
Analysis of grey relationship between morphological traits and body weight of pearl oyster (Pinctada fucata)
-
摘要:
文章采用灰色关联分析方法对合浦珠母贝(Pinctada fucata)12个家系的壳长、壳高、壳宽和绞合线长4个形态性状与体质量的关联度进行了分析和评价。结果显示,12个家系的平均体质量、壳长、壳高、壳宽和绞合线长分别为35.96 g、57.36 mm、59.24 mm、23.55 mm和56.93 mm。各个性状的变异系数较大(8.30%~21.30%),其中壳高和壳宽的变异系数相对较小,而体质量的变异系数相对较大。体质量与各形态性状灰色关联系数之和以壳高为最大(8.950 8),且家系间较均一,壳长为最小(8.189 8),但家系间变化幅度较大。体质量与4个形态性状的关联度大小趋势为壳高(0.745 9)>绞合线长(0.714 8)>壳宽(0.705 9)>壳长(0.682 5)。根据上述结果,可将壳高作为间接选育合浦珠母贝体质量性状的重要评价指标,以提高体质量的选育效率。
Abstract:We conducted a grey relational analysis to examine the relationship between shell traits and body weight of 12 selective breeding families of Pinctada fucata. The results show that the average body weight, shell length, shell height, shell width and hinge length were 35.96 g, 57.36 mm, 59.24 mm, 23.55 mm and 56.93 mm, respectively. The coefficients of variation in body weight and shell traits were ranked as follows: body weight (21.30%) > shell length (9.91%) > hinge length (9.12%) > shell height (8.85%) > shell width (8.30%). The grey relational degrees between each shell trait and body weight were ranked as follows: shell height (0.745 9) > hinge length (0.714 8) > shell width (0.705 9) > shell length (0.682 5). It is suggested that shell height which had the closest relationship with body weight could be regarded as an important factor for evaluation of P.fucata breeding practice for high growth lines selection and improvement of selection efficiency.
-
随着人类对海洋开发利用的日益增长,海洋渔业自然资源的衰退速度远远大于它的恢复速度,对于海洋渔业资源数量动态的研究日益成为海洋渔业科学研究的热点之一[1-6]。实际种群分析法(virtual population analysis,VPA)[7-8],又称为世代分析法(cohort analysis,CA)[9],是渔业资源评估的经典模型之一。它通过分析渔业历史产量数据(分年龄组产量数据)和其附属数据[捕捞努力量或资源量指数(CPUE)数据]来研究渔业的种群结构(分年龄组的种群数量和捕捞死亡系数),是总允许捕捞产量(total allowable catch,TAC)计算的主要模型之一[10-12]。
由于VPA方法本身具有不确定性,单纯的分年龄组产量数据不能计算最末年(和最高年龄)的捕捞死亡系数(和种群数量),所以,VPA的计算通常需要分年龄组产量数据和附属数据(捕捞努力量或CPUE数据)[11-12]。为此,产生了2类模型,集成统计VPA方法(integrated statistical VPA)和调谐VPA方法(tuning VPA)。前者在北美有较多应用[13-14],它们尽可能准确地描述了种群的动态和允许所有的观测因子包含误差,然而,通常它们需要估计大量的参数和需要大量的计算,并且有时计算结果不稳定[10];后者发源于欧洲[11, 15],虽然缺乏严格的数学基础,它们简单易于理解和不需要繁杂的计算,在实际的渔业资源评估中有着广泛的应用。本文集中探讨了后者。虽然对VPA的调谐方法的研究已经有所报道[16-17],但是它们尚缺乏系统性、完整性,特别是缺乏应用现代蒙特卡罗模拟技术对它们的研究。
本文研究了最具有典型性的3种VPA的调谐方法,JAM(judicious average method)[11],LS(Laurec and Shepherd method)[15],Hybrid[11, 16]。应用系统的蒙特卡罗模拟技术对这3种方法的研究是本文对鱼类种群动力学的贡献。
1. 方法
VPA的概念最早由FRY[7]提出,但只是在GULLAND[8]之后,这个思想才得到广泛认可。GULLAND方法应用了鱼类种群动力学的基础公式:
$$ C_{y, a}=N_{y, a} \frac{F_{y, a}}{Z_{y, a}}\left[1-\exp \left(-Z_{y, a}\right)\right] $$ (1) $$ N_{y+1, a+1}=N_{y, a} \exp \left(-Z_{y, a}\right) $$ (2) 这里y是时间,a是年龄,C是产量,N是种群数量,F是捕捞死亡系数,Z是总死亡系数。
公式(1)/公式(2),则得:
$$ \frac{C_{y, a}}{N_{y+1, a+1}}=\frac{F_{y, a}\left[1-\exp \left(-Z_{y, a}\right)\right]}{Z_{y, a} \exp \left(-Z_{y, a}\right)} $$ (3) 联合公式(2)和(3)则构成了VPA方法的计算公式[1]。
给定最末年(和最高年龄)的捕捞死亡系数(或种群数量),VPA可以迅速和方便地把难以理解的分年龄产量数据转化为易于理解的捕捞死亡系数和世代强度。由于VPA的不确定性,亦即VPA的解不是唯一的,任何一个给定的最末年(和最高年龄)的捕捞死亡系数(或种群数量),可以计算出一组捕捞死亡系数和世代强度,所以,需要另外的信息(通常是捕捞努力量或资源量指数)来确定正确的最末年(和最高年龄)捕捞死亡系数(或种群数量)。在为此产生的2类VPA模型(见前)中,调谐VPA方法以其简约性有着广泛的应用[11, 17]。最典型的调谐VPA方法主要有3种:JAM、LS和Hybrid(混合方法)[11]。
JAM法(明智的平均方法)[11]:当只有分年龄组的产量数据时,对于每一个年龄,假设最末几年(通常是3~5年)的捕捞死亡系数(F)相对稳定,则最末年的F是其前几年F的平均值。同样对于每年,假设最高几个年龄(3~5年龄)的开发率曲线平坦,则最高年龄的F是其前几年龄F的平均值。重复以上计算,通常情况下VPA将很快收敛于一个稳定解。
LS法[15]:当存在捕捞努力量(E)(或资源量指数)时,假设捕捞效率(q,catchability)稳定(q=F/E),类似于JAM方法,可以计算得最末年和最高年龄的q,于是F。
Hybrid法[11, 16]:类似于LS方法,只不过假设捕捞效率有随时间变化的趋势(可能由于捕捞努力量的标准化有误差,或种群的分布和数量有变化,或环境条件有变化),应用回归法可以计算最末年的q。最高年龄的q的计算同LS方法。
2. 数据和模拟分析
模拟的产量和种群数量分别由公式(1)和(2)计算。假设补充恒定,选择性在高年龄趋于平坦。捕捞效率不随时间变化。随着时间变化的捕捞努力量被划分为3种情况,恒定、增加和减少,后两者的变化幅度为每年变化10%。白噪声(white noise)水平设定为0,10%,20%,30%。正态分布的随机变量(白噪声)通过Box-Mueller方法产生,即选择2个单一分布的随机数U1和U2并设定:
$$ Z_1=\sqrt{-2 \log \left(U_1\right)} \cos \left(2 \pi U_2\right) $$ (4) $$ Z_2=\sqrt{-2 \log \left(U_1\right)} \sin \left(2 \pi U_2\right) $$ (5) Z1和Z2便是平均值等于0,方差为1的正态随机变量。
模拟分析重复了300次以获得稳定的解。模拟计算的结果以补充的“偏倚比率”表示,即估计值与真实值比率的以2为底的对数。所以0.0值表示无偏估计,1.0值表示2倍的过高估计,-1.0是2倍的过低估计[18]。我们还计算了平均绝对离差(mean absolute deviation,MAD)[19],亦即计算了补充的偏倚比率的绝对值。本文用箱型图表示了300次重复的MAD。
3. 结果
图 1是在捕捞努力量不随着时间变化的模拟数据中,当白噪声的变异系数为0~30%时,表示300次重复的MAD结果的箱型图。在没有白噪声时(图 1-a),3种方法的估计值都逼近于模拟数据的真值。在白噪声增加时,3种方法的估计值(MAD的中值)仍然接近于真值,且JAM方法的结果比另外2种方法的结果要好。
![]() 图 1 在捕捞死亡系数不随着时间变化的模拟数据中,当白噪声的变异系数为WH=0(a), 0.1(b), 0.2(c), 0.3(d)时,表示300次重复的MAD结果的箱型图MAD是平均绝对离差,即补充的偏倚比率的绝对值; “偏倚比率”是估计值与真实值比率的以2为底的对数。
图 1 在捕捞死亡系数不随着时间变化的模拟数据中,当白噪声的变异系数为WH=0(a), 0.1(b), 0.2(c), 0.3(d)时,表示300次重复的MAD结果的箱型图MAD是平均绝对离差,即补充的偏倚比率的绝对值; “偏倚比率”是估计值与真实值比率的以2为底的对数。
最具有典型性的3种调谐VPA方法是:JAM,LS,HybridFigure 1. MAD boxplots of 300 replicates for the status quo fishing mortality and white noises levels (coefficient of variation) of WH=0(a), 0.1(b), 0.2(c), 0.3(d)MAD is mean absolute deviation, i.e. the absolute bias ratio of recruitments.
Bias ratio is the ratios of estimated to true parameter values on log2 scale.
The three most commonly used VPA tuning methods are: JAM, LS, Hybrid.图 2是在捕捞努力量随时间增加的模拟数据的箱型图。在没有白噪声时(图 2-a),LS和Hybrid方法的估计值都逼近于模拟数据的真值,而JAM方法的估计值偏离真值较远。在白噪声增加时,3种方法的估计值(MAD的中值)不再接近于真值,且LS和Hybrid方法的结果相对比JAM方法的结果要好一些。
![]() 图 2 在捕捞死亡系数随时间增加的模拟数据中,当白噪声的变异系数为WH=0(a),0.1(b),0.2(c),0.3(d)时,表示300次重复的MAD结果的箱型图更多的解释见图 1的标题Figure 2. MAD boxplots of 300 replicates for the fishing mortality which increased with time and white noises levels (coefficient of variation) of WH=0(a), 0.1(b), 0.2(c), 0.3(d)For more see the caption of Fig. 1.
图 2 在捕捞死亡系数随时间增加的模拟数据中,当白噪声的变异系数为WH=0(a),0.1(b),0.2(c),0.3(d)时,表示300次重复的MAD结果的箱型图更多的解释见图 1的标题Figure 2. MAD boxplots of 300 replicates for the fishing mortality which increased with time and white noises levels (coefficient of variation) of WH=0(a), 0.1(b), 0.2(c), 0.3(d)For more see the caption of Fig. 1.图 3是在捕捞努力量随时间减少的模拟数据的箱型图。在没有白噪声时(图 3-a),JAM的估计值逼近于模拟数据的真值,而Hybrid方法和LS方法的估计值偏离真值较远。在白噪声增加时,3种方法的估计值(MAD的中值)均不同程度地偏离了真值,且JAM方法的结果相对比LS和Hybrid方法的结果要好一些。
![]() 图 3 在捕捞死亡系数随时间减少的模拟数据中,当白噪声的变异系数为WH=0(a),0.1(b),0.2(c),0.3(d)时,表示300次重复的MAD结果的箱型图更多的解释见图 1的标题Figure 3. MAD boxplots of 300 replicates for the fishing mortality which decreased with time and white noises levels (coefficient of variation) of WH=0(a), 0.1(b), 0.2(c), 0.3(d)For more see the caption of Fig. 1.
图 3 在捕捞死亡系数随时间减少的模拟数据中,当白噪声的变异系数为WH=0(a),0.1(b),0.2(c),0.3(d)时,表示300次重复的MAD结果的箱型图更多的解释见图 1的标题Figure 3. MAD boxplots of 300 replicates for the fishing mortality which decreased with time and white noises levels (coefficient of variation) of WH=0(a), 0.1(b), 0.2(c), 0.3(d)For more see the caption of Fig. 1.4. 讨论
4.1 VPA调谐方法的选择
对JAM、LS和Hybrid 3种方法的蒙特卡罗模拟研究表明,这3种VPA调谐方法对数据中包含的白噪声都不太敏感,但是对捕捞死亡系数F的变化比较敏感。从我们的研究结果来看,当F随着时间有增加倾向的情况下,LS和Hybrid比较稳定,JAM则对这种趋势尤为敏感(图 2);当F随着时间有减少倾向的情况下,JAM比较稳定,LS和Hybrid则对该趋势尤为敏感(图 3)。因此,我们建议,当采用3种VPA调谐方法对某种资源进行分析评估时,首先应对历年的F值的变化进行研究。当F的变化不明显时,3种方法都是可选的方法,并且受白噪声的影响不大;当F随着时间有增加倾向的情况下,LS和Hybrid方法比较适用;而当F随着时间有减少倾向的情况下,则JAM比较理想。
4.2 对数据的选择
(1) 如果一个渔业种群积累有大量的数据,通常并不是盲目地把它们全部地收入到计算过程中,我们需要有选择地利用它们。特别是最低和最高年龄组的数据通常含有大量的取样误差,对它们的使用需要额外的谨慎。一般来说,取样精度的验前估计是没有的,但是,如果在数据矩阵中出现了大量的零,几乎可以肯定地指示取样存在严重的误差。在这种情况下,最好的解决办法可能就是把这些出现大量零的数据矩阵从计算中删除。
(2) 众所周知,渔业种群是一个处于不断变化之中的动态过程,历史数据不能代表现在的情况。这是因为对于捕捞方式、商业捕捞船队或调查船队等可能存在一些没有记录的变化;或由于气候的变化或种间关系的影响,种群的空间分布可能发生了变化;或数据积累的过程可能发生了变化。一般而言,人们通常更倾向于相信最近的数据。在实际的VPA计算中我们建议使用最近5~10年的数据,20年以上的数据可能太老了[18, 20]。
(3) 有必要在分析中决定是否包括所有船队的数据。在实际中把坏数据和好数据放在一起使用的意义不大,它只会增加准备数据的工作量和增加大量的输出结果,而且没有或有很少益处。现代的调谐VPA技术尚不能拒绝坏数据,而且常常会受到负面的影响,特别是数据中的那些最近年份的异常值。所以要拒绝那些被认知出的低质量的数据,虽然目前尚无明确的鉴别标准来辨别数据的好坏。
4.3 最高年龄的捕捞死亡系数
第二个需要作出的重要决定是如何选择最高年龄的捕捞死亡系数。在VPA计算的早期,通常是由反复试验来确定这些捕捞死亡系数。当捕捞死亡系数的总体水平较高(大约处于1.0)时,由于VPA的收敛特性,它们对其它年龄的捕捞死亡系数的计算结果影响不大。然而,对于中低水平的捕捞死亡系数,错误的最高年龄捕捞死亡系数可能导致无法解释的结果或奇怪的开发模式。
所以我们通常假设渔业种群的开发模式在最高的几个年龄组是平坦的,亦即假设最高年龄的捕捞死亡系数是其相邻的几个(3~5个)低年龄组的捕捞死亡系数的平均值。仍然需要强调的是不能选择低的捕捞死亡系数,因为这可以破坏VPA的收敛特性。如果对于一个渔业种群不能确定其捕捞死亡系数的水平,建议对最高的几个年龄组使用高水平的捕捞死亡系数。
4.4 自然死亡系数
VPA计算的另一个困难是假设已知自然死亡系数,且不随时间变化。虽然这不是一个合理的假设,但是更复杂的模型(例如MSVPA,多鱼种VPA)需要更多的取样(确定种间关系)和更多的计算量[19, 21]。所以,虽然在理论上应该使用MSVPA技术,但是实际的种群评估大多是以单鱼种为基础的。这对于短期预报(一个渔汛或一年)影响不大,但是对于长期预报(5~10年)可能会产生严重的误差。
-
![]()
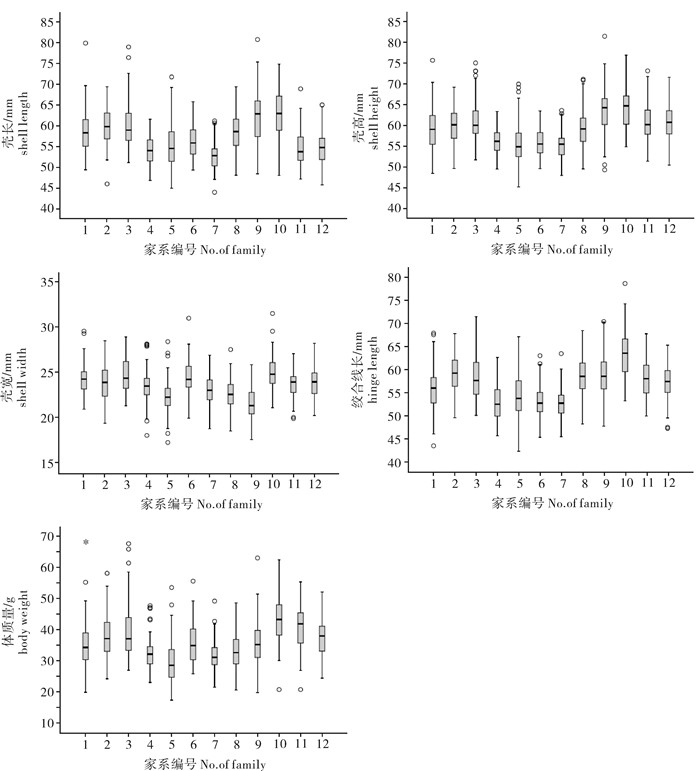
图 1 合浦珠母贝不同家系5个性状的箱线图
盒子由中位数、第一四分位数和第三四分位数组成;温和异常值和极端异常值分别用“○”和“*”表示
Figure 1. Box plots of five traits of different families of P.fucata
The medium, 25th and 75th percentiles are plotted as boxes; mild outliers and extreme outliers are shown as ○ and *, respectively.
表 1 合浦珠母贝不同家系生长性状无量纲化结果
Table 1 Nondimensionalization of growth traits of different P.fucata families
性状trait 变量编号No. of variables 家系编号No.of family 1 2 3 4 5 6 7 8 9 10 11 12 体质量/g body weight X′0 -0.261 4 0.443 2 1.005 9 -0.901 4 -1.628 4 -0.089 6 -1.048 3 -0.704 7 -0.107 1 1.777 8 1.167 8 0.346 1 壳长/mm shell length X′1 0.310 3 0.742 5 0.7721 -1.009 9 -0.702 0 -0.382 3 -1.385 8 0.298 5 1.381 9 1.660 1 -0.873 7 -0.811 6 壳高/mm shell height X′2 -0.002 8 0.239 1 0.580 2 -1.049 5 -1.241 7 -1.109 2 -1.341 0 -0.042 5 1.388 5 1.584 0 0.477 6 0.517 3 壳宽/mm shell width X′3 0.670 5 0.345 0 1.189 4 -0.143 3 -1.221 6 0.741 8 -0.489 1 -0.946 9 -1.964 2 1.362 3 0.223 0 0.233 1 绞合线长/mm hinge length X′4 -0.313 2 0.728 9 0.403 5 -1.214 5 -0.763 9 -1.189 5 -1.311 6 0.553 7 0.547 4 2.059 0 0.397 2 0.103 0  下载: 导出CSV
下载: 导出CSV
表 2 合浦珠母贝不同家系关联系数母序列和子序列的绝对差值
Table 2 Absolute difference between reference array and comparative array of different P.fucata families
绝对差值absolute difference (Δi) 家系编号No.of family 1 2 3 4 5 6 7 8 9 10 11 12 Δ1 0.571 7 0.299 3 0.233 8 0.108 5 0.926 4 0.292 7 0.337 5 1.003 2 1.489 0 0.117 7 2.041 5 1.157 7 Δ2 0.258 6 0.204 1 0.425 7 0.148 1 0.386 7 1.019 6 0.292 7 0.662 2 1.495 6 0.193 8 0.690 2 0.171 2 Δ3 0.931 9 0.098 2 0.183 5 0.758 1 0.406 8 0.831 4 0.559 2 0.242 2 1.857 1 0.415 5 0.944 8 0.113 0 Δ4 0.051 8 0.285 7 0.602 4 0.313 1 0.864 5 1.099 9 0.263 3 1.258 4 0.654 5 0.281 2 0.770 6 0.243 1
下载: 导出CSV
表 3 合浦珠母贝5个性状的描述性统计量
Table 3 Descriptive statistics of five traits of P.fucata
性状trait 平均值mean 最大值maximum 最小值minimum 极差ranges 标准差SD 变异系数/% CV 体质量/g body weight 35.96 68.81 17.27 51.54 7.66 21.30 壳长/mm shell length 57.36 80.84 43.82 37.02 5.69 9.91 壳高/mm shell height 59.24 81.58 45.07 36.51 5.24 8.85 壳宽/mm shell width 23.55 31.50 17.25 14.25 1.96 8.30 绞合线长/mm hinge length 56.93 78.68 42.36 36.32 5.19 9.12
下载: 导出CSV
表 4 合浦珠母贝不同家系体质量与各形态性状的灰色关联系数
Table 4 Grey relational coefficient between body weight and each morphological trait of different P.fucata families
家系编号No. of family 关联系数(ξi) relational coefficient ξ1 ξ2 ξ3 ξ4 1 0.673 5 0.838 4 0.549 3 1.000 0 2 0.812 5 0.875 7 0.958 5 0.821 0 3 0.854 9 0.741 5 0.890 6 0.660 8 4 0.949 8 0.917 6 0.602 9 0.804 1 5 0.550 8 0.762 1 0.751 3 0.568 9 6 0.816 6 0.525 7 0.579 1 0.505 8 7 0.789 7 0.816 6 0.678 9 0.835 3 8 0.529 9 0.637 3 0.849 2 0.470 6 9 0.427 4 0.426 2 0.372 7 0.640 2 10 0.942 1 0.883 1 0.746 8 0.823 8 11 0.350 2 0.626 9 0.545 7 0.598 7 12 0.492 3 0.899 8 0.946 0 0.848 6 合计total 8.189 7 8.950 9 8.471 0 8.577 8
下载: 导出CSV
表 5 合浦珠母贝不同家系各形态性状对体质量的灰色关联度
Table 5 Grey relational degree of each shell trait of different P.fucata families
性状trait 关联度relational degree 关联序relational order 壳长/mm shell length 0.682 5 4 壳高/mm shell height 0.745 9 1 壳宽/mm shell width 0.705 9 3 绞合线长/mm hinge length 0.714 8 2
下载: 导出CSV
-
[1] 郭华阳, 陈明强, 王雨, 等. 黄边糙鸟蛤野生群体主要经济性状间的相关性及通径分析[J]. 南方水产科学, 2013, 9(2): 1-8. doi: 10.3969/j.issn.2095-0780.2013.02.001 [2] 刘伟, 苏胜彦, 董在杰, 等. 3个鲤群体的微卫星标记与生长性状相关性分析[J]. 南方水产科学, 2012, 8(3): 17-24. doi: 10.3969/j.issn.2095-0780.2012.03.003 [3] 肖述, 喻子牛, 符政君. 香港巨牡蛎雌雄群体的数量性状通径分析[J]. 南方水产科学, 2011, 7(4): 1-9. doi: 10.3969/j.issn.2095-0780.2011.04.001 [4] 刘永新, 刘英杰, 周勤, 等. 牙鲆主要生长性状与体质量的灰色关联度分析[J]. 中国水产科学, 2014, 21(2): 205-213. doi: 10.3724/SP.J.1118.2014.00205 [5] 王玉涛, 郭丽君, 邓勇, 等. 利用皮特兰作父系商品猪肉品质的灰色关联度分析[J]. 上海交通大学学报: 农业科学版, 2008, 26(6): 524-527. doi: 10.3969/j.issn.1671-9964.2008.06.005 [6] 李世凯, 江敏, 潘若雷, 等. 罗氏沼虾养殖塘叶绿素a与水质因子的灰色关联分析[J]. 生态与农村环境学报, 2014, 30(2): 257-261. doi: 10.3969/j.issn.1673-4831.2014.02.019 [7] 权月伟, 米换房, 翟雷霞, 等. 棉花品种主要数量性状与皮棉产量的灰色关联分析[J]. 棉花科学, 2013, 35(4): 33-36. doi: 10.3969/j.issn.2095-3143.2013.04.006 [8] 马文清, 郭强, 邱文武, 等. 利用灰色关联分析法评价甘蔗自育新品系[J]. 广东农业科学, 2014, 41(5): 71-78. doi: 10.3969/j.issn.1004-874X.2014.05.017 [9] 吕耀平. 影响越冬罗非鱼死亡率环境因子的灰色关联分析[J]. 水产科学, 2005, 24(8): 14-16. doi: 10.3969/j.issn.1003-1111.2005.08.005 [10] 苏胜彦, 董在杰, 曲疆奇, 等. 3个鲤群体杂交后代生长性状的灰色关联及复合杂交后代的体重预测分析[J]. 水产学报, 2011, 35(1): 20-26. doi: 10.3724/SP.J.1231.2011.17109 [11] 龙敏明, 黄桂菊, 邹记兴, 等. 育珠对合浦珠母贝N19和Prismalin-14基因表达水平的影响[J]. 南方水产科学, 2013, 9(5): 58-63. doi: 10.3969/j.issn.2095-0780.2013.05.010 [12] 罗会, 刘宝锁, 黎火金, 等. 合浦珠母贝不同地理群体的形态差异和判别分析[J]. 广东农业科学, 2013, 40 (12): 171-174. doi: 10.3969/j.issn.1004-874X.2013.12.052 [13] TAKASHI A, TAKASHI I, NARIAKI I, et al. Post-operative care of implanted pearl oysters Pinctada fucata in low salinity seawater improves the quality of pearls[J]. Aquaculture, 2014, 422/423: 232-238. doi: 10.1016/j.aquaculture.2013.12.022
[14] OKAMOTO C, KOMARU A, HAYASHI M, et al. Variation of shell-closing strength among several families in pearl oyster, Pinctada fucata martensii[J]. Aquac Sci, 2006, 54(4): 525-529. doi: 10.11233/aquaculturesci1953.54.525
[15] ISHIKAWA T, OKAMOTO C, HAYASHI M, et al. The heritability of the shell-closing strength in the Japanese pearl oyster, Pinctada fucata martensii[J]. Aquac Sci, 2009, 57(1): 77-82. doi: 10.11233/aquaculturesci.57.77
[16] 黎火金, 刘宝锁, 罗会, 等. 合浦珠母贝幼贝生长性状的遗传参数估计[J]. 中国水产科学, 2013, 20(6): 1-6. doi: 10.3724/SP.J.1118.2013.01182 [17] HE M X, GUAN Y Y, YUAN T, et al. Realized heritability and response to selection for shell height in the pearl oyster Pinctada fucata (Gould)[J]. Aquac Res, 2008, 39(8): 801-805. doi: 10.1111/j.1365-2109.2008.01889.x
[18] DENG YW, FU S, DU X D, et al. Realized heritability and genetic gain estimates of larval shell length in the Chinese pearl oyster Pinctada martensii at three different salinities[J]. N Am J Aquac, 2009, 71(4): 302-306. doi: 10.1577/A08-024.1
[19] YU S, HE M X. Differential gene expression identified by RNA-Seq and qPCR in two sizes of pearl oyster (Pinctada fucata)[J]. Gene, 2014, 538(2): 313-322. doi: 10.1016/j.gene.2014.01.031
[20] LIU W G, HUANG X D, LIN J S, et al. Effect of temperature on gene expression in the pearl oyster Pinctada fucata[J]. J Ocean Univ China, 2014, 13(3): 509-515. doi: 10.1007/s11802-014-2147-y
[21] 喻达辉, 王小玉, 黄桂菊, 等. 合浦珠母贝遗传连锁图谱的构建[J]. 中国水产科学, 2007, 14(3): 361-368. https://www.nstl.gov.cn/paper_detail.html?id=f3613d6f72cb32b3e95aeffe0a13e956 [22] 何毛贤, 史兼华, 林岳光, 等. 马氏珠母贝生长性状的相关分析[J]. 海洋科学, 2006, 30(11): 1-4. https://cstj.cqvip.com/Qikan/Article/Detail?id=23295382&from=Qikan_Article_Detail [23] DENG Y W, DU X D, WANG Q H, et al. Correlation and path analysis for growth traits in F1 population of pearl oyster Pinctada martensii[J]. Mar Sci Bull, 2008, 10(2): 68-73. https://www.researchgate.net/publication/267423852_Correlation_and_Path_Analysis_for_Growth_Traits_in_F_1_Population_of_Pearl_Oyster_Pinctada_martensii
[24] 陈飞飞, 黄桂菊, 陈明强, 等. 合浦珠母贝三亚养殖群体生长性状的相关与通径分析[J]. 广东农业科学, 2012, 39(9): 122-125. doi: 10.3969/j.issn.1004-874X.2012.09.040 [25] 谷龙春, 李金碧, 喻达辉, 等. 合浦珠母贝双列杂交家系的建立与遗传分析[J]. 水产学报, 2010, 34(1): 26-31. doi: 10.3724/SP.J.1231.2010.06498 [26] 李建立, 王春德, 李朝霞, 等. 紫扇贝和海湾扇贝杂交家系的生长和通径分析[J]. 海洋科学, 2012(8): 15-20. https://www.nstl.gov.cn/paper_detail.html?id=13d0b46ad914629fc344d8e145bcf16c [27] 孙泽伟, 郑怀平, 杨彦鸿, 等. 近江牡蛎养殖群体数量性状间的相关及通径分析[J]. 中国农业通报, 2010, 26(6): 332-336. doi: 10.11924/j.issn.1000-6850.2009-2269 [28] 刘广文, 林坚士, 何毛贤. 不同贝龄华贵栉孔扇贝数量性状的通径分析[J]. 南方水产科学, 2012, 8(1): 43-48. doi: 10.3969/j.issn.2095-0780.2012.01.007 [29] 严福升, 王志刚, 刘旭东, 等. 3月龄牙鲆形态性状对体质量的通径分析[J]. 渔业科学进展, 2010, 31(2): 45-50. http://journal.yykxjz.cn/yykxjz/ch/reader/view_abstract.aspx?file_no=20100207&flag=1 [30] 王凯, 刘海金, 刘永新, 等. 牙鲆形态性状对体重的影响效果分析[J]. 上海水产大学学报, 2008, 17(6): 655-660. http://shhydxxb.ijournals.cn/shhy/article/abstract/200806161



计量
- 文章访问数: 3632
- HTML全文浏览量: 192
- PDF下载量: 1640
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
