
Analysis on stomach content of bigeye tuna (Thunnus obesus) in the eastern-central Pacific Ocean
-
摘要:
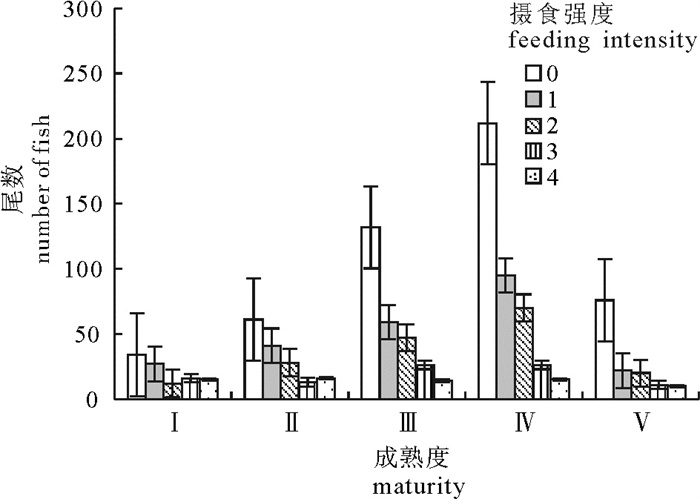
大眼金枪鱼(Thunnus obesus)是金枪鱼延绳钓的主要捕捞对象,分析其胃含物组成有助于更好地认识其摄食特征。根据2010年9月至2011年1月在太平洋中东部海域延绳钓渔业中采集的大眼金枪鱼胃含物样品,采样传统胃含物分析法对大眼金枪鱼摄食种类组成、多样性和摄食强度进行研究。结果表明,大眼金枪鱼的胃含物主要包括鱼类、头足类和甲壳类,其中优势饵料生物隶属于柔鱼科、褶胸鱼科和蛇鲭科。摄食强度组成随叉长变化而显著变化(单因素方差分析,P < 0.05)。空胃的个体所占比例很高,且随性腺成熟度增加而增大,成熟度Ⅰ期至Ⅴ期的空胃率依次为32.7%、38.4%、47.5%、50.7%和50.7%。摄食强度组成不存在显著的性别间差异(K-S检验,P>0.05)。各月份摄食强度组成差异显著(单因素方差分析,P < 0.05),10月平均摄食强度最高,1月最低。
Abstract:Bigeye tuna (Thunnus obesus, BET) is one of the main target species by tuna longline. Analysis on stomach content of BET can improve understanding its feeding characteristics. Based on BET stomach samples collected from longline fishery in the eastern-central Pacific Ocean from September 2010 to January 2011, we analyzed the prey composition, diversity and feeding intensity (i.e., stomach fullness) of BET. The results show that fishes, cephalopods and crustacean were main preys of BET, with dominant prey species groups of Ommastrephidae, Sternoptychidae and Gempylidae. The feeding intensity compositions of BET were significantly different among various fork-length samples as indicated by One-Way ANOVA (P < 0.05). The proportion of empty stomachs in BET samples which was high, increasing along with gonad maturity stages (empty stomach percentages were 32.7%, 38.4%, 47.5%, 50.7% and 50.7% for maturity stages I~V, respectively). There was no significant difference in feeding intensity composition between males and females (K-S test, P>0.05). Moreover, the feeding intensity composition differed monthly (One-Way ANOVA, P < 0.05), with the highest stomach fullness in October and the lowest in January.
-
中越北部湾划界后,渔场面积缩小,导致近海捕捞压力加大,部分渔船转向中、西沙渔场作业寻求出路。中、西沙群岛海域位于中国南海中北部,渔业资源丰富,是中国的传统优良渔场①。在该海域作业的渔船主要来自南海3省、区,作业方式以灯光围网和灯光罩网为主,另外有少量钓具、潜捕和刺网作业。受海况和南海北部休渔的影响,每年的作业时间集中在2~5月。中国长期以来一直关注中、西沙渔场的渔业资源状况,多次派出监测船进行调查[1-7],然而未对在该海域作业渔船的捕捞能力进行过研究。不了解当地的渔业生产状况就无法有效地实施渔业管理,长此以往必然威胁到渔业资源的可持续利用和岛礁生物多样性的保护。文章根据2008年渔港调查数据,分析了在中、西沙群岛海域作业的灯光围网船和灯光罩网船的捕捞能力,以期为该渔业的有效管理和中、西沙群岛海域中上层渔业资源的合理开发利用提供参考。
① 西、南、中沙渔业资源调查组. 西、中沙、南沙北部海域大洋性鱼类资源调查报告[R]. 广州: 国家水产总局南海水产研究所, 1979.
1. 材料与方法
1.1 数据来源
2008年2~10月期间在广东闸坡、广西北海和海南临高、三亚等渔港,对在中、西沙海域灯光作业渔船的生产状况进行了详细的调查,共收集和整理出2008年2~5月该海域作业的26艘灯光围网船的715份渔捞记录和16艘灯光罩网船的771份渔捞记录。渔捞记录包含了主机功率、网具规格、作业时间、作业网次、作业位置、渔获种类和渔获产量等信息。
1.2 方法
分别计算出灯光围网和灯光罩网各渔区及渔获种类的单位捕捞努力量渔获量(catch per unit effort,CPUE),计算了2种CPUE:单船网次产量CPUE1和单船单位功率网次产量CPUE2。
$$ \mathrm{CPUE}_1=\frac{\text { catch }}{\text { sets }} $$ (1) $$ \mathrm{CPUE}_2=\frac{\text { catch }}{\text { sets } \times \text { power }} $$ (2) 式中catch表示单船渔获量(kg);sets表示投网次数;power表示主机功率数(kW)。
采用广义线性模型(generalized linear model,GLM)对灯光围网船和灯光罩网船的产量和努力量数据进行了方差分析,分析影响生产效益的主要原因。广义线性模型如下:
$$ C=\mathtt{μ}+P+S+L+G+A+\mathtt{ε} $$ (3) 式中因变量为单船春季总产量C,μ为全局平均值,自变量有主机功率P、投网次数S、灯光功率L、网具规格G,作业渔区数A,残差ε~N(0,σ2)。自变量均为分类变量,按数值大小分成了不同的等级(表 1和表 2)。
表 1 灯光围网自变量的分类Table 1. Independent variable of light attracting purse seiners主机功率/kW
main engine
power等级
level投网次数
net setting等级
level网衣面积/104 m2
netting areas等级
level灯光功率
/104 kW
light power等级
level渔区数
number of
fishing zones等级
level≤200 1 ≤15 1 ≤10 1 ≤15 1 1 1 201~300 2 16~30 2 10~15 2 >15 2 2 2 301~400 3 31~45 3 15~20 3 - - 3 3 ≥401 4 >45 4 >20 4 - - - - 表 2 灯光罩网自变量的分类Table 2. Independent variable of light attracting falling gears主机功率/kW
main engine
power等级
level投网次数
net setting等级
level网衣面积/104 m2
netting areas等级
level灯光功率
/104 kW
light power等级
level渔区数
number of
fishing zones等级
level≤200 1 ≤200 1 ≤150 1 ≤200 1 ≤5 1 201~250 2 201~300 2 151~200 2 201~250 2 6~10 2 251~300 3 301~400 3 >200 3 >250 3 >10 3 ≥300 4 >400 4 - - - - - - 利用随机生产边界法[8] (stochastic production frontiers,SPF)对灯光围网船和灯光罩网船的捕捞效率进行了评价。假定有劳动力、资本2个投入因子,且生产函数满足Cobb-Douglas形式,即:
$$ Y_t=A L_t^a K_t^b T_t $$ (4) 式中Yt为时间t内的总产出,Lt为时间t内的劳动力的量,Kt为时间t内的资本量,而Tt为一种技术趋势。实际应用中,在分析截面数据(cross-section data)时,取(4)式的对数形式。
在计算Cobb-Douglas生产边界时,采用如下方程:
$$ \ln Y=\beta_0+\beta_1 \ln (L)+\beta_2 \ln (K)+e $$ (5) 式中,Y、K和L分别为产出、资本和劳动力,e为偏差。令X0=1nY, X1=1nL, X2=1nK,则(5)式可以转化为线性形式:
$$ X_0=\beta_0+\beta_1 X_1+\beta_2 X_2+e $$ (6) 再令X = (1, X1, X2), C = (β0, β1, β2)T,则(6)式可写成
$$ X_0=X C+e $$ (7) 现在的问题归结为如何根据样本点计算参数矢量C的估计值Ĉ,使得产出的估计值大于实际值,也即偏差项e≥0,即e=XĈ-X0≥0 (8)
同时还应保证偏差的平方和e′e为最小。使前沿面与样本点最接近。因为(8)式中e≥0,所以不必求偏差的平方和最小,而只求偏差和最小即可,故问题可简化为一个二次线性规划:
$$ \begin{aligned} & \min Z=L e, \text { s t } \quad X \hat{C} \geqslant X_0 \\ & \hat{C} \geqslant 0 \end{aligned} $$ (9) 式中L= (1, 1, …, 1)。将e=XĈ-X0代入(9)式,则目标函数为
$$ Z=L\left(X \hat{C}-X_0\right) $$ (10) 据此,解这个线性规划问题,就可以得到参数估计值Ĉ,于是得到一个前沿生产函数。实际中,β0的估计值可能为负,为了保证线性规划问题有可行解,可令X= (-1, X1, X2),此时计算出的β0值就为正,满足线性规划约束条件Ĉ≥0。各个投入单元的技术效率可以根据实际值和估计值的比值确定。
在运用SPF对捕捞能力进行评估时,以主机功率×灯光功率为固定投入要素(资本因子),以投网次数为可变投入要素(劳动力因子),应用Cobb-Douglas生产函数计算了每艘渔船的技术效率。技术效率的计算采用了COELI[9]编写的Frontier 4.1运算软件。
2. 结果
2.1 CPUE计算结果
被调查的26艘灯光围网船的单船产量在25.03~229.73 t之间,平均100.77 t。CPUE1在1.00~12.52 t · 网-1,平均4.72 t · 网-1。其中海南三亚籍渔船的平均CPUE1为4.41 t · 网-1,海南临高籍渔船的平均CPUE1为3.39 t · 网-1,广东阳西籍渔船的平均CPUE1为10.24 t · 网-1。CPUE2为3.38~37.84 kg · (网· kW)-1,平均16.96 kg · (网· kW)-1。其中海南三亚籍渔船的平均CPUE2为10.12 kg · (网· kW)-1,海南临高籍渔船的平均CPUE2为16.81 kg · (网· kW)-1,广东阳西籍渔船的平均CPUE2为29.50 kg · (网· kW)-1。
被调查的16艘灯光罩网船的单船产量在5.93~130.03 t之间,平均60.05 t。CPUE1在49.40~451.49 kg · 网-1,平均179.87 kg · 网-1。其中广西籍渔船的平均CPUE1为183.25 kg · 网-1,海南籍渔船的平均CPUE1为169.74 kg · 网-1。CPUE2在0.22~2.05 kg · (网· kW)-1,平均0.77 kg · (网·kW)-1。其中广西籍渔船的平均CPUE2为0.83 kg· (网· kW)-1,海南籍渔船的平均CPUE2为0.62 kg· (网· kW)-1。
2.2 渔获组成
统计2种作业方式的渔获物发现,2008年春季灯光围网渔获物中产量超过200 t的有颌圆鲹(Decapterus lajang)、蓝圆鲹(Decapterus maruadsi)、扁舵鲣(Auxis thazard)、羽鳃鲐(Rastrelliger kanagurta)、小鲔(Euthynnus alleteratus)和大目金枪鱼(Thunnus obesus),分别占总渔获重量的44.89%、22.46%、11.29%、6.83%、6.55%和2.95% (图 1)。灯光罩网主要渔获种类为柔鱼(Ommastrephes spp.)、蓝圆鲹和颌圆鲹,分别占总渔获重量的85.73%、4.43%和4.43% (图 2)。
2.3 方差分析
GLM对灯光围网作业的总方差解释率达到84.2%。影响产量的最重要因素是投网次数(P < 0.01),这符合一般常识,有效投网次数越多,产量越高。其次是作业范围(P < 0.05),作业范围越大,寻找好渔场的概率越高,也表明其生产积极高。其他如主机功率、灯光功率和网具尺度对灯光围网作业的影响较小(P>0.05)(表 3)。
表 3 灯光围网作业的方差分析表Table 3. Variance analysis of light attracting purse seiners误差来源
sourceⅢ类残差平方和
type Ⅲ sum of squares自由度
degree of freedom均方差
mean squareF检验
F test显著性
significanceμ 53 294.22 1 53 294.22 56.13 0.00 主机功率 power 6 083.19 3 2 027.73 2.14 0.15 投网次数 sets 20 873.44 3 6 957.82 7.33 0.00 灯光功率 light power 141.93 1 141.93 0.15 0.71 作业渔区数 area 16 692.29 3 5 564.10 5.86 0.01 网衣面积 netting size 57.56 3 19.19 0.02 0.99 误差 errors 11 392.89 12 949.41 - - GLM对灯光罩网作业的总方差解释率达到98.9%。影响产量的最重要因素亦是投网次数(P < 0.01),有效投网次数越多,产量越高。其次是网衣长度(P < 0.05),网衣长度越大,覆盖的面积越大,渔获的容量也越大。其他如主机功率、灯光功率和作业渔区数对灯光罩网作业的影响较小(P>0.05)(表 4)。
表 4 灯光罩网作业的方差分析表Table 4. Variance analysis of light attracting falling gears误差来源
sourceⅢ类残差平方和
type Ⅲ sum of squares自由度
degree of freedom均方差
mean squareF检验
F test显著性
significanceμ 6 163.77 1 6 163.77 106.19 0.00 主机功率 power 262.02 3 87.34 1.50 0.37 投网次数 sets 6 166.99 3 2 055.66 35.42 0.01 灯光功率 light power 264.26 2 132.13 2.28 0.25 网衣长度 net length 1 545.81 2 772.90 13.32 0.03 作业渔区数 area 301.91 2 150.96 2.60 0.22 误差 errors 174.13 3 58.04 - - 2.4 技术效率分析
Frontier 4.1软件计算结果显示灯光围网船作业的技术效率在34.7%~91.7%,平均69.4%,其中海南三亚籍渔船66.3%,海南临高籍渔船68.6%,广东阳西籍渔船78.0%。灯光围网船的技术效率与主机功率、灯光功率和作业次数等无直接的关系,而与CPUE2有关(R=0.64)(图 3)。灯光罩网船的技术效率在50.4%~92.1%,平均78.2%。其中广西籍渔船平均76.6%,海南籍渔船平均82.9%,技术效率与CPUE2的相关系数0.63(图 4)。
![]() 图 3 灯光围网船的单船产量与技术效率Figure 3. Single vessel yield and technique efficiency of light attracting purse seiners
图 3 灯光围网船的单船产量与技术效率Figure 3. Single vessel yield and technique efficiency of light attracting purse seiners![]() 图 4 灯光罩网船的单船产量与技术效率Figure 4. Single vessel yield and technique efficiency of light attracting falling gears
图 4 灯光罩网船的单船产量与技术效率Figure 4. Single vessel yield and technique efficiency of light attracting falling gears3. 讨论
3.1 SPF评价捕捞能力的优缺点
SPF是目前国际上比较推崇的一种新兴的计算捕捞能力的方法,在国内只见冯春雷等[8]用该方法评价了浙江省的海洋捕捞效率,不似数据包络分析法(data envelopment analysis, DEA)被广泛地使用[10]。SPF用来评价单艘渔船的捕捞能力在国内还是首次,主要面临投入单位的效率是否完全发挥,以及可变投入对产出的贡献度等问题[8]。该方法输出的结果简单,无法像DEA那样对多个投入和产出要素进行规划,不得不借助于方差分析手段对投入要素的贡献度进行解释。由于SPF是基于生产函数的统计分析方法,将投入划分为资本投入和劳动力投入,如文章中将渔船的主机和灯源等作为资本投入,将投网次数作为劳动力投入。SPF分析问题直观实用,参数设置少,因此不失为值得推荐使用的方法。
3.2 2种作业技术效率出现差异的原因
中、西沙作业的灯光围网网周长度范围480~850 m,网衣拉直长度180~240 m;而灯光罩网周长范围145~245 m。比较而言,围网作业面积和深度都比罩网大。灯光围网每晚作业2~3次,平均灯光诱鱼时间3~4 h,每次起网时间约1 h;灯光罩网每晚作业10~16次,除第一网次诱鱼时间较长外,其余灯光诱鱼时间都少于30 min,每次起网时间约20 min。
由于2种作业过程存在差别,以致渔获组成有明显的不同。由图 3和图 4可知,灯光围网瞄准捕捞的是游泳速度比灯光罩网捕捞的柔鱼快得多的中上层鱼类,灯光围网作业技术要求高,下网动作快,起网时间长,不像灯光罩网那样操作简单,总体上技术效率比灯光罩网低。因此,要提高灯光围网的技术效率,就要提高瞄准捕捞技术,增加网次产量,并且利用探鱼仪等助渔设备,寻找到适宜的渔场。
3.3 中、西沙渔业资源的合理开发利用
中、西沙群岛海域蕴藏丰富的渔业资源,尤其是中上层渔业资源具有较大的开发潜力。但作业地点距离大陆远,无法像近海渔业那样方便地实施监控。除了中国渔民在此短期作业外,该海域还有大量越南渔民作业。因此,很有必要定期开展调查和监测,评估作业渔船的捕捞能力,对进入该海域的渔船进行控制,科学规划好总生产容量,为中、西沙渔业资源的可持续利用和生物多样性保护提供保障。
致谢: 感谢金盛2号总指挥乐元伦、平太荣29号船长毛小久和船员等人在海上调查期间给予本研究的帮助, 谨致谢忱! -
![]()
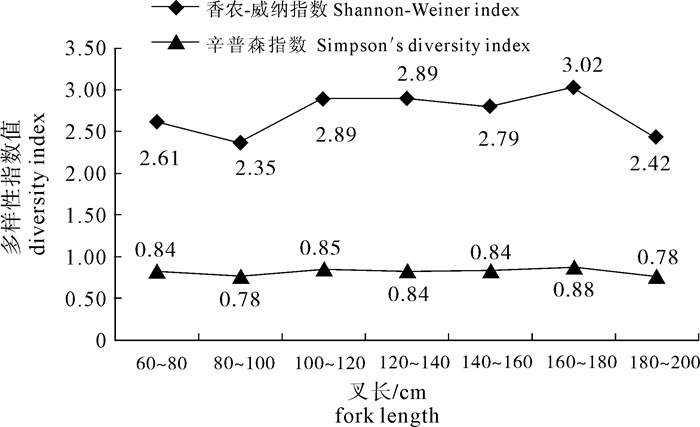
图 2 不同叉长组的香农-威纳指数值和辛普森指数值
Figure 2. Simpson′s diversity index and Shannon-Weiner index for diet composition in different fork-length groups
![]()
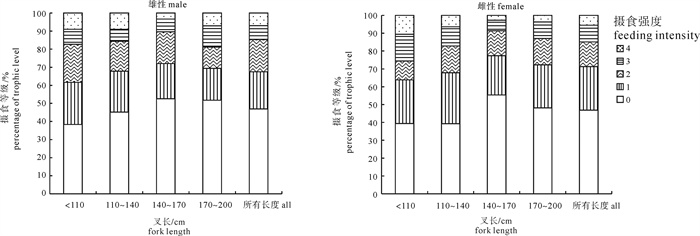
图 3 不同叉长组的摄食强度组成百分比
Figure 3. Percentage of feeding intensity for bigeye tuna with different fork lengths
![]()
图 4 摄食强度分布随性腺成熟度的变化
Figure 4. Change of feeding intensity composition along with gonad maturity stage for bigeye tuna
![]()
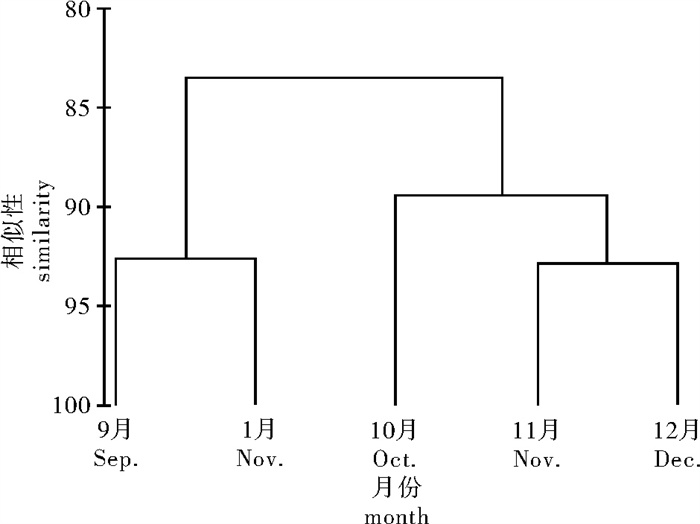
图 5 基于摄食强度的欧式距离构建的大眼金枪鱼聚类关系图
Figure 5. Clustering tree of Euclidean distant of bigeye tuna based on feeding intensity
表 1 大眼金枪鱼的胃含物组成
Table 1 Stomach content composition of bigeye tuna
类别group 饵料生物prey species 个数百分比percentage 出现频次frequency 鱼类fishes 帆蜥鱼Alepisaurus ferox 0.19 0.28 竹筴鱼Trachurus japonicus 2.48 3.69 秋刀鱼Cololnbis snira 2.48 3.69 蛇鲭Gemplus serpens 12.95 19.32 宽尾鳞鲀Abalistes stellatus 0.38 0.57 虱目鱼Chanos chanos 4.38 6.53 白腹鲭Scomber japonicus 5.14 5.97 低褶胸鱼Sternoptyx diaphana 19.81 29.55 头足类Cephalopods 柔鱼科Ommastrephidae 24.00 35.80 乌贼科Sepiidae 0.19 0.28 甲壳类Crustacean 刚毛对虾Peneus setiferus 13.14 19.60 海蜘蛛Pycnogonida 0.19 0.28 未辨别unidentified - 14.48 21.59  下载: 导出CSV
下载: 导出CSV
表 2 太平洋大眼金枪鱼的胃含物组成研究比较
Table 2 Comparison of different studies on stomach content of bigeye tuna in the Pacific Ocean
% 海域sea area 鱼种species 硬骨鱼类
teleost头足类
Cephalopod甲壳类
Crustacean其他
other东太平洋[16] the eastern Pacific Ocean 47 8 45 - 巴布亚新几内亚[22] Papua New Guinea 60 21 15 4 法属波利尼西亚[23] French Polynesia 75 22 1 1 法属波利尼西亚[22] French Polynesia 58 27 11 4 新喀里多尼亚[22] New Caledonia 80 14 1 5 基里巴斯(该研究) Kiribati (this paper) 47 24 14 15
下载: 导出CSV
-
[1] SABATIÉ R, MARSAC F, HALLIER J P, et al. Feeding partitioning among tuna taken in surface and mid-water layers: the case of yellowfin (Thunnus albacares) and bigeye (T. obesus) in the western tropical Indian Ocean[J]. 2004, 3(1): 51-62. https://www.documentation.ird.fr/hor/fdi:010042881
[2] MÉNARD F, LABRUNE C, SHIN Y J, et al. Opportunistic predation in tuna: a size-based approach[J]. Mar Ecol Prog Ser, 2006, 27(3): 223-231. https://www.documentation.ird.fr/hor/PAR00001262
[3] 宋利明, 许柳雄, 陈新军. 大西洋中部大眼金枪鱼垂直分布与温度、盐度的关系[J]. 中国水产科学, 2004, 11(6): 561-566. doi: 10.3321/j.issn:1005-8737.2004.06.013 [4] 朱国平, 许柳雄, 周应祺, 等. 印度洋中西部和大西洋西部水域大眼金枪鱼的食性比较[J]. 生态学报, 2007, 27(1): 135-141. doi: 10.3321/j.issn:1000-0933.2007.01.016 [5] 张玉生, 杨清良, 陈瑞祥, 等. GB/T 12763.6-2007, 海洋调查规范[S]. 北京: 中国标准出版社, 2007: 38-44. [6] 成庆泰, 郑葆珊. 中国鱼类系统检索[M]. 北京: 科学出版社, 1987: 20-30. [7] POTIER M, MARSAC F, CHEREL Y, et al. Forage fauna in the diet of three large pelagic fishes (lancetfish, swordfish and yellowfin tuna) in the western equatorial Indian Ocean[J]. Fish Res, 2007, 83(1): 60-72. doi: 10.1016/j.fishres.2006.08.020
[8] BUCKLEY T W, MILLER B S. Feeding habits of yellowfin tuna associated with fish aggregation devices in American Samoa[J]. Bull Mar Sci, 1994, 55(2/3): 2-3.
[9] MÉNARD F, STÉQUERT B, RUBIN A, et al. Food consumption of tuna in the equatorial Atlantic ocean: FAD-associated versus unassociated schools[J]. Aquat Living Resour, 2000, 13(4): 233-240. doi: 10.1016/S0990-7440(00)01066-4
[10] 杨德康, 孟庆闻. 印度洋西北海域的鱼类[M]. 上海: 上海人民美术出版社, 2000: 10-23. [11] 杨德康, 孟庆闻. 中东大西洋底层鱼类[M]. 上海: 上海人民美术出版社, 2000: 12-20. [12] 戴小杰, 许柳雄. 世界金枪鱼渔业渔获物物种原色图鉴[M]. 北京: 海洋出版社, 2007: 1-124. [13] 牛翠娟, 娄安如, 孙儒泳, 等. 基础生态学[M]. 2版. 北京: 高等教育出版社, 2007: 160-161. [14] RAWLINS M, OXENFORD H A, FANNING P. Preliminary investigation of the diets of large oceanic pelagic species of importance to the longline fishery in Barbados[J]. MSc Research Paper, University of the West Indies, Barbados, 2007, 58: 244-249. https://core.ac.uk/outputs/19540473/
[15] 秦海明. 长江口盐沼潮沟大型浮游动物群落生态学研究[D]. 上海: 复旦大学, 2011. 10.7666/d.y2069227 [16] ALVERSON F G. The food of yellowfin and skipjack tunas in the eastern tropical Pacific Ocean[J]. Inter-Am Trop Tuna Comm Bull, 1963, 7(5): 293-396.
[17] 许柳雄, 朱国平, 宋利明. 印度洋中西部水域大眼金枪鱼的食性[J]. 水产学报, 2008, 32(3): 387-394. doi: 10.3321/j.issn:1000-0615.2008.03.009 [18] BARD F X, YEN S, STEIN A. Habitat of deep swimming tuna (Thunnus obsesus, T. albacares, T. alalunga) Ⅲ Central South Pacific[J]. ICCAT Collect Vol Sci Pap, 1999, 49 (3): 309-317.
[19] SATO K, YOKAWA K, SAITO H, et al. Preliminary stomach contents analysis of pelagic fish collected by Shoyo-Maru 2002 research cruise in the Atlantic Ocean[J]. ICCAT Collect Vol Sci Pap, 2004, 56(3): 1096-1114. https://www.researchgate.net/publication/285471906_Preliminary_stomach_contents_analysis_of_pelagic_fish_collected_by_Shoyo-Maru_2002_research_cruise_in_the_Atlantic_Ocean
[20] PERRIN W F, WARNER R R, FISCUS C H, et al. Stomach contents of porpoise, Stenella spp., and yellowfin tuna, Thunnus albacares, in mixed-species aggregations[J]. Fish Bull, 1973, 71(4): 1077-1091. https://spo.nmfs.noaa.gov/content/stomach-contents-porpoise-stenella-spp-and-yellowfin-tuna-thunnus-albacares-mixed-species
[21] BAKHOUM S A. Diet overlap of immigrant narrow-barred Spanish mackerel Scomberomorus commerson (Lac., 1802) and the largehead hairtail ribbonfish Trichiurus lepturus (L., 1758) in the Egyptian Mediterranean coast[J]. Anim Biodivers Conserv, 2007, 30(2): 147-160. https://www.oalib.com/research/2537106
[22] ALLAIN V, SPC N. What do tuna eat? A tuna diet study[J]. SPC Fish Newslett, 2005, 112: 20-22.
[23] BERTRAND A, BARD F X, JOSSE E. Tuna food habits related to the micronekton distribution in French Polynesia[J]. Mar Biol, 2002, 140(5): 1023-1037. doi: 10.1007/s00227-001-0776-3




计量
- 文章访问数: 3638
- HTML全文浏览量: 158
- PDF下载量: 1722
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
