、全面和可靠:1)漂流单片刺网的网目对捕捞对象有高度的选择性,即与HAMLEY[7]的“20%理论”高度吻合;2)捕捞对象的体型直接影响刺网对其的渔获性能;3)目标种类的相对体围为1.0~1.1时相对渔获率最高。3种刺网454尾渔获中,仅42尾样品偏离“20%理论”(占9.25%);偏离“20%理论”的样品中仅1尾在下限区间(< 0.8),其余均在上限区间(>1.2)。从捕捞原理上分析,鱼体相对体围小于0.8时鱼可轻易穿越网目,渔获率很低;鱼体相对体围大于1.2时还有一部分鱼被捕获,因为其刺挂部位可位于最大体周长之前的某个部位。刺网对3种目标种类的有效捕捞区域均为鳃盖骨到背鳍与臀鳍起点处(图 3)。很显然,有效捕捞区域的大小(鱼体长轴方向)顺序为康氏马鲛>龙头鱼>中国鲳。因此,康氏马鲛的渔获率曲线右侧(相对体围>1.2)长于龙头鱼。由此可见,刺网渔获性能与渔获物的体型特征有密切关系。鱼体体周长接近或稍大于网目内径周长时刺网的捕捞效率最高,这与国外相关研究的结论一致[8]。
3.3 研究中长度特征的选择与分析
在刺网渔具性能和选择性研究中,估算和探讨网具对捕捞对象的最适捕捞体长是研究的关键。由于体长指标在渔业管理上有广泛应用[16-17],且其测量方便,很多流刺网选择性研究均采用体长作为研究变量[9, 18-19]。但是,实际捕捞过程中对于渔获方式为刺挂的种类,能否被捕获是由体周长(体围)和网目的内径周长决定的。因此,选择体围作为研究变量更为合理。鱼体的体围和体长存在一定线性关系,在研究中可以先估算最适捕捞体围,然后再推算出最适捕捞体长。
3.4 今后的研究方向
该文的研究结论可为制定南海区流刺网最小网目尺寸提供参考,但研究中仍然存在一些需要解决的问题:1)结果中体长和体周的相关性系数较小,这会影响用最适捕捞体周推算最适捕捞体长的结果;2)如何减少体围的测量误差。虽然该文对3种流刺网最小网目尺寸进行了初步的探讨,但结果还需要完善和补充。目标种类的最小可捕体长(开捕规格)是制定刺网最小网目尺寸的基础[22-23],但文章引用的开捕规格制定时间较早。其中,中国鲳目前尚无开捕规格报道;龙头鱼的开捕规格则来自东海区的相关研究。建议有关部门尽快出台南海区中国鲳、康氏马鲛和龙头鱼的最新开捕规格,然后采用多种不同网目尺寸的刺网进行研究,将选择性试验与该研究进行有机结合,为制定南海区流刺网最小网目尺寸提供更准确、科学的参考。
-
![]()
图 2 目标种类体长(叉长)和体围线性回归
Figure 2. Relationship of body length (fork length) and girth of target species
表 1 南海流刺网渔获组成调查情况表
Table 1 Gillnet survey in the South China Sea
网具类型
gillnet调查时间
survey time渔船
fishing vessel渔场
fishing ground目标种类
target species网具主尺度
main dimension网目尺寸
mesh size白鲳刺网
P.chinensis gillnet2013年1月、2013年4月 粤西24313
(10.4 m,20.2 kW)阳江市沙扒镇近海
(111°30′ E~111°33′ E,21°24′ N~20°26′ N)中国鲳
P.chinensis72.00 m×4.27 m 190 mm 马鲛刺网
S.commersoni gillnet2012年4月、2012年11月 粤电渔42073
(33 m,132 kW)海南岛东北部
(111°08′~111°26′E,20°18′N~20°39′N)康氏马鲛
S.commersoni43.60 m×45.16 m 142 mm 龙头鱼刺网
H.nehereus gillnet2012年9月 粤阳东18111
(21 m,95 kW)珠海市三灶镇近海
(113°22′E~113°25′E,21°51′N~21°56′N)龙头鱼
H.nehereus68.08 m×5.33 m 43 mm  下载: 导出CSV
下载: 导出CSV
表 2 流刺网最小网目尺寸
Table 2 Minimal mesh sizes of three gillnets
种类
species体长(L)与体周(G)的关系
relationship of length and girth开捕规格/mm
minimal landing size相对体围峰值点
relative length体周长/mm
relative girth最小网目尺寸/mm
minimal mesh size中国鲳P.chinensis G=0.840 1L+162.4 140 1.03 288 144 龙头鱼H.nehereus G=0.409L+4.1054 160[21] 1.00 70 35 康氏马鲛S.commersoni G=0.395 1L+22.285 280[4] 1.09 144 72
下载: 导出CSV
-
[1] 杨吝, 卢伙胜, 吴壮, 等. 南海区海洋渔具渔法[M]. 广州: 广东科技出版社, 2002: 13-32. [2] 黄锡昌. 海洋捕捞手册[M]. 北京: 农业出版社, 1990: 306-307. [3] 中华人民共和国农业部渔业局. 中国渔业统计年鉴2012 [M]. 北京: 中国农业出版社, 2013. [4] 杨炳忠, 杨吝, 谭永光, 等. 南海北部马鲛流刺网渔获组成初步分析[J]. 南方水产科学, 2013, 9 (1): 16-21. doi: 10.3969/j.issn.2095-0780.2013.01.003 [5] 张鹏, 杨吝, 张旭丰, 等. 南海北部金线鱼流刺网渔业情况分析[J]. 南方水产, 2008, 4(6): 101-107. doi: 10.3969/j.issn.2095-0780.2008.06.015 [6] HE P G. Behavior of marine fishes: capture processes and conservation challenges[M]. Oxford: Wiley-Blackwell, 2010: 183-203.
[7] HAMLEY J M. Review of gillnet selectivity [J]. J Fish Res Board Can, 1975, 32 (2): 1943-1969. doi: 10.1139/f75-233
[8] REIS E G, PAWSON M G. Fish morphology and estimating selectivity by gillnets [J]. Fish Res, 1999, 39(3): 263-273. doi: 10.1016/S0165-7836(98)00199-4
[9] 张鹏, 杨吝, 张旭丰, 等. 刺网网目尺寸对南海区金线鱼选择性研究[J]. 南方水产, 2005, 1(2): 61-66. doi: 10.3969/j.issn.2095-0780.2005.02.012 [10] 张鹏, 杨吝, 张旭丰, 等. 南海区金线鱼刺网网目选择性[J]. 中国水产科学, 2010, 17(5): 1085-1093. https://www.zhangqiaokeyan.com/academic-journal-cn_journal-fishery-sciences-china_thesis/0201261551000.html [11] 杨炳忠, 杨吝, 谭永光, 等. 马鲛体型特征与网目尺寸关系的初步探讨[J]. 南方水产科学, 2013, 9 (5): 120-125. doi: 10.3969/j.issn.2095-0780.2013.05.018 [12] 杨炳忠, 杨吝, 谭永光, 等. 龙头鱼流刺网网目捕获机理的初步研究[J]. 渔业现代化, 2013, 40(3): 46-50. doi: 10.3969/j.issn.1007-9580.2013.03.010 [13] SPARRE P, URSIN E, VENEMA S C. Introduction to tropical fish stock assessment (Part 1 manual)[J]. FAO Fisheries Technical Paper, 1989: 306. doi: 10.1067/mge.2000.107714
[14] 孙满昌, 张健, 钱卫国, 等. 渔具渔法选择性[M]. 北京: 中国农业出版社, 2004: 111-118. [15] 张健, 孙满昌. 刺网渔具选择性研究进展[J]. 中国水产科学, 2006, 16(6): 1040-1048. doi: 10.3321/j.issn:1005-8737.2006.06.027 [16] MILLAR R B, FRYER R J. Estimating of size-selection curves of towed gear, traps, nets and hooks [J]. Rev Fish Biol Fish, 1999, 9(1): 89-116. doi: 10.1023/A:1008838220001
[17] MILLAR R B. Estimation the size-selectivity of fishing gear by condition to trouser trawls [J]. J Amer Stat Assoc, 1992, 87(420): 962-968. doi: 10.1080/01621459.1992.10476250
[18] 李灵智, 黄洪亮, 王磊, 等. 东黄海区银鲳流刺网网目尺寸选择性研究[J]. 海洋渔业, 2010, 32(1): 89-94. doi: 10.3969/j.issn.1004-2490.2010.01.013 [19] 尤宗博, 李显森, 赵宪勇, 等. 蓝点马鲛大网目流刺网的选择性研究[J]. 水产学报, 2014, 38(2): 289-296. doi: 10.3724/SP.J.1231.2014.48950 [20] 杨炳忠, 杨吝, 谭永光, 等. 南海北部龙头鱼流刺网渔获组成初步分析[J]. 广东农业科学, 2013, 39(2): 99-102. doi: 10.3969/j.issn.1004-874X.2013.02.034 [21] 林龙山. 东海区龙头鱼数量分布及其环境特征[J]. 上海海洋大学学报, 2009, 18 (1): 66-71. https://www.zhangqiaokeyan.com/academic-journal-cn_detail_thesis/0201294218597.html [22] QUEIROLO D, GAETE E, AHUMADA M. Gillnet selectivity for Chilean hake (Merluccius gayi gayi Guichenot, 1848) in the bay of Valparaiso [J]. J Appl Ichthyol, 2013, 29(4): 775-781. doi: 10.1111/jai.12209
[23] PARK H H, MILLAR R B, BAE B S et al. Size selectivity of Korean flounder (Glyptocephalus stelleri) by gillnets and trammel nets using an extension of SELECT for experiments with differing mesh sizes[J]. Fish Res, 2011, 107(1/2/3): 196-200. doi: 10.1016/j.fishres.2010.10.020
-
期刊类型引用(7)
1. 梁敏,章雪琴. 水产品中大环内酯类药物残留检测技术的研究进展. 食品安全质量检测学报. 2021(02): 595-601 .  百度学术
百度学术
2. 黄珂,王旭峰,王强,赵东豪,杨宏亮,李刘冬. 分散固相萃取-超高效液相色谱串联质谱法同时测定水产品中4种PPCPs. 海洋环境科学. 2020(02): 296-301 . 百度学术
3. 刘敬先. HPLC法分析猪肉中泰乐菌素残留. 现代畜牧兽医. 2019(01): 12-16 . 百度学术
4. 周秀锦,张静,林维宣,王秋燕,周向阳. 质谱检测泰乐菌素的方法研究进展. 安徽农业科学. 2016(23): 31-34 . 百度学术
5. 陈惠兰,张晓燕,陈磊,殷耀,丁涛,朱文君,黄娟,杨雯筌. 蜂蜜中泰乐菌素A的稳定性研究. 中国蜂业. 2016(02): 48-50 . 百度学术
6. 周伟娥,张元,李伟青,李绍辉,蒋受军,李红娜,郑阳,张峰. 动物源性食品中大环内酯类药物前处理及检测方法研究进展. 食品与发酵工业. 2015(12): 241-247 . 百度学术
7. 申艳敏,刘振峰,李涛,任保增. 饲料药物添加剂泰乐菌素检测方法研究进展. 粮食与饲料工业. 2014(10): 63-65 . 百度学术
其他类型引用(2)
计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 9
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
