
Influence of nitrogen and phosphorus enrichment on phytoplankton community structure in Nansha area in summer
-
摘要:
为探讨夏季南沙海域氮(N)、磷(P)加富对浮游植物群落结构的影响,2013年夏季在南沙海域西南大陆架S1站(9°30′N,109°30′E)和曾母暗沙附近S2站(4°30′N,109°30′E)进行了多组添加N、P营养盐的现场加富实验。通过实验和数据分析对2个站位的浮游植物种类组成、丰度、多样性指数变化和群落更替情况进行了研究。结果表明:1)营养加富后浮游植物种类数增加;2)各加富组随着添加浓度增高,Shannon-Wiener多样性指数下降;3)从添加单一营养盐到复合营养盐组,浮游植物群落的优势类群出现了从甲藻到硅藻的演替;4)营养盐加富在短期内会使浮游植物丰度快速增加,但超过一定水平后,其促进作用减弱甚至抑制甲藻的生长;5)随着各组营养盐添加浓度的增高,加N、加P组的种群总丰度和甲藻门、硅藻门丰度变化不大,加N+P组的总丰度和硅藻门丰度显著上升,甲藻门丰度显著下降,说明N和P共同限制了浮游植物生长。
Abstract:To investigate the potential influence of nitrogen and phosphate on phytoplankton community structure, we conducted field experiments and enrichment incubation experiments in Nansha area [the southwest continental shelf and near the James shoal area: S1 (9°30′N, 109°30′E) and S2 (4°30′N, 109°30′E)] in summer of 2013. The results show that: 1) The number of phytoplankton species increased after enrichment; 2) Shannon-Wiener diversity index declined with increasing nutrient concentration; 3) Pyrrophyta and Bacillariophyta were the dominant species before and after the enrichment incubation, respectively. Pyrrophyta had more competitive advantages than Bacillariophyta in the oligotrophic region, leading to the shift from Pyrrophyta to Bacillariophyta; 4) Nutrient enrichment caused rapid growth of phytoplankton in a short time, but it inhibited phytoplankton growth when exceeding a certain level and the inhibition was associated with the phytoplankton species; 5) With increasing nutrients concentration in each group, the abundance of Pyrrophyta and Bacillariophyta in single N and single P groups had little change, but the abundance of Pyrrophyta in mixed N and P groups decreased significantly, and the abundance of Bacillariophyta increased significantly. Mixed N and P limited the phytoplankton growth.
-
Keywords:
- Nansha area /
- phytoplankton /
- nutrient enrichments in situ incubation /
- nitrogen /
- phosphorus
-
浮游植物是海洋生态系统中最重要的初级生产者,对海洋生态系统的物质循环和能量流动起关键作用,其组成与多样性反映了群落的结构类型和存在状态[1],其盛衰将直接或间接地制约海洋生产力的发展,从而对海洋生物资源的开发利用起重要影响[2]。开展营养盐加富实验中海洋浮游植物种类和生物量变化的研究,对探索南沙海域生态系统的结构和功能具有十分重要的意义。中国对于南沙海域浮游植物的报道较少,仅对该海域浮游植物群落的数量种类组成、多样性等群落特征进行了研究[3-6]。
南沙群岛位于南海南部,经纬度范围跨度大,3°37′N~11°57′N,108°00′E~118°00′E,具有典型热带海洋生态环境特征,受地形和季风驱动,中尺度涡频发,呈多涡环流结构[7-10],生境复杂,其复杂的生态环境孕育了多样的生物组成。目前尚未见到南沙海域营养盐加富实验对浮游植物群落生态特征影响的报道,为探讨该海域氮(N)和磷(P)对浮游植物群落结构特征的影响,笔者采用现场营养盐加富实验方法,在南沙海域进行了14个处理组的N、P营养盐加富培养实验,对西南大陆架区和曾母暗沙附近海域2个站位浮游植物的群落特征进行了较为深入的研究,以期弥补该海域营养盐加富对浮游植物群落影响的研究空白,并为南沙海域生物资源的持续开发利用和生态系统群落结构演替规律及其生物多样性变化研究提供基础数据。
1. 材料与方法
1.1 站点设置及理化环境因子初始值
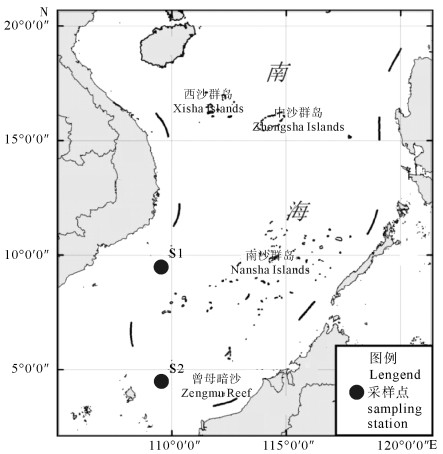
在2013年“南锋”号调查船出海调查期间,分别于2013年6月24日和7月1日进行现场表层采样,设置西南大陆架区的S1(9°30′N,109°30′E)和曾母暗沙附近的S2(4°30′N,109°30′E)2个站位[11](图 1)。各采样点的环境参数见表 1。2个站点的真光层内由于浮游植物生长的消耗,N、P营养盐的浓度经常处于很低的水平。
表 1 采样点理化环境因子初始值Table 1 Initial physicochemical properties of sampling stations站位
station水深/m
depth采样水层/m
sampling water layer温度/℃
temperature盐度
salinitypH ρ(溶解氧)/mg·L-1
DOρ(叶绿素a)/mg·m-3
Chl ac(NO2--N)/μmol·L-1 c(NO3--N)/μmol·L-1 c(NH4+-N)/μmol·L-1 c(PO43--P)/μmol·L-1 c(SiO32--Si)/μmol·L-1 S1 -1 583 -5 29.52 33.11 8.575 6.537 0 0.06 0.010 0 0.660 9 0.675 8 0.057 5 4.500 0 S2 -147 -5 30.12 33.08 8.585 7.446 5 0.09 0.063 9 0.284 1 0.015 0 0.011
(低于检测限)4.202 5 1.2 营养盐加富实验
实验以天然浮游植物群落为培养对象,选取N、P 2种添加元素,N、P之间比重参照Redfield比值设置为16:1,并结合南沙海域的寡营养盐等情况,每个实验站共设置14个营养盐加富实验组和1个未经加富处理的空白对照组(表 2),每组采水样2 L,平行双样,单营养和多营养加富同时进行培养4 d,以便相互验证。船载海水光照培养装置和加富实验操作详见文献[12]。
表 2 加富实验设计Table 2 Experimental design for enrichment实验组
group添加元素
additional nutrientc(NO3--NaNO3)/μmol·L-1 c(PO43--NaH2PO4)/μmol·L-1 0 - - - 1 N 4 - 2 N 8 - 3 N 16 - 4 N 32 - 5 P - 0.25 6 P - 0.5 7 P - 1 8 P - 2 9 N,P 4 0.25 10 N,P 8 0.5 11 N,P 16 1 12 N,P 32 2 13 N,P 32 1 14 N,P 32 4 注:“-”表示不添加
Note:“-” indicates no addition.1.3 样品采集与保存
水样采集、营养盐水样的处理与保存,以及叶绿素a测定、海水表层温、盐度的测定同文献[12~13]。2个站点分别于培养的第4天模拟实验结束后18:00~19:00采平行双样各400 mL用鲁哥氏液固定后暗处保存。参照文献[14~16]的方法浓缩沉淀24 h后浓缩并定容至25 mL,用计数框在AO倒置显微镜×200和×400倍下进行定性和定量分析。计算出浮游植物的丰度和生物量[17-18]。同时,现场测定pH、水温、溶解氧等初始理化指标。
1.4 样品分析与数据处理
优势度值(Y)[19]由式(1)来确定;浮游植物多样性指数的统计分析采用Shannon-Wiener多样性指数(H′)和Pielous均匀度指数(J′)[20], 分别用式(2)和(3)来计算。
$$ Y = \left( {{n_i}/N} \right) \times {f_i} $$ (1) $$ {H^\prime } = - \sum\limits_{i = 1}^S {{P_i}} {\log _2}{P_i} $$ (2) $$ J' = H'/{\log _2}S $$ (3) 式中ni为第i种的数量,N为采集样品中所有种类总个体数,S为样品中的种类总数,Pi为第i种的个体数与该样品总个体数比值(ni/N),f为该种在所采样品中出现的频率。
采用SPSS 21.0统计分析软件进行单因子方差(One-Way ANOVA)和LSD多重比较分析处理,以P<0.05作为显著差异水平,以P<0.01作为极显著差异水平。通过Origin 8.0完成图件绘制。
2. 结果与分析
2.1 营养盐和叶绿素a浓度变化
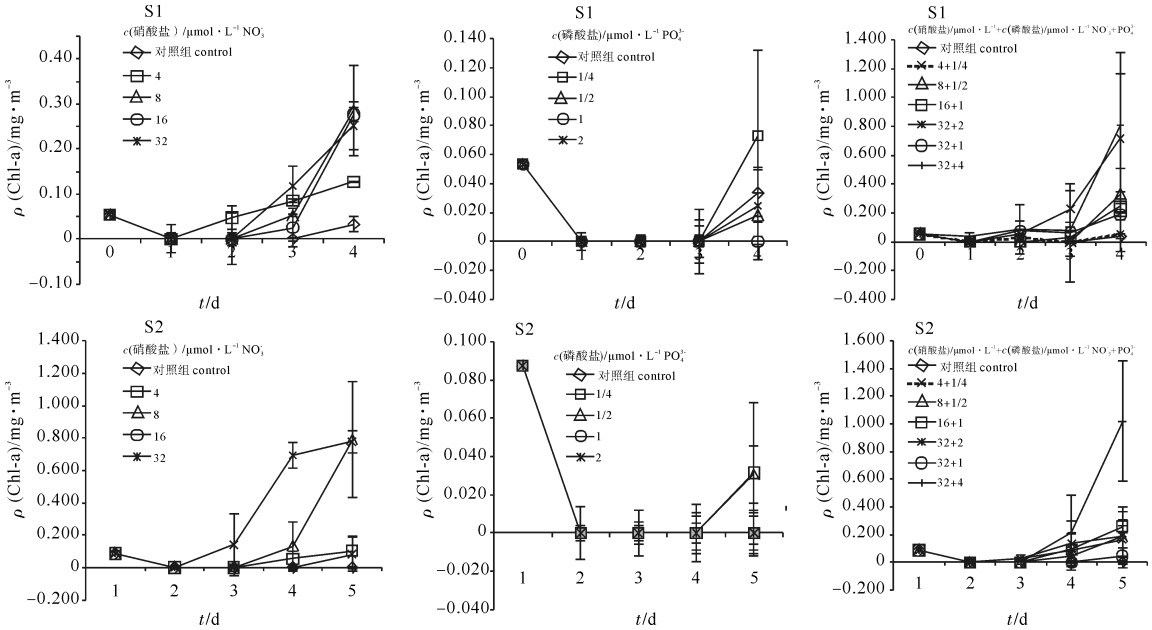
培养过程中,加P组的硝酸盐、磷酸盐及其浮游植物群落结构都变化不大,具体变化情况参照文献[13],说明加P对浮游植物群落结构无影响。加N组和加N+P组的硝酸盐浓度和磷酸盐的浓度变化随着叶绿素a质量浓度升高而显著降低。各站点叶绿素a质量浓度的变化情况见图 2。
![]() 图 2 各站点实验组的叶绿素a质量浓度变化Fig. 2 Temporal variation of chlorophyll a in experimental groups
图 2 各站点实验组的叶绿素a质量浓度变化Fig. 2 Temporal variation of chlorophyll a in experimental groups2.2 种类组成
原始水样两站位共鉴定浮游植物3门81种,加富培养4 d结束后种类数增加到3门170种(表 3)。硅藻门种类最多,47属132种;甲藻门5属39种;蓝藻门2属2种,占种类数的1.14%。出现种类较多的属为角藻属(Ceratium)15种、根管藻属(Rhizosolenia)15种、角刺藻属(Corethron)12种、角毛藻属(Chaetoceros)11种、圆筛藻属(Coscinodiscus)10种。
表 3 浮游植物种类组成及丰富度分布Table 3 Spieces composition and abundance of phytoplankton in different groups门
Phylum种类
species丰富度 abundance S1 S2 对照 control 氮 N 磷 P 氮+磷 N+P 对照 control 氮 N 磷 P 氮+磷 N+P 硅藻门 平滑角刺藻 Corethron laevis + + ++ ++ ++ + ++ Bacillarionphyta 窄隙角刺藻 C.affinis ++ + + ++ + + 大西洋角刺藻 C.atlanticus ++ ++ + ++ ++ ++ + 洛氏角刺藻 C.lorenzianus + + + + + + ++ 秘鲁角刺藻 C.peruvianus ++ +++ ++ + + + 悬垂角刺藻 C.pendulum ++ ++ + + + ++ ++ 扁面角刺藻 C.compressus ++ + + ++ ++ + +++ 金色角刺藻 C.aurivillei + + + ++ ++ + 均等角刺藻 C.aequatoriate ++ ++ + ++ + + + ++ 瘤面角刺藻 C.bacteriastroides ++ + + ++ ++ +++ ++ + 密连角刺藻 C.densus + ++ + + ++ + + 拟弯角刺藻 C.pseudocurvisetus ++ + ++ ++ 笔尖形根管藻 Rhizosolenia styliformis +++ +++ + + ++ + 伯氏根管藻 R.bergonii + ++ + + +++ ++ ++ 渐尖根管藻 R.acuminata + + ++ ++ ++ ++ +++ 距端根管藻 R.calcaravis ++ ++ ++ + + + + ++ 圆柱根管藻 R.cylindrus + + + ++ ++ ++ + 刚毛根管藻 R.setigera ++ + ++ ++ ++ +++ ++ 中华根管藻 R.sinensis + +++ ++ + + + 斯托根管藻 R.stolterfothii + ++ + + ++ + ++ 脆根管藻 R.fragilissima + + ++ + ++ + 斯氏根管藻 R.stolterforthii + + ++ ++ + ++ 卡斯根管藻 R.castracanei + ++ +++ + + 克菜根管藻 R.clevei + ++ ++ ++ ++ 翼根管藻 R.alata ++ + + ++ + + ++ 粗根管藻 R.robusta ++ + + ++ + 秘鲁角毛藻 Chaetoceros peruvianus ++ ++ + ++ + ++ 扭链角毛藻 C.tortissimus ++ ++ +++ ++ + 劳氏角毛藻 C.lorenzianus + + ++ + + + 窄隙角毛藻 C.affinis ++ + ++ ++ + ++ 密聚角毛藻 C.coarctatus ++ + + + + ++ 根状角毛藻 C.radicans + ++ + +++ ++ + 卡氏角毛藻 C.castracanei ++ ++ + ++ ++ + 长刺角毛藻 C.tetrastichon +++ +++ + ++ ++ + ++ 圆柱角毛藻 C.teres ++ ++ ++ +++ ++ + + 牟氏角毛藻 C.muelleri + ++ + ++ + ++ 旋链角毛藻 C.curvisetus + ++ ++ + + ++ + 弓束圆筛藻 Coscinodiscus astromphalus +++ ++ + ++ ++ + ++ 离心列圆筛藻 C.excentricus ++ ++ + + ++ + ++ ++ 虹彩圆筛藻 C.oculusiridis + + ++ + ++ + + 多束圆筛藻 C.divisus ++ + + + ++ 星脐圆筛藻 C.astromphalus + ++ + + ++ 高圆筛藻 C.nobilis + ++ + ++ +++ 辐射列圆筛藻 C.radiatus + +++ ++ + 辐射圆筛藻 C.radiatvs + + ++ ++ + 肾形圆筛藻 C.reniformis ++ ++ ++ + + ++ 斑点圆筛藻 C.punctulatus ++ + ++ ++ ++ + + 薄壁半管藻 Hemiaulus membranaceus + ++ + + + ++ 中华半管藻 H.sinensis ++ ++ + +++ ++ +++ ++ 霍氏半管藻 H.hauckii ++ ++ + + + ++ 印度半管藻 H.indicus + + + ++ ++ ++ ++ + 中国半管藻 H.chinensis ++ ++ ++ +++ + + 长耳盒形藻 Biddulphia aurita + + + + ++ + 中华盒形藻 B.sinensis ++ ++ ++ + +++ 网纹盒形藻 B.reticulata + ++ ++ + ++ 活动盒形藻 B.mobillensis + + ++ + ++ ++ ++ 长辐杆藻 Bacteriastrum elongatum ++ + + ++ + + ++ 变异辐杆藻 B.varians ++ + + ++ ++ ++ 透明辐杆藻 B.hyalinum ++ + ++ + + + 丛毛辐杆藻 B.comosum + + ++ + ++ ++ 地中海辐杆藻 B.mediterraneum ++ ++ + ++ + + + +++ 优美辐杆藻 B.delicatulum + + + + ++ ++ 双凹梯形藻 Climacodium biconcavum ++ ++ + ++ 佛朗梯形藻 C.frauenfeldianum + + + + +++ 五角星三角藻 Triceratium pentacrinus ++ + ++ ++ + 网纹三角藻 T.reticulum + ++ + + ++ 蜂窝三角藻 T.favus ++ ++ ++ +++ ++ 美丽三角藻 T.formosum + ++ ++ + + +++ 细纹三角藻 T.affine + + ++ ++ ++ + 克里星脐藻 Asteromphalus elegans + ++ ++ + + +++ 长卵面星脐藻 A.cleveanus + + ++ ++ 扇形星脐藻 A.flabellatus ++ + ++ + ++ 美丽星脐藻 A.elegans ++ + + ++ ++ ++ 日本星杆藻 A.japonica + ++ +++ + + + 美丽星杆藻 A.formosa + + + ++ 锤状中鼓藻 Bellerochea malleus ++ ++ ++ ++ 太阳双尾藻 Ditylum sol ++ + + ++ + 布氏双尾藻 D.brightwelli + ++ + +++ + 近缘斜纹藻 Pleurosigma affine ++ ++ + ++ + ++ 宽角斜纹藻 P.angulatum + ++ ++ + + 柔弱斜纹藻 P. delicatulum + ++ ++ ++ + ++ + 小斜纹藻 P.diminutum ++ + + ++ + 长斜纹藻 P.elongatum + +++ ++ ++ ++ + 楔形半盘藻 Hemidiscus cuneiformis + + ++ + + 哈德半盘藻 H.hardmannianus + + +++ ++ ++ 三舌辐裥藻 Actinoptychus trilingulatus ++ ++ ++ ++ ++ 球状辐裥藻 A.annulatus ++ + ++ + ++ 华美辐裥藻 A.splendens ++ ++ ++ ++ + + 大星纹藻 Asterolampra vanheurckii +++ ++ + + + 南方星纹藻 A.marylandica + + + ++ + + ++ 范氏星纹藻 A.vanheurckii + ++ ++ + 广卵罗氏藻 Roperia latiovala ++ ++ ++ + +++ ++ 方格罗氏藻 R.tesselata ++ + ++ ++ + 南极细柱藻 Leptocylindrus antarcticul ++ ++ ++ + ++ 地中海细柱藻 L.mediterraneus ++ ++ ++ + 亚德里亚细柱藻 L.adriaticus + + ++ ++ 丹麦细柱藻 L.danicus + ++ + + ++ 伏氏海毛藻 Thalassiothrix frauenfeldii + +++ ++ + 长海毛藻 T.longissima ++ ++ ++ + ++ + 细弱海链藻 Thalassiosira subtili + ++ ++ + + 透明海链藻 T.hyalina ++ + + ++ 诺登海链藻 T.nordenskioldii + ++ ++ + ++ + 圆海链藻 T.rotula ++ + ++ ++ + + 太阳漂流藻 Planktoniella sol ++ +++ ++ ++ 美丽漂流藻 P.formosa + + ++ ++ + ++ + 热带戈斯藻 Gossleriella tropica ++ ++ + ++ ++ ++ 几内亚藻 Guinardia flaccida ++ + + + +++ ++ 金色花盘藻 Chrysanthemodiscus floriatus ++ + ++ + ++ + 细弱明盘藻 Hyalodiscus subtilis ++ + ++ + ++ 优美施罗藻 Schroederella delicatula ++ + ++ +++ ++ + ++ 地中海指管藻 Dactyliosolen mediterraneus + + ++ 小环毛藻 Corethron hystrix ++ + + + 菱形海线藻 Thalassionema nitzschioides + ++ + ++ ++ + 巨盘藻 Ethmodiscus gazellae + + ++ + + ++ 北方劳德藻 Lauderia borealis ++ ++ + ++ + 短纹楔形藻 Licmophora abbreviata ++ ++ + ++ + ++ 串珠梯楔藻 Climacosphenia moniliger + ++ + ++ ++ 来都小环藻 Cyclotella ladogensis ++ +++ ++ + + 梅里小环藻 C.meneghiniana ++ ++ + 奇妙辐环藻 Actinocyclus alienus + + ++ ++ ++ 爱氏辐环藻 A.ehrenbergii ++ ++ + ++ + 中肋骨条藻 Skeletonema costatum ++ ++ ++ 尖刺菱形藻 Nitzschia pungens ++ ++ +++ + +++ + 大洋脆杆藻 Fragilaria oceanica ++ + + ++ ++ + 菱形海线藻 Thalassionema nitzschioides + ++ + + + 新月桥弯藻 Cymbella lunata + ++ ++ ++ ++ 七边乳头藻 Mastogonia heptagona ++ +++ ++ + + + 蛛网藻 Arachnoidiscus ehrenbergii + ++ ++ + ++ 印度扭鞘藻 Streptothece indica ++ ++ 翼茧形藻 Amphiprora alata ++ + ++ + 单点条纹藻 Striatella unipunctata ++ + + + ++ 掌状冠盖藻 Stephanopyxis palmeriana + + ++ 甲藻门 钩梨甲藻 Pyrocystis hamulus ++ ++ + Dinophyta 新月梨甲藻 P.lunula ++ + + ++ ++ + ++ 夜光梨甲藻 P.noctiluca + ++ +++ +++ 浅弧梨甲藻 P.gerbaulti +++ + ++ +++ 梭梨甲藻 P.fusiformis + + + + 丰满多甲藻 Peridinium grande + + ++ 大洋多甲藻 P.oceanicum ++ ++ ++ + + + 优美多甲藻 P.elegans ++ +++ ++ + ++ 扁平多甲藻 P.depressum +++ + + + ++ 大多甲藻 P.grande + ++ + ++ 链状多甲藻 P.catenatum + + + 偏转角藻 Ceratium deflexum ++ ++ + + + 扭角藻 C.contortum + ++ ++ ++ ++ 三角角藻 C.tripos ++ + + +++ +++ 羊头角藻 C.arietinum ++ + ++ + ++ 瘤状角藻 C.gibberum ++ ++ + ++ + + 二裂角藻 C.biceps ++ +++ +++ ++ 腊台角藻 C.candelebrum + + ++ + 长梭角藻 C.extensum ++ + ++ ++ ++ 镰角藻 C.falcatum + + + + + ++ 叉状角藻 C.furca + + +++ ++ + 网纹角藻 C.hexacanthum +++ + ++ + 长角角藻 C.macroceros ++ ++ ++ 波状角藻 C.trichoceros ++ + ++ + 马西里亚角藻 C.massilense +++ ++ ++ 二齿双管藻 Amphisolenia bidentata ++ ++ ++ + 三叉双管藻 A.trinax ++ + +++ + + 四齿双管藻 A.schauinslandii ++ ++ ++ ++ 四叶鸟尾藻 Ornithocercus steinii + ++ +++ ++ + + 美丽鸟尾藻 O.splendidus ++ + + + + 方鸟尾藻 O.quadratus + + + + ++ 华美鸟尾藻 O.magnificus ++ + + ++ ++ + 扁甲藻 Pyrophacus horologicum + + + + + 具尾鳍藻 Dinophysis caudata ++ ++ ++ 勇士鳍藻 D.miles + + + ++ ++ 刺尖甲藻 Oxyitoxum scolopax ++ + ++ + + + 蓝藻门 近膜质席藻 Phormidium submenbranaceum ++ + + + Cyanophyta 胞内植生藻 Richelia intracellularis + + + + + + 注:+. 少见种;+ +. 常见种;+ + +. 优势种
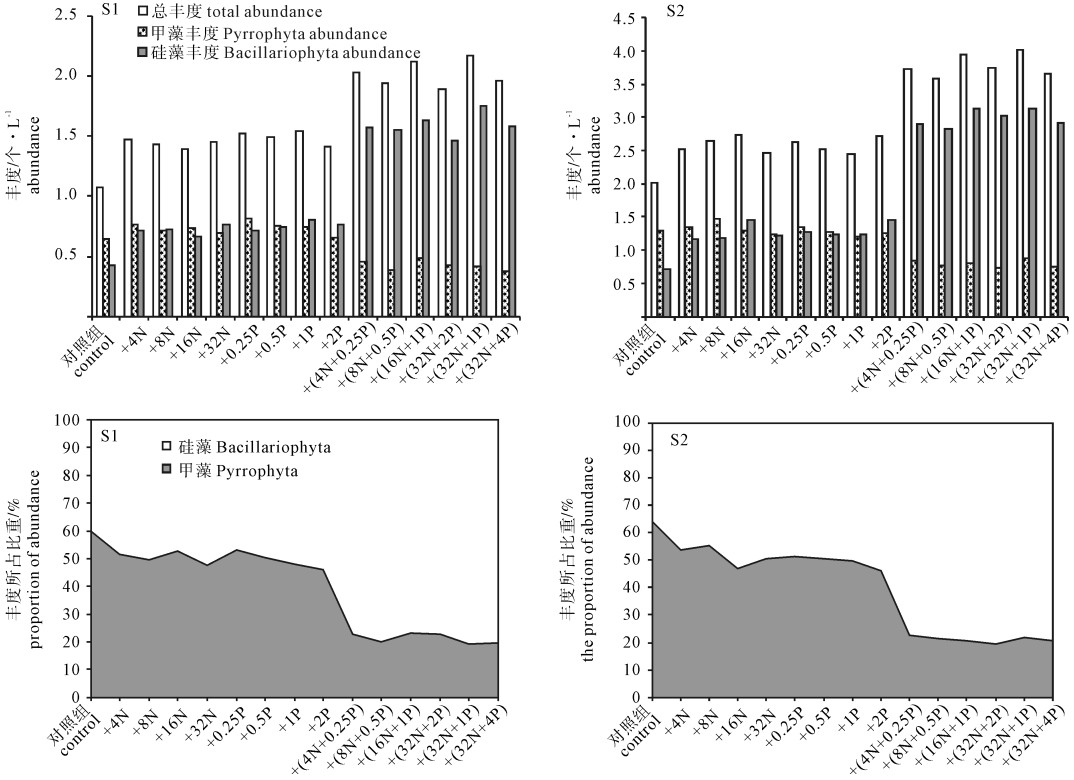
Note:+. rare species;+ +. common species;+ + +. dominant species与对照组相比,两站位单一添加组的总丰度平均值增加幅度为1.07~1.47、2.01~2.59。硅藻门丰度的增加幅度分别为0.43~0.73和0.72~1.27,甲藻门随添加浓度增加变化不大(图 3)。
![]() 图 3 两站位浮游植物丰度的变化情况Fig. 3 Temporal variation of abundance of phytoplankton in experimental groups
图 3 两站位浮游植物丰度的变化情况Fig. 3 Temporal variation of abundance of phytoplankton in experimental groups2.3 丰度
与对照组和单一添加组对比,加N+P组的硅藻门丰度均显著增高(P < 0.05),增加幅度分别为0.43~1.59和0.72~2.99,甲藻门丰度均显著下降(P < 0.05),下降幅度分别为0.73~0.43和1.31~0.80。
对照组的甲藻丰度所占比重稍大于硅藻,加N、P组都是甲藻和硅藻丰度持平各占约50%,添加N+P后硅藻丰度所占比重约是甲藻的3.8倍。
综上,加N和加P后甲藻与硅藻密度均出现增长但变化并不显著(P>0.05),且硅藻门和甲藻门的丰度和所占比重持平。加N+P后浮游植物总丰度、硅藻门丰度及其丰度所占比重明显上升,甲藻门反之。
2.4 优势种
根据各实验组浮游植物在群落出现的丰度和对生物量的贡献程度确定各组的优势种。
两站位的空白对照组浮游植物优势种均以甲藻为主(66.7%),其种类主要是优美多甲藻(P.elegans)、浅弧梨甲藻(P.gerbaulti)、叉状角藻(C.furca)、马西里亚角藻(C.massilense)、三角角藻(C.tripos)、二裂角藻(C.biceps)、夜光梨甲藻(P.noctiluca)、三叉双管藻(A.trinax)。而硅藻次之(33.3%),其主要优势种为笔尖形根管藻(R.styliformis)、长刺角毛藻(C.tetrastichon)、中华半管藻(H.sinensis)、尖刺菱形藻(N.pungens)。各营养加富组的浮游植物优势类群以硅藻为主(80.4%),而甲藻次之(19.6%)(表 4)。说明加富后硅藻更适宜生存,浮游植物发生了从甲藻到硅藻的演替。
表 4 浮游植物优势种组成Table 4 Composition of dominant species of phytoplanktonS1站 Station 1 S2站位 Station 2 对照 control 氮 N 磷 P 氮+磷 N+P 对照 control 氮 N 磷 P 氮+磷 N+P 优美多甲藻
P.elegans秘鲁角刺藻
C.peruvianus七边乳头藻
M.heptagona哈德半盘藻
H.hardmannianus三角角藻
C.tripos瘤面角刺藻
C.bacteriastroides尖刺菱形藻
N.pungens中华盒形藻
B.sinensis浅弧梨甲藻
P.gerbaulti中华根管藻
R.sinensis笔尖形根管藻
R. tyliformis美丽漂流藻
P.formosa二裂角藻
C.biceps蜂窝三角藻
T.favus中华半管藻
H.sinensis地中海辐杆藻
B.mediterraneum叉状角藻C.furca 弓束圆筛藻
C.astromphalus来都小环藻
C.ladogensis圆柱角毛藻
C.teres夜光梨甲藻
P.noctiluca伯氏根管藻
R. bergonii广卵罗氏藻
R.latiovala扁面角刺藻
C.compressus马西里亚角藻
C.massilense大星纹藻
A.vanheurckii伏氏海毛藻
T.frauenfeldii辐射列圆筛藻
C.radiatus三叉双管藻
A.trinax布氏双尾藻
D.brightwelli刚毛根管藻
R. setigera佛朗梯形藻
C.frauenfeldianum笔尖形根管藻
R.styliformis长刺角毛藻
C.tetrastichon扭链角毛藻
C.tortissimus中国半管藻
H.chinensis中华半管藻
H.sinensis根状角毛藻
C. radicans几内亚藻
G.flaccida渐尖根管藻
R.acuminata长刺角毛藻
C.tetrastichon长斜纹藻
P.elongatum二裂角藻
C.biceps日本星杆藻
A.japonica尖刺菱形藻
N.pungens三叉双管藻
A.trinax美丽三角藻
T.formosum克里星脐藻
A.elegans马西里亚角藻
C.massilense四叶鸟尾藻
O.steinii优美施罗藻
S.delicatula三角角藻
C.tripos高圆筛藻
C.nobilis优美多甲藻
P.elegans卡斯根管藻
R.castracanei浅弧梨甲藻
P.gerbaulti夜光梨甲藻
P.noctiluca叉状角藻
C.furca2.5 Shannon-Wiener多样性与均匀度指数
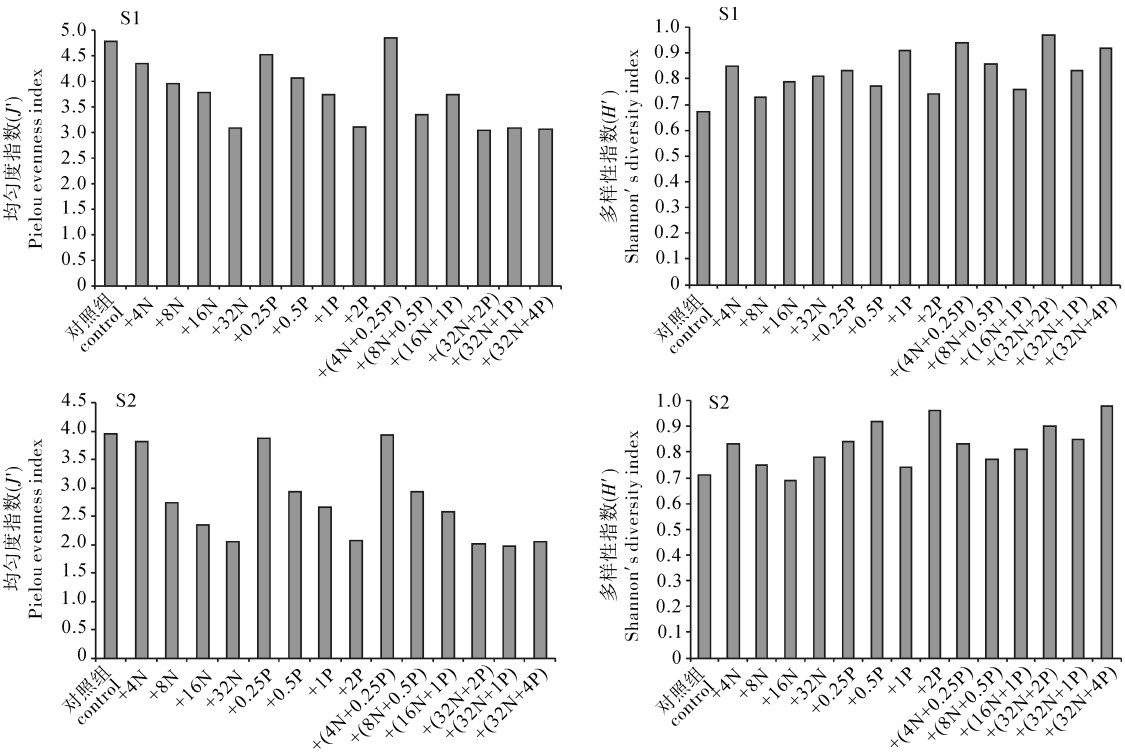
Shannon-Wiener多样性指数对于维持水生态稳定性具有关键性作用,所以常用来评价海域生物种类组成的多样性水平。物种的均匀度也是反映群落结构特征的一个重要指标,是群落是否成熟和稳定的特征之一[21]。
S1、S2站位空白对照组H′依次为4.78、3.95,J′依次为0.67、0.71。与对照组相比,加N组和加P组、加N+P组的H′均显著下降,但是J′均稍上升。
加富组多样性指数介于3.72~4.83之间,S1站位平均值为3.76,最高值出现在4N+0.25P组,为4.86;S2站位平均值为2.80,最高值出现在4N+0.25P组,为3.93(图 4)。
![]() 图 4 两站位浮游植物Shannon-Wiener多样性和均匀度指数的变化Fig. 4 Temporal variation of Shannon-Wiener diversity index and evenness index of phytoplankton
图 4 两站位浮游植物Shannon-Wiener多样性和均匀度指数的变化Fig. 4 Temporal variation of Shannon-Wiener diversity index and evenness index of phytoplanktonJ′介于0.67~0.98,平均值为0.825。各加富组随着单一和复合营养盐添加浓度升高,J′均无显著性变化(P>0.05)。通常以均匀度>0.3作为浮游植物多样性较好的标准进行综合评价[22],结果显示在水体中分布比较均匀,该海域浮游植物群落加富后优势种发生了逆转性的演替。
3. 讨论
3.1 营养盐加富对种类组成的影响
对照组反映该海区浮游植物种类丰富,这与海区受陆源污染物质和人类影响较小有关。浮游植物优势种以甲藻为主,这与李开枝等[4]的报道类似。一般而言,在0~75 m的水层采样,甲藻在细胞数量和种类上比硅藻更有优势。南沙海域浮游植物的调查分析[23]表明,浮游植物种类组成不同于近岸海域,优势种为耐高盐、大洋暖水性的甲藻和硅藻类,甲藻种类大大高于中国其他海区,反映出热带、外洋性特征。因为在该营养盐缺乏的海区,甲藻较硅藻有更高的生态适应性[13, 24-26],因此,其相对更具有群落竞争优势。和原始水样相比,加富后实验组种类数增加,说明加富促进了各种浮游植物的生长。陈纯等[1]在研究营养加富对浮游植物群落演替和多样性影响时发现,适量添加营养盐产生的上行效应可缓解浮游植物之间的竞争程度,提高浮游植物群落的多样性。
3.2 营养盐加富对种群丰度和群落结构的影响
加富前,表现为甲藻比硅藻的丰度值大。在越靠近外海营养盐缺乏的海区,甲藻在低营养盐浓度的环境下适应能力较硅藻强,因此甲藻类在浮游植物中更具有竞争优势,所占比重越高,占50%以上[14],硅藻反之。加富后,单一的添加N、P对浮游植物群落结构无显著影响,共同添加N+P则显著促进硅藻门丰度快速上升,甲藻反之。说明该海域是N+P共同限制了浮游植物生长,这和彭欣等[14]在南海北部的加富实验结果一致。
综上发现,营养盐加富的干扰导致了南沙海域夏季浮游植物群落出现了从甲藻到硅藻的演替。说明外界环境条件的改变较群落初始物种组成对浮游植物群落的演替顺序更具决定性作用[27]。在南海北部加富培养前后也发现了此现象[14]。说明加富前水体中较低的营养盐浓度限制了浮游植物的生长,营养盐加富在短期内引起浮游植物丰度的快速增加。随着营养盐的消耗和浮游动物的牧食作用,当第4天丰度增加到一定程度后稳定不变甚至开始降低,浮游植物丰度降低到一定量时,由于竞争和牧食作用得到缓解,浮游植物现存量在短期内会有回升趋势,所以营养加富组的浮游植物现存量存在一定波动[1]。
当N+P浓度超过一定水平后,其促进作用减弱甚至还会抑制某些物种的生长,该抑制作用在主要优势种上得以充分体现[28]。甲藻这种赤潮藻类通过上下洄游移动到所需要的营养盐浓度的水层才能正常生长。甲藻数量的根本性下降也可能是因为容器的大小、高低等外在实验装置条件或者其引起的“瓶壁效应”影响了浮游植物的种群变化。
实验结束时发现添加N/P比为32 : 1组最适合硅藻生长,也验证了高N/P比有利于硅藻生长[29]。硅藻类在浮游植物中占优势,这与ERLA等[30]在美国Galveston湾和CHEN等[31]培养报道结果一致,营养盐比重不失衡且营养盐浓度高的海区,氮磷盐的联合效应促使硅酸盐充分为硅藻硅质外壳的形成提供物质基础,有利于硅藻生长。由于现场加富实验改变了浮游植物的环境条件,如增加了营养盐利用率,定期的扰动(晃瓶)等已经被证实也可促进硅藻的旺发[32]。
3.3 营养盐加富对浮游植物Shannon-Wiener多样性指数的影响
空白对照组多样性指数较高,两站点各加富组均随着添加浓度升高,H′连续明显下降(P<0.05)。孙晓庆等[28]在探讨营养盐和光照对浮游植物群落结构影响时发现营养盐加富使浮游植物H′明显下降,说明浮游植物群落结构发生变化,有生态异常和不稳定趋向,是对环境信息的反馈。
此次南沙海域加富实验的浮游植物群落的H′为3.72~4.83,S1、S2两站位的平均值分别为3.76和2.80。2002年宋星宇等[3]夏季在该海区大面调查发现浮游植物的H′平均值为4.01,中南半岛东南部存在浮游植物分布高值区和低值区,这和S1、S2站位出现高低差异保持一致。整体来看,研究海域浮游植物的群落结构相对较为简单,H′较低。主要是因为在对营养盐的竞争过程中,营养盐的供应对浮游植物群落结构起重要的调节作用,出现了以硅藻为主的优势种。营养盐添加的浓度越高,其比例离Redfield比值越远,浮游植物物种就越少[33]。南沙群岛海区浮游植物的分布与海区地形、水文、营养盐、浮游动物的摄食等因素对浮游植物群落结构和多样性变化也会产生一定影响。
-
![]()
图 2 各站点实验组的叶绿素a质量浓度变化
Figure 2. Temporal variation of chlorophyll a in experimental groups
![]()
图 3 两站位浮游植物丰度的变化情况
Figure 3. Temporal variation of abundance of phytoplankton in experimental groups
![]()
图 4 两站位浮游植物Shannon-Wiener多样性和均匀度指数的变化
Figure 4. Temporal variation of Shannon-Wiener diversity index and evenness index of phytoplankton
表 1 采样点理化环境因子初始值
Table 1 Initial physicochemical properties of sampling stations
站位
station水深/m
depth采样水层/m
sampling water layer温度/℃
temperature盐度
salinitypH ρ(溶解氧)/mg·L-1
DOρ(叶绿素a)/mg·m-3
Chl ac(NO2--N)/μmol·L-1 c(NO3--N)/μmol·L-1 c(NH4+-N)/μmol·L-1 c(PO43--P)/μmol·L-1 c(SiO32--Si)/μmol·L-1 S1 -1 583 -5 29.52 33.11 8.575 6.537 0 0.06 0.010 0 0.660 9 0.675 8 0.057 5 4.500 0 S2 -147 -5 30.12 33.08 8.585 7.446 5 0.09 0.063 9 0.284 1 0.015 0 0.011
(低于检测限)4.202 5  下载: 导出CSV
下载: 导出CSV
表 2 加富实验设计
Table 2 Experimental design for enrichment
实验组
group添加元素
additional nutrientc(NO3--NaNO3)/μmol·L-1 c(PO43--NaH2PO4)/μmol·L-1 0 - - - 1 N 4 - 2 N 8 - 3 N 16 - 4 N 32 - 5 P - 0.25 6 P - 0.5 7 P - 1 8 P - 2 9 N,P 4 0.25 10 N,P 8 0.5 11 N,P 16 1 12 N,P 32 2 13 N,P 32 1 14 N,P 32 4 注:“-”表示不添加
Note:“-” indicates no addition.
下载: 导出CSV
表 3 浮游植物种类组成及丰富度分布
Table 3 Spieces composition and abundance of phytoplankton in different groups
门
Phylum种类
species丰富度 abundance S1 S2 对照 control 氮 N 磷 P 氮+磷 N+P 对照 control 氮 N 磷 P 氮+磷 N+P 硅藻门 平滑角刺藻 Corethron laevis + + ++ ++ ++ + ++ Bacillarionphyta 窄隙角刺藻 C.affinis ++ + + ++ + + 大西洋角刺藻 C.atlanticus ++ ++ + ++ ++ ++ + 洛氏角刺藻 C.lorenzianus + + + + + + ++ 秘鲁角刺藻 C.peruvianus ++ +++ ++ + + + 悬垂角刺藻 C.pendulum ++ ++ + + + ++ ++ 扁面角刺藻 C.compressus ++ + + ++ ++ + +++ 金色角刺藻 C.aurivillei + + + ++ ++ + 均等角刺藻 C.aequatoriate ++ ++ + ++ + + + ++ 瘤面角刺藻 C.bacteriastroides ++ + + ++ ++ +++ ++ + 密连角刺藻 C.densus + ++ + + ++ + + 拟弯角刺藻 C.pseudocurvisetus ++ + ++ ++ 笔尖形根管藻 Rhizosolenia styliformis +++ +++ + + ++ + 伯氏根管藻 R.bergonii + ++ + + +++ ++ ++ 渐尖根管藻 R.acuminata + + ++ ++ ++ ++ +++ 距端根管藻 R.calcaravis ++ ++ ++ + + + + ++ 圆柱根管藻 R.cylindrus + + + ++ ++ ++ + 刚毛根管藻 R.setigera ++ + ++ ++ ++ +++ ++ 中华根管藻 R.sinensis + +++ ++ + + + 斯托根管藻 R.stolterfothii + ++ + + ++ + ++ 脆根管藻 R.fragilissima + + ++ + ++ + 斯氏根管藻 R.stolterforthii + + ++ ++ + ++ 卡斯根管藻 R.castracanei + ++ +++ + + 克菜根管藻 R.clevei + ++ ++ ++ ++ 翼根管藻 R.alata ++ + + ++ + + ++ 粗根管藻 R.robusta ++ + + ++ + 秘鲁角毛藻 Chaetoceros peruvianus ++ ++ + ++ + ++ 扭链角毛藻 C.tortissimus ++ ++ +++ ++ + 劳氏角毛藻 C.lorenzianus + + ++ + + + 窄隙角毛藻 C.affinis ++ + ++ ++ + ++ 密聚角毛藻 C.coarctatus ++ + + + + ++ 根状角毛藻 C.radicans + ++ + +++ ++ + 卡氏角毛藻 C.castracanei ++ ++ + ++ ++ + 长刺角毛藻 C.tetrastichon +++ +++ + ++ ++ + ++ 圆柱角毛藻 C.teres ++ ++ ++ +++ ++ + + 牟氏角毛藻 C.muelleri + ++ + ++ + ++ 旋链角毛藻 C.curvisetus + ++ ++ + + ++ + 弓束圆筛藻 Coscinodiscus astromphalus +++ ++ + ++ ++ + ++ 离心列圆筛藻 C.excentricus ++ ++ + + ++ + ++ ++ 虹彩圆筛藻 C.oculusiridis + + ++ + ++ + + 多束圆筛藻 C.divisus ++ + + + ++ 星脐圆筛藻 C.astromphalus + ++ + + ++ 高圆筛藻 C.nobilis + ++ + ++ +++ 辐射列圆筛藻 C.radiatus + +++ ++ + 辐射圆筛藻 C.radiatvs + + ++ ++ + 肾形圆筛藻 C.reniformis ++ ++ ++ + + ++ 斑点圆筛藻 C.punctulatus ++ + ++ ++ ++ + + 薄壁半管藻 Hemiaulus membranaceus + ++ + + + ++ 中华半管藻 H.sinensis ++ ++ + +++ ++ +++ ++ 霍氏半管藻 H.hauckii ++ ++ + + + ++ 印度半管藻 H.indicus + + + ++ ++ ++ ++ + 中国半管藻 H.chinensis ++ ++ ++ +++ + + 长耳盒形藻 Biddulphia aurita + + + + ++ + 中华盒形藻 B.sinensis ++ ++ ++ + +++ 网纹盒形藻 B.reticulata + ++ ++ + ++ 活动盒形藻 B.mobillensis + + ++ + ++ ++ ++ 长辐杆藻 Bacteriastrum elongatum ++ + + ++ + + ++ 变异辐杆藻 B.varians ++ + + ++ ++ ++ 透明辐杆藻 B.hyalinum ++ + ++ + + + 丛毛辐杆藻 B.comosum + + ++ + ++ ++ 地中海辐杆藻 B.mediterraneum ++ ++ + ++ + + + +++ 优美辐杆藻 B.delicatulum + + + + ++ ++ 双凹梯形藻 Climacodium biconcavum ++ ++ + ++ 佛朗梯形藻 C.frauenfeldianum + + + + +++ 五角星三角藻 Triceratium pentacrinus ++ + ++ ++ + 网纹三角藻 T.reticulum + ++ + + ++ 蜂窝三角藻 T.favus ++ ++ ++ +++ ++ 美丽三角藻 T.formosum + ++ ++ + + +++ 细纹三角藻 T.affine + + ++ ++ ++ + 克里星脐藻 Asteromphalus elegans + ++ ++ + + +++ 长卵面星脐藻 A.cleveanus + + ++ ++ 扇形星脐藻 A.flabellatus ++ + ++ + ++ 美丽星脐藻 A.elegans ++ + + ++ ++ ++ 日本星杆藻 A.japonica + ++ +++ + + + 美丽星杆藻 A.formosa + + + ++ 锤状中鼓藻 Bellerochea malleus ++ ++ ++ ++ 太阳双尾藻 Ditylum sol ++ + + ++ + 布氏双尾藻 D.brightwelli + ++ + +++ + 近缘斜纹藻 Pleurosigma affine ++ ++ + ++ + ++ 宽角斜纹藻 P.angulatum + ++ ++ + + 柔弱斜纹藻 P. delicatulum + ++ ++ ++ + ++ + 小斜纹藻 P.diminutum ++ + + ++ + 长斜纹藻 P.elongatum + +++ ++ ++ ++ + 楔形半盘藻 Hemidiscus cuneiformis + + ++ + + 哈德半盘藻 H.hardmannianus + + +++ ++ ++ 三舌辐裥藻 Actinoptychus trilingulatus ++ ++ ++ ++ ++ 球状辐裥藻 A.annulatus ++ + ++ + ++ 华美辐裥藻 A.splendens ++ ++ ++ ++ + + 大星纹藻 Asterolampra vanheurckii +++ ++ + + + 南方星纹藻 A.marylandica + + + ++ + + ++ 范氏星纹藻 A.vanheurckii + ++ ++ + 广卵罗氏藻 Roperia latiovala ++ ++ ++ + +++ ++ 方格罗氏藻 R.tesselata ++ + ++ ++ + 南极细柱藻 Leptocylindrus antarcticul ++ ++ ++ + ++ 地中海细柱藻 L.mediterraneus ++ ++ ++ + 亚德里亚细柱藻 L.adriaticus + + ++ ++ 丹麦细柱藻 L.danicus + ++ + + ++ 伏氏海毛藻 Thalassiothrix frauenfeldii + +++ ++ + 长海毛藻 T.longissima ++ ++ ++ + ++ + 细弱海链藻 Thalassiosira subtili + ++ ++ + + 透明海链藻 T.hyalina ++ + + ++ 诺登海链藻 T.nordenskioldii + ++ ++ + ++ + 圆海链藻 T.rotula ++ + ++ ++ + + 太阳漂流藻 Planktoniella sol ++ +++ ++ ++ 美丽漂流藻 P.formosa + + ++ ++ + ++ + 热带戈斯藻 Gossleriella tropica ++ ++ + ++ ++ ++ 几内亚藻 Guinardia flaccida ++ + + + +++ ++ 金色花盘藻 Chrysanthemodiscus floriatus ++ + ++ + ++ + 细弱明盘藻 Hyalodiscus subtilis ++ + ++ + ++ 优美施罗藻 Schroederella delicatula ++ + ++ +++ ++ + ++ 地中海指管藻 Dactyliosolen mediterraneus + + ++ 小环毛藻 Corethron hystrix ++ + + + 菱形海线藻 Thalassionema nitzschioides + ++ + ++ ++ + 巨盘藻 Ethmodiscus gazellae + + ++ + + ++ 北方劳德藻 Lauderia borealis ++ ++ + ++ + 短纹楔形藻 Licmophora abbreviata ++ ++ + ++ + ++ 串珠梯楔藻 Climacosphenia moniliger + ++ + ++ ++ 来都小环藻 Cyclotella ladogensis ++ +++ ++ + + 梅里小环藻 C.meneghiniana ++ ++ + 奇妙辐环藻 Actinocyclus alienus + + ++ ++ ++ 爱氏辐环藻 A.ehrenbergii ++ ++ + ++ + 中肋骨条藻 Skeletonema costatum ++ ++ ++ 尖刺菱形藻 Nitzschia pungens ++ ++ +++ + +++ + 大洋脆杆藻 Fragilaria oceanica ++ + + ++ ++ + 菱形海线藻 Thalassionema nitzschioides + ++ + + + 新月桥弯藻 Cymbella lunata + ++ ++ ++ ++ 七边乳头藻 Mastogonia heptagona ++ +++ ++ + + + 蛛网藻 Arachnoidiscus ehrenbergii + ++ ++ + ++ 印度扭鞘藻 Streptothece indica ++ ++ 翼茧形藻 Amphiprora alata ++ + ++ + 单点条纹藻 Striatella unipunctata ++ + + + ++ 掌状冠盖藻 Stephanopyxis palmeriana + + ++ 甲藻门 钩梨甲藻 Pyrocystis hamulus ++ ++ + Dinophyta 新月梨甲藻 P.lunula ++ + + ++ ++ + ++ 夜光梨甲藻 P.noctiluca + ++ +++ +++ 浅弧梨甲藻 P.gerbaulti +++ + ++ +++ 梭梨甲藻 P.fusiformis + + + + 丰满多甲藻 Peridinium grande + + ++ 大洋多甲藻 P.oceanicum ++ ++ ++ + + + 优美多甲藻 P.elegans ++ +++ ++ + ++ 扁平多甲藻 P.depressum +++ + + + ++ 大多甲藻 P.grande + ++ + ++ 链状多甲藻 P.catenatum + + + 偏转角藻 Ceratium deflexum ++ ++ + + + 扭角藻 C.contortum + ++ ++ ++ ++ 三角角藻 C.tripos ++ + + +++ +++ 羊头角藻 C.arietinum ++ + ++ + ++ 瘤状角藻 C.gibberum ++ ++ + ++ + + 二裂角藻 C.biceps ++ +++ +++ ++ 腊台角藻 C.candelebrum + + ++ + 长梭角藻 C.extensum ++ + ++ ++ ++ 镰角藻 C.falcatum + + + + + ++ 叉状角藻 C.furca + + +++ ++ + 网纹角藻 C.hexacanthum +++ + ++ + 长角角藻 C.macroceros ++ ++ ++ 波状角藻 C.trichoceros ++ + ++ + 马西里亚角藻 C.massilense +++ ++ ++ 二齿双管藻 Amphisolenia bidentata ++ ++ ++ + 三叉双管藻 A.trinax ++ + +++ + + 四齿双管藻 A.schauinslandii ++ ++ ++ ++ 四叶鸟尾藻 Ornithocercus steinii + ++ +++ ++ + + 美丽鸟尾藻 O.splendidus ++ + + + + 方鸟尾藻 O.quadratus + + + + ++ 华美鸟尾藻 O.magnificus ++ + + ++ ++ + 扁甲藻 Pyrophacus horologicum + + + + + 具尾鳍藻 Dinophysis caudata ++ ++ ++ 勇士鳍藻 D.miles + + + ++ ++ 刺尖甲藻 Oxyitoxum scolopax ++ + ++ + + + 蓝藻门 近膜质席藻 Phormidium submenbranaceum ++ + + + Cyanophyta 胞内植生藻 Richelia intracellularis + + + + + + 注:+. 少见种;+ +. 常见种;+ + +. 优势种
Note:+. rare species;+ +. common species;+ + +. dominant species
下载: 导出CSV
表 4 浮游植物优势种组成
Table 4 Composition of dominant species of phytoplankton
S1站 Station 1 S2站位 Station 2 对照 control 氮 N 磷 P 氮+磷 N+P 对照 control 氮 N 磷 P 氮+磷 N+P 优美多甲藻
P.elegans秘鲁角刺藻
C.peruvianus七边乳头藻
M.heptagona哈德半盘藻
H.hardmannianus三角角藻
C.tripos瘤面角刺藻
C.bacteriastroides尖刺菱形藻
N.pungens中华盒形藻
B.sinensis浅弧梨甲藻
P.gerbaulti中华根管藻
R.sinensis笔尖形根管藻
R. tyliformis美丽漂流藻
P.formosa二裂角藻
C.biceps蜂窝三角藻
T.favus中华半管藻
H.sinensis地中海辐杆藻
B.mediterraneum叉状角藻C.furca 弓束圆筛藻
C.astromphalus来都小环藻
C.ladogensis圆柱角毛藻
C.teres夜光梨甲藻
P.noctiluca伯氏根管藻
R. bergonii广卵罗氏藻
R.latiovala扁面角刺藻
C.compressus马西里亚角藻
C.massilense大星纹藻
A.vanheurckii伏氏海毛藻
T.frauenfeldii辐射列圆筛藻
C.radiatus三叉双管藻
A.trinax布氏双尾藻
D.brightwelli刚毛根管藻
R. setigera佛朗梯形藻
C.frauenfeldianum笔尖形根管藻
R.styliformis长刺角毛藻
C.tetrastichon扭链角毛藻
C.tortissimus中国半管藻
H.chinensis中华半管藻
H.sinensis根状角毛藻
C. radicans几内亚藻
G.flaccida渐尖根管藻
R.acuminata长刺角毛藻
C.tetrastichon长斜纹藻
P.elongatum二裂角藻
C.biceps日本星杆藻
A.japonica尖刺菱形藻
N.pungens三叉双管藻
A.trinax美丽三角藻
T.formosum克里星脐藻
A.elegans马西里亚角藻
C.massilense四叶鸟尾藻
O.steinii优美施罗藻
S.delicatula三角角藻
C.tripos高圆筛藻
C.nobilis优美多甲藻
P.elegans卡斯根管藻
R.castracanei浅弧梨甲藻
P.gerbaulti夜光梨甲藻
P.noctiluca叉状角藻
C.furca
下载: 导出CSV
-
[1] 陈纯, 李思嘉, 肖利娟, 等. 营养盐加富和鱼类添加对浮游植物群落演替和多样性的影响[J]. 生态学报, 2013, 33(18): 5777-5784. doi: 10.5846/stxb201304010584 [2] 张婷, 李林, 宋立荣. 熊河水库浮游植物群落结构的周年变化[J]. 生态学报, 2009, 29(6): 2971-2979. doi: 10.3321/j.issn:1000-0933.2009.06.025 [3] 宋星宇, 黄良民, 钱树本, 等. 南沙群岛邻近海区春夏季浮游植物多样性研究[J]. 生物多样性, 2002, 10(3): 258-268. doi: 10.3321/j.issn:1005-0094.2002.03.002 [4] 李开枝, 郭玉洁, 尹健强, 等. 南沙群岛海区秋季浮游植物物种多样性及数量变化[J]. 热带海洋学报, 2005, 24(3): 25-30. doi: 10.3969/j.issn.1009-5470.2005.03.004 [5] 戴明, 刘华雪, 廖秀丽, 等. 初秋南沙群岛海域网采浮游植物群落特征及其与环境因子的关系[J]. 应用生态学报, 2013, 24(12): 3553-3560. [6] 李冬融, 戴鑫烽, 陆斗定, 等. 2012年夏季南海西北部网采浮游植物群落结构[J]. 海洋学研究, 2014, 32(3): 87-96. doi: 10.3969/j.issn.1001-909X.2014.03.012 [7] 方文东, 郭忠信, 黄羽庭. 南海南部海区的环流观测研究[J]. 科学通报, 1997, 42(21): 2264-2271. doi: 10.1360/csb1997-42-21-2264 [8] CHU P C, FAN C, LOZANO C J, et al. An airborne expendable bathythermograph survey of the South China Sea, May 1995[J]. J Geophys Res Oceans (1978-2012), 1998, 103(C10): 21637-21652. doi: 10.1029/98JC02096
[9] 苏纪兰. 南海环流动力机制研究综述[J]. 海洋学报: 中文版, 2006, 27(6): 1-8. doi: 10.3321/j.issn:0253-4193.2005.06.001 [10] 黄企洲, 方文东, 陈荣裕. 南沙群岛海区西南季风变化与表层流结构和演变关系的探讨[J]. 热带海洋学报, 2001, 20(1): 18-26. doi: 10.3969/j.issn.1009-5470.2001.01.003 [11] LIU H, LI G, TAN Y, et al. Latitudinal changes (6°S-20°N) of summer ciliate abundance and species compositions in surface waters from the Java Sea to the South China Sea[J]. Acta Oceanol Sin, 2013, 32(4): 66-70. doi: 10.1007/s13131-013-0299-z
[12] 陈露, 戴明, 肖雅元, 等. 南沙群岛海域夏季营养盐对浮游植物生长的限制[J]. 生态学杂志, 2015, 34(5): 1342-1350. doi: 10.13292/j.1000-4890.20150311.063 [13] 郭永坚, 罗昭林, 朱长波, 等. 水产养殖对流沙湾浮游植物群落特征的影响[J]. 南方水产科学, 2015, 11(2): 57-65. doi: 10.3969/j.issn.2095-0780.2015.02.008 [14] 彭欣, 宁修仁, 孙军, 等. 南海北部浮游植物生长对营养盐的响应[J]. 生态学报, 2006, 26(12): 3959-3968. doi: 10.3321/j.issn:1000-0933.2006.12.006 [15] 唐松. 印度洋海水营养盐添加模拟实验中浮游植物生长的营养盐限制作用[J]. 海洋科学, 2010, 34(7): 34-40. [16] 徐燕青, 陈建芳, 高生泉, 等. 太平洋中西部海域浮游植物营养盐的潜在限制[J]. 生态学报, 2012, 32(2): 394-401. doi: 10.5846/stxb201012141778 [17] JUN S, DONGYAN L, SHUBEN Q. Study on phytoplankton biomassⅠ. Phytoplankton measurement biomass from cell volume or plasma volume [J]. Acta Oceanol Sin, 1999, 2: 9. https://xueshu.baidu.com/usercenter/paper/show?paperid=a8f468c892bcc0a5d154bf7178f84be4&site=xueshu_se
[18] HILLEBRAND H, DURSELEN C D, KIRSCHTEL D, et al. Biovolume calculation for pelagic and benthic microalgae[J]. J Phycol, 1999, 35(2): 403-424. doi: 10.1046/j.1529-8817.1999.3520403.x
[19] 张伟, 李纯厚, 贾晓平, 等. 大亚湾混凝土鱼礁和铁质鱼礁附着生物群落结构的季节变化[J]. 南方水产科学, 2015, 11(1): 9-17. doi: 10.3969/j.issn.2095-0780.2015.01.002 [20] PIELOU E C. An introduction to mathematical ecology[M]. New York: Wiley-Interscience, 1966: 1-286. doi: 10.2307/3799632
[21] 孙军, 刘东艳, 魏皓, 等. 琉球群岛邻近海域浮游植物多样性的模糊综合评判[J]. 海洋与湖沼, 2001, 32(4): 445-453. doi: 10.3321/j.issn:0029-814X.2001.04.014 [22] 徐兆礼, 陈亚瞿. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系[J]. 生态学杂志, 1989, 8(4): 13-15. https://xueshu.baidu.com/usercenter/paper/show?paperid=2c2a4932f8939d1409e44c01d7a927c8&site=xueshu_se [23] 中国科学院南沙综合科学考察队. 南沙群岛及其邻近海区海洋生物多样性研究Ⅱ[C]. 北京: 海洋出版社, 1996: 11-27. https://xueshu.baidu.com/usercenter/paper/show?paperid=4350d903d67fb203b5672e1115c3a9f6&site=xueshu_se [24] 朱根海, 宁修仁, 蔡昱明, 等. 南海浮游植物种类组成和丰度分布的研究[J]. 海洋学报: 中文版, 2003, 25(S2): 8-23. https://xueshu.baidu.com/usercenter/paper/show?paperid=fd9b3e4daad5ffc8b28a9ea8c702eabf&site=xueshu_se&hitarticle=1 [25] 顾新根, 袁骐, 杨蕉文, 等. 长江口羽状锋海区浮游植物的生态研究[J]. 中国水产科学, 1995, 2(1): 1-15. doi: 10.3321/j.issn:1005-8737.1995.01.006 [26] 李瑞香, 朱明远, 王宗灵, 等. 东海两种赤潮生物种间竞争的围隔实验[J]. 应用生态学报, 2003, 14(7): 1049-1054. doi: 10.3321/j.issn:1001-9332.2003.07.004 [27] 黄伟, 朱旭宇, 曾江宁, 等. 氮磷比对浙江近岸浮游植物群落结构影响的实验研究[J]. 海洋学报: 中文版, 2012, 34(5): 128-138. [28] 孙晓庆, 董树刚, 汤志宏. 营养盐和光照对浮游植物群落结构的影响[J]. 南方水产, 2008, 4(1): 1-9. doi: 10.3969/j.issn.2095-0780.2008.01.001 [29] HEIL C A, REVILLA M, GLIBERT P M, et al. Nutrient quality drives differential phytoplankton community composition on the southwest Florida shelf[J]. Limnol Oceanogr, 2007, 52(3): 1067-1078. doi: 10.4319/lo.2007.52.3.1067
[30] ERLA B E, LUMSDEN S E, PINCKNEY J L. Nutrient pulsing as a regulator of phytoplankton abundance and community composition in Galveston Bay, Texas[J]. J Exp Mar Biol Ecol, 2004, 303(2): 197-220. doi: 10.1016/j.jembe.2003.11.016
[31] CHEN Y L, CHEN H, KARL D M, et al. Nitrogen modulates phytoplankton growth in spring in the South China Sea[J]. Cont Shelf Res, 2004, 24(4): 527-541. doi: 10.1016/j.csr.2003.12.006
[32] 康燕玉, 梁君荣, 高亚辉, 等. 氮、磷比对两种赤潮藻生长特性的影响及藻间竞争作用[J]. 海洋学报, 2006, 28(5): 117-122. doi: 10.3321/j.issn:0253-4193.2006.05.015 [33] 曲克明, 陈碧鹃, 袁有宪, 等. 氮磷营养盐影响海水浮游硅藻种群组成的初步研究[J]. 应用生态学报, 2000, 27(3): 445-448. doi: 10.3321/j.issn:1001-9332.2000.03.029


计量
- 文章访问数: 3196
- HTML全文浏览量: 158
- PDF下载量: 814
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
