
Characteristics of phytoplankton community in mesh enclosure isolated ponds with polyculture of Litopenaeus vannamei and Mugil cephalus
-
摘要:
在池塘中构建围网研究了凡纳滨对虾(Litopenaeus vannamei)单养及对虾-鲻(Mugil cephalus)混养池塘中浮游植物的动态变化特征。围网内鲻放养量依次为0(M0)、250(M 250)、500(M500)和800(M800)尾。结果显示,池塘浮游植物共计7门39属65种,其中绿藻门最多(21属33种),蓝藻门次之(7属17种),细胞丰度随鲻放养密度的增大而升高,养殖池塘藻类以小球藻为主(50%以上)。养殖末期鲻放养组绿藻丰度普遍高于对照组,蓝藻丰度反之。M250组绿藻丰度最高而蓝藻最低,且与对照组呈显著差异(P<0.05),其多样性指数在养殖末期居各组最高。养殖前中期高密度鲻组(M500、M800)浮游植物藻相较优,后期则以低密度鲻组M250最优。研究表明,虾塘围网混养鲻可有效提高池塘中绿藻密度,降低蓝藻密度,增加浮游植物群落多样性。
Abstract:We studied the characteristics of phytoplankton community in mesh enclosure isolated pond with polyculture of Litopenaeus vannamei and Mugil cephalus. Shrimp ponds were stocked with M.cephalus inside the mesh enclosure at densities of 0 (M0), 250 (M250), 500 (M500) and 800 (M800), respectively. Totally 7 phylums, 39 genera and 65 species of phytoplankton were identified, among which Chlorphyta (21 genera and 33 species) was the most abundance phylum, followed by Cyanophyta (7 genera and 17 species). The pond was dominated by Chlorella vulgaris (up to 50%). In general, phytoplankton abundance increased with increasing density of M.cephalus. In end stage, M250 attained the lowest abundance of blue-green algae and the highest abundance of green algae, both of which presented significant differences with M0 (P < 0.05). In addition, the diversity index of M250 was the highest among the treatments. Better growth of algae was observed in M500 and M800 (with higher mullet stocking densities) in early and middle stages, and in group M500 (with lower mullet stocking density) in late stage. The results indicated that mesh enclosure isolated pond with polyculture of L.vannamei and M.cephalus could achieve higher density of green algae, lower blue-green algae and higher diversity of phytoplankton community.
-
Keywords:
- shrimp /
- mullet /
- mesh enclosure /
- polyculture /
- phytoplankton
-
近年来凡纳滨对虾(Litopenaeus vannamei)养殖业面临的环境问题日益严峻,对虾白斑综合症(white spot syndrome virus,WSSV)、对虾早期死亡综合症(early mortality syndrome,EMS)等恶性病害频频出现[1-3]。随着虾病的蔓延,鱼虾混养模式病害少、水质优、效益高的特点被重新重视,研究表明凡纳滨对虾与金钱鱼(Scatophagus argus)[4]、红罗非鱼(Oreochromis spp.)[5]等鱼类品种混养均有较好的养殖效果。鲻(Mugil cephalus)广泛分布于热带亚热带地区,在淡水、半咸水及海水中均可生长,具有生长迅速、肉质鲜美、食性广杂、适应性强的特点,兼具较好的生态与经济价值[6],虾鲻混养已在华南对虾养殖区得到逐步推广和优化。
浮游植物、浮游动物对水质变化的响应极为灵敏,可以作为养殖池塘水体环境监测的指示生物[7],池塘水体环境可通过水体浮游植物生物量、丰度、多样性等指标的综合评价而获知[8]。当前中国虾鲻混养模式下有关水体环境的研究较少[9],尤其是在混养池塘浮游植物变化特征方面,仅见孔谦[10]对精养池塘的部分报道。鱼虾分隔混养通常采用双层围网或封闭网箱[11-13]进行,但此类方法虾池塘中央区域有机物积累去除率不高,对虾残饵也未得到充分利用,且混养鱼类捕食病、死虾的防病作用也受到限制。有鉴于此,笔者通过改进围网分隔养殖方式,对围绕池塘对虾-鲻网围分隔养殖系统浮游植物动态变化特征展开研究,以期为混养模式的改进和混养密度的优化提供科学依据。
1. 材料与方法
1.1 试验池塘与放养情况
试验于2012年5月~8月在茂名市电白冠利达科技生物养殖有限公司养殖场(21°32′ 35″ N,111° 23′05″ E)进行。池塘面积平均为3 330 m2,平均水深1.5 m,池塘内2台水车式增氧机呈对角分布。从养殖场随机挑选3个区域,每个区域内各有4口池塘(即4个不同的处理组)呈连片分布,不同处理组在上述3个区域内各设置1个重复。每个池塘进排水系统结构相同且各自独立。各组中凡纳滨对虾放养密度均为90尾· m-2,平均规格为0.8 cm · 尾-1;而各组池塘内鲻放苗量则依次为0(M0)、250(M250)、500(M500)和800(M800)尾,平均规格为0.3 kg · 尾-1,其中M0为对照组。放养鲻的各池塘中均搭建长方形围网(围网内水面约为800 m2),鲻均放养于围网之内,围网的网孔为方形,规格约2.5 cm×2.5 cm,对虾能自由出入网孔,而鲻则不能,围网的上边缘平齐于增氧机引起的池塘水流的内圈切线。另外,每个池塘围网外放养平均体质量为0.4 kg的埃及革胡子鲶(Clarias leather)150尾。各组投喂饲料为“粤海”牌南美白对虾饲料。池塘养殖现场效果如图 1所示。
![]() 图 1 对虾-鲻围网分隔混养现场效果图Figure 1. Real scene shooting of mesh enclosure isolated pond with polyculture of L.vannamei and M.cephalus
图 1 对虾-鲻围网分隔混养现场效果图Figure 1. Real scene shooting of mesh enclosure isolated pond with polyculture of L.vannamei and M.cephalus各试验池塘于2012年5月8日清塘,放苗及收获时间见表 1。
表 1 各处理组放苗及收获时间Table 1. Stocking and harvest time in different treatments处理组
treatment池塘
pond No.放苗日期stocking time 收获日期harvest time L M C L M C M0 504 05-18 ― 05-27 09-05 ― 09-05 M0 704 05-18 ― 05-27 09-12 ― 09-12 M0 1 205 05-18 ― 05-19 09-01 ― 09-01 M250 204 05-18 05-18 05-27 09-04 09-04 09-04 M250 805 05-28 05-26 05-27 09-06 09-06 09-06 M250 1 109 05-23 05-30 05-19 09-07 09-07 09-07 M500 705 05-18 05-21 05-27 08-29 08-29 08-29 M500 1 204 05-28 06-01 05-29 08-19 08-19 08-19 M500 104 05-18 05-18 05-27 09-15 09-15 09-15 M800 1 111 05-23 05-30 05-19 09-06 09-06 09-06 M800 706 05-18 05-20 05-27 08-20 08-20 08-20 M800 103 05-18 05-18 05-27 08-11 08-11 08-11 注:L、M、C分别代表凡纳滨对虾、鲻和胡子鲶;M0、M250、M500和M800代表鱼类放养数量(尾);后表同此
Note:L,M and C represent L.vannamei,M.cephalus and C.leather,respectively.M0, M250, M500 and M800 represent the fish stocking number.The same case in the following table.养殖结束后统计的各处理组对虾成活率、终体质量与产量见表 2。
表 2 各处理组对虾成活率、终体质量与产量Table 2. Survival, final weight and farming yield of shrimps in different treatments处理组
treatment成活率/%
survival终体质量/g
final weight产量/g·m-2
farming yieldM0 77.73±11.96 12.38±1.31 8.63±1.23 M250 69.70±8.43 14.30±2.37 9.05±2.34 M500 71.23±6.84 13.34±3.24 8.43±1.25 M800 74.93±4.78 11.02±1.11 7.41±0.53 1.2 样品的采集、测定与计算
于2012年6月2日~8月18日采集样品,平均采样周期为15 d。
浮游植物样品为池塘中央及四周的混合水样,充分混合后保存于1 L聚乙烯瓶中。水样经5%的福尔马林溶液固定后静置24~48 h,浓缩后采用浮游生物计数框镜检并计数,鉴定参考书目为《中国淡水藻志》。浮游植物的生态学特征采用香农多样性指数(H′)、Pielou物种均匀度指数(J)、优势度指数(Y)、Margalef丰富度指数(R)等指标进行研究。各指数计算公式[14]依次为:
$$ H^{\prime}=-\sum\limits_{i=1}^S P_i \log _2 P_i $$ (1) $$ J=\frac{H^{\prime}}{\log _2 S} $$ (2) $$ Y=f_i \times \frac{n_i}{N} $$ (3) $$ R=\frac{(S-1)}{\ln N} $$ (4) 式中Pi=ni/N;ni为物种i的个体数;N为群落样品总个体数;S为群落中的物种总数;fi为该种在该地区出现的频率。
浮游植物密度计算公式[15]为:
$$ N=\frac{n \times A \times V}{A C \times V_a} $$ (5) 式(5)中N为每升原水样中的浮游植物丰度(个· L-1),A为计数框面积(mm2),AC为计数面积(mm2),V为1 L原水样沉淀浓缩后的体积(mL),Va为计数框的体积(mL),n为计数所得的浮游植物数目。
浮游植物生物量通过显微镜下计数并测量各种类体积,按照比重1 : 1换算为干质量值来进行定量[16]。悬浮颗粒物[包括总悬浮颗粒物(TPM)、颗粒有机物(POM)、颗粒无机物(PIM)]样品经GF/F玻璃纤维滤膜抽滤后,采用灼烧法测定[17],叶绿素a(Chl-a)样品经90%的丙酮萃取后用紫外-可见分光光度计测定[18]。
此外,每次采样时水体总氨氮(NH4-N)、硝态氮(NO3-N)、亚硝态氮(NO2-N)、活性磷酸盐(PO4-P)、总氮(TN)、总磷(TP)均按《养殖水化学实验》[19]中所述方法采集和测定;pH、水温(T)、盐度(S)、溶解氧(DO)用多参数水质检测仪(YSI 556,美国产)实时测定。
1.3 数据分析
采用SPSS 13.0软件进行数据处理和统计分析。不同处理先做单因素方差分析(One-Way ANOVA),分析前先进行方差齐性检验,而后通过Duncan比对差异,以P<0.05作为差异显著水平。经SPSS 13.0处理数据后,用Origin 8.5绘制图表。
2. 结果
2.1 Chl-a质量浓度
养殖期间各组的Chl-a质量浓度前期较低(<100 μg · L-1),中期迅速升高到初期值2倍以上(>150 μg · L-1),经历7月中旬的短暂下降后,于8月初达到峰值,末期又有所下降(图 2)。整体而言,对照组M0的Chl-a质量浓度略高于鲻放养组,M800组的Chl-a质量浓度与M0较为接近。养殖后期鲻放养组Chl-a质量浓度随鲻密度增大而升高(M800>M500>M250)。
![]() 图 2 养殖期间叶绿素a质量浓度不同小写字母代表差异显著(P<0.05),下图同此Figure 2. Concentration of Chl-a during the culture period Different small letters indicate significant difference (P < 0.05). The same case in the following figures.
图 2 养殖期间叶绿素a质量浓度不同小写字母代表差异显著(P<0.05),下图同此Figure 2. Concentration of Chl-a during the culture period Different small letters indicate significant difference (P < 0.05). The same case in the following figures.2.2 浮游植物生物量及细胞丰度
养殖初期浮游植物细胞丰度较低,其后迅速升高,7月中旬稍有下降,其后在8月初达到峰值,养殖末期细胞丰度又有一定程度的下降(图 3)。整体而言,浮游植物细胞丰度随鲻放养量的增多而增大。浮游植物生物量初始值低于10 mg · L-1,此后快速增加,其整体变化迁移规律与细胞丰度变化相似:M0组在7月中旬生物量达到最大,此后变动幅度较小;M800最高值也出现在7月中旬,但此后下降趋势明显;M250和M500于8月初达到最高值,养殖末期各组差异不显著(P>0.05)。
![]() 图 3 养殖期间浮游植物生物量(A)和细胞丰度(B)Figure 3. Biomass (A) and density (B) of phytoplankton during culture period
图 3 养殖期间浮游植物生物量(A)和细胞丰度(B)Figure 3. Biomass (A) and density (B) of phytoplankton during culture period2.3 浮游植物鉴定结果
池塘中浮游植物共鉴定出7门39属65种(表 3)。绿藻门最多,为21属33种;其次为蓝藻门,7属17种。绿藻门的栅藻属(9种)和蓝藻门的色球藻属(9种)种类最多。其余门类种类较少:硅藻门5种、甲藻门、裸藻门、隐藻门均为3种,黄藻门仅1种。
表 3 池塘中浮游植物种类组成Table 3. Composition of microalgae species in ponds种类species 种类species 绿藻门Chlorophyta 蓝藻门Cyanophyta 小球藻属Chlorella 小球藻C.vulgaris 假鱼腥藻属Pseudanabaena 假鱼腥藻Pseudanabaena sp. 集球藻属Palmellococcus 集球藻Palmellococcus sp. 念珠藻属Nostoc 念珠藻Nostoc sp. 单针藻属Monoraphidium 单针藻Monoraphidium sp. 平裂藻属Merismopedia 小平裂藻M.minima 扁藻属Tetraselmis 扁藻T.Chui 细小平裂藻M.tenuissima 多芒藻属Golenkinia 多芒藻G.radiata 优美平裂藻M.elegans 集星藻属Actinastrum 集星藻Actinastrum sp. 色球藻属Chroococcus 膨胀色球藻C.turgidus 空星藻属Coelastrum 空星藻Coelastrum sp. 微小色球藻C.minutu 卵囊藻属Oocystis 细小卵囊藻O.pusilla 内生色球藻C.endophyticus 波吉卵囊藻O.borgei 粘连色球藻C.cohaerens 卵囊藻Oocystis sp. 湖沼色球藻C.limneticus 肾爿藻属Nephroselmis 肾爿藻Nephroselmis sp. 色球藻Chroococcus sp. 十字藻属Crucigenia 四足十字藻C.tetrapedia 小形色球藻C.minor 四角十字藻C.quadrata 易变色球藻C.varius 双月藻属Dicloster 双月藻Dicloster sp. 巨大色球藻C.giganteus 四棘藻属Treubaria 四棘藻T.triappendiculata 隐球藻属Aphanocapsa 细小隐球藻A.elachista 四角藻属Tetraedron 三角四角藻T.trigonum 束球藻属Coelosphaerium 圆胞束球藻C.naegelianum 塔胞藻属Pyramidomonas 娇柔塔胞藻P.delicatula 席藻属Phormidittmtertue 小席藻P.tentts 蹄形藻属Kirchneriella 蹄形藻K.lunaris 硅藻门Bacillariophyta 网球藻属Dictyosphaeria 网球藻D.cavernosa 小环藻属Cyclotella 小环藻Cyclotella sp. 微芒藻属Micractiai 微芒藻M.pusillum 卵形藻属Cocconeis 卵形藻Cocconeis sp. 纤维藻属Ankistrodesmus 纤维藻Ankistrodesmus sp. 舟形藻属Navicula 舟形藻Navicula sp. 镰形纤维藻A.falcatus 菱形藻属Nitzsehia 新月菱形藻N.closteztuma 小空星藻属Coelastrum 小空星藻C.microporum 三角藻属Triceratim 三角藻属Triceratim sp. 衣藻属Chlamydomonas 衣藻Chlamydomonas sp. 甲藻门Pyrrophyta 栅藻属Scenedesmus 四尾栅藻S.quadricauda 裸甲藻属Gymnodinium 裸甲藻G.aerucyinosum 斜生栅藻S.obliquus 多甲藻属Peridinium 多甲藻Peridinium sp. 二形栅藻S.dimorphus 角甲藻属Ceratium 角甲藻Ceratium sp. 被甲栅藻S.armatus 裸藻门Euglenophyta 二尾栅藻S.bicauda 裸藻属Euglena 鱼形裸藻E.pisciformis 栅藻Scenedesmus sp. 裸藻Euglena sp. 齿牙栅藻S.denticulatus 囊裸藻属Trachelomonas 囊裸藻Trachelomonas sp. 小刺栅藻S.microspina 隐藻门Cryptophyta 双对栅藻S.bijuba 隐藻属Cryptomonas 隐藻Cryptomonas sp. 黄藻门Xanthophyceae 卵形隐藻C.ovata 棘刺藻属Centritractus 棘刺藻Centritractus sp. 尖尾蓝隐藻C.acuta 整个养殖过程中,养殖池塘浮游植物以绿藻门和蓝藻门为主,尤其是蓝藻门,在养殖中后期其累计优势度占据了池塘50%以上(图 4)。黄藻门仅发现棘刺藻1种,且仅于养殖末期在M250组的204池塘出现,故未在图中绘出。硅藻门仅养殖前期零星出现一定优势度,而甲藻、裸藻、隐藻从未出现优势种,出现时间主要在养殖中后期。养殖初始,M0和M250 2组的绿藻丰度显著低于M500及M800组(P<0.05),而蓝藻丰度显著高过M500及M800组(P<0.05)。从前期到中期,M800组绿藻丰度快速下降,蓝藻则呈倍性增长,M250绿藻丰度逐渐增多;养殖后期各组的绿藻丰度均有一定程度的回升,蓝藻则相反;养殖末期M0组绿藻最低而蓝藻最高,M250组绿藻丰度最高而蓝藻最低,以上2组间蓝藻、绿藻丰度均存在显著差异(P<0.05)。
![]() 图 4 绿藻门(A)、蓝藻门(B)、硅藻门(C)、甲藻门(D)、裸藻门(E)和隐藻门(F)的优势度指数Figure 4. Accumulated Simpson dominance indices of Chlorophyta (A), Cyanophyta (B), Bacillariophyta (C), Pyrrophyta (D), Euglenophyta (E) and Cryptophyta(F)
图 4 绿藻门(A)、蓝藻门(B)、硅藻门(C)、甲藻门(D)、裸藻门(E)和隐藻门(F)的优势度指数Figure 4. Accumulated Simpson dominance indices of Chlorophyta (A), Cyanophyta (B), Bacillariophyta (C), Pyrrophyta (D), Euglenophyta (E) and Cryptophyta(F)处于绝对优势种(6次采样中累计2次以上优势度超过0.1)的绿藻和蓝藻变化规律与图 4-A、图 4-B的规律基本相同,但养殖末期各组间的差异不显著(图 5)。蓝藻门中,小球藻在养殖的所有时期均占据绝对优势(50%以上);蹄形藻在养殖初期在各组均有分布,随着养殖时间推移,主要出现于M800组中,而在养殖末期又在各组中有一定量的分布,且比较均匀;养殖初期网球藻在M500组中为优势种,其余时间则优势非常微弱,仅中后期在M0组中分布稍多。蓝藻门绝对优势种为细小平裂藻、细小隐球藻、微小色球藻。其中,微小色球藻主要在6月、7月出现,而在养殖的后期微乎其微。细小隐球藻在养殖前期为主要的优势种,随着养殖时间的推移,其优势逐渐被细小平裂藻所取代(尤其是7月中旬),而在养殖末期再次占据一定优势。
![]() 图 5 养殖期间绿藻门(A)和蓝藻门(B)绝对优势种Figure 5. Absolute dominant species of Chlorophyta (A) and Cyanophyta (B) during culture period
图 5 养殖期间绿藻门(A)和蓝藻门(B)绝对优势种Figure 5. Absolute dominant species of Chlorophyta (A) and Cyanophyta (B) during culture period2.4 浮游植物群落结构特征指数
养殖前期H′约为1.5,养殖后期上升至2.0左右,养殖中期M500组该指数显著低于其余组(P<0.05)。整体而言,养殖期间M0组的物种丰富度指数较高,仅在养殖后期略低于M250或M800。鲻放养组的物种均匀度指数常高于对照组M0(图 6)
![]() 图 6 香农多样性指数(A)、Margalef丰富度指数(B)及Pielou物种均匀度指数(C)Figure 6. Shannon′s diversity index (A), Margalef richness index (B) and Pielou evenness index (C)
图 6 香农多样性指数(A)、Margalef丰富度指数(B)及Pielou物种均匀度指数(C)Figure 6. Shannon′s diversity index (A), Margalef richness index (B) and Pielou evenness index (C)2.5 悬浮物变动特征
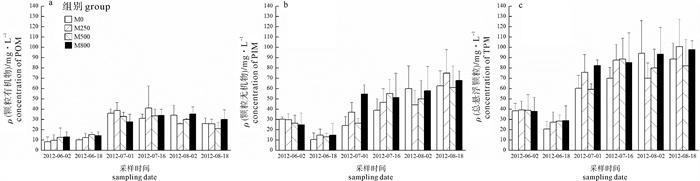
池塘水体中TPM主要存在形式为PIM,其质量浓度可达POM的2倍。养殖前期POM质量浓度较低,中期迅速升高,末期稍有下降;PIM在养殖1个月时质量浓度降至与POM接近,其后迅速升高。整体而言,与养殖初期相比,养殖末期TPM质量浓度平均升高一倍(图 7)。
![]() 图 7 养殖期间水体中颗粒有机物(A)、颗粒无机物(B)和总悬浮颗粒物质量浓度(C)Figure 7. Concentrations of particulate organic matter (A), particulate inorganic matter (B) and total particulate matter (C) in pond water during culture period
图 7 养殖期间水体中颗粒有机物(A)、颗粒无机物(B)和总悬浮颗粒物质量浓度(C)Figure 7. Concentrations of particulate organic matter (A), particulate inorganic matter (B) and total particulate matter (C) in pond water during culture period2.6 相关性分析
相关性分析显示(表 4),养殖期间浮游植物生物量与Chl-a质量浓度呈正相关,且相关系数表现为随鲻放养密度的增大而减小。除M250组(呈负相关)外,浮游植物细胞丰度与Chl-a质量浓度呈显著正相关(P<0.05),而与生物量则呈显著或非显著的正相关。相似地,除M250组外,浮游植物细胞丰度和TPM间表现为显著(P<0.05)或极显著(P<0.01)的正相关性,且相关系数随鲻放养密度的增大而下降。整体而言,M0组中各指标间的相关性最高,M500组次之,2组均为正相关;M800指标间的相关系数较低,而M250指标间的相关性普遍最低,且浮游植物Chl-a、细胞丰度、生物量与TPM及PIM呈负相关。
表 4 叶绿素a质量浓度、生物量、浮游植物细胞丰度及悬浮物间的相关系数Table 4. Pearson′s correlation coefficient of Chl-a concentration, biomass, phytoplankton density and suspended matter处理组
treatment指标
index叶绿素a
Chl-a生物量
biomass细胞丰度
phytoplankton density处理组
treatment指标
index叶绿素a
Chl-a生物量
biomass细胞丰度
phytoplankton densityM0 生物量 0.581* M500 生物量 0.466 细胞丰度 0.485* 0.521* 细胞丰度 0.481* 0.674** TPM 0.704** 0.491* 0.524* TPM 0.568* 0.323 0.641** POM 0.867** 0.608** 0.691** POM 0.701** 0.399 0.554* PIM 0.333 0.228 0.200 PIM 0.322 0.182 0.534* M250 生物量 0.476* M800 生物量 0.277 细胞丰度 -0.052 0.323 细胞丰度 0.573* 0.166 TPM -0.479* -0.201 -0.228 TPM 0.302 0.053 0.474* POM 0.319 0.065 -0.145 POM 0.412 -0.039 0.133 PIM -0.643** -0.246 -0.206 PIM 0.101 0.081 0.450 注:* *表示在0.01水平上显著相关,*表示在0.05水平上显著相关
Note: * *. very significant at 0.01 level;*. significant at 0.05 level3. 讨论
3.1 浮游植物细胞丰度变化特征
彭聪聪等[20]发现,对虾海水滩涂养殖池塘前、后期浮游植物丰度分别为5.12×104~95.41×104个· L-1、66.11×104~1.28×109个· L-1。与之相比,试验中养殖前期对虾单养池塘浮游植物个体丰度为27.29×107~32.39×107个· L-1,鲻混养组为4.21×107~40.61×107个· L-1;养殖中后期对虾单养池塘浮游植物个体丰度为44.60×107~93.28×107个· L-1,鲻混养组为66.91×107~128.58×107个· L-1,池塘浮游植物丰度远高于彭聪聪等[20]的研究结果,而与闫家国等[21]报道的珠江口对虾养殖池塘(44.7×107个· L-1)相近。养殖中后期池塘浮游植物密度随着鲻放养密度的增加而增大。混养池塘的浮游藻类密度通常高于对照M0。
3.2 悬浮物、Chl-a变化特征及相关性分析
TPM分为POM和PIM,前者主要为浮游动植物残体,后者主要为碎屑矿物、粘土矿物等,在内陆水体中沉积物的再悬浮使得矿物颗粒在TPM中占据较大比例[22]。对虾精养池塘[23](密度200尾·m-2)TPM质量浓度为20~76 mg · L-1,随养殖时间的推移而逐渐增加。对虾-罗非鱼混养池塘[24]中TPM质量浓度为(83.1±34.1)mg · L-1,POM质量浓度为(34.3±11.8)mg · L-1,二者显著相关,呈“高-低-低-高”的变化趋势。与之相比,试验各组TPM质量浓度前期低、中后期高,平均质量浓度<40 mg · L-1,主要以PIM形式存在,该结果较接近申玉春等[25]所构建的虾-鱼-贝-藻优化生态养殖区的结果(TPM平均45.4 mg · L-1)。对虾精养池塘[23]中Chl-a与总悬浮物间呈不显著正相关,而在半精养虾池[26](密度为75尾· m-2)中二者呈极显著正相关(P<0.01)。对照组M0的结果与上述半精养虾池相似,且相关性更高,表明对虾对水体沉积物再悬浮产生的影响较小。
养殖期间各组Chl-a质量浓度为(17.90±20.43)~(309.98±153.58) μg · L-1,与韦璐等[27]对上海奉贤区凡纳滨对虾养殖场的调查结果相近,而普遍高于LUCAS等[28]对热带虾池的研究结果。试验池塘Chl-a质量浓度与浮游植物生物量呈正相关性,与浮游植物细胞丰度(M250组除外)呈显著正相关(P<0.05),这符合一般规律:野外围隔试验水中,Chl-a与生物量一般呈显著正相关[29];当优势藻种稳定单一时,Chl-a与藻密度一般显著正相关,而藻种发生明显更替时,Chl-a与藻密度可能相关或不相关[30]。
3.3 浮游植物群落结构组成及多样性指数分析
养殖池塘盐度的高低对浮游植物群落结构有显著影响,盐度介于10~30的池塘中硅藻占优势,主要为舟形藻、桥弯藻等;而盐度较小(<5)的养殖池中以蓝藻占优势,主要为微囊藻、平裂藻、水花束丝藻、色球藻等[31]。珠江口对虾养殖池塘[21]盐度为3.36,以绿藻门居多(11属16种),其次为蓝藻门(6属8种),细小平裂藻的优势度较大。试验池塘盐度为2~3,浮游植物种类主要表现为与珠江口对虾养殖池塘相似的淡水群落特征。
通常情况下,藻类的多样性指数值越大,群落结构越复杂,稳定性越大,水质越净[32]。H′>1时群落多样性为正常,J>0.3时浮游植物多样性较好[33-34]。试验中各组的H′与J值均满足上述条件,且养殖中后期对照组M0数据接近闫家国等[21]的秋季虾池多样性数据(H′为2.35,J为0.47)及刘孝竹等[35]对珠海低盐度虾池的周年观测数据(H′平均2.5~3.2),浮游植物的种类数、丰度及多样性指数总体上表现为养殖前期低后期高的特征,表明养殖池塘浮游植物群落整体处于稳定状态。
王育红等[36]发现微型浮游植物群落H′受多因素影响,且群落中各种类的丰度为其直接决定因素。郭志勋等[37]发现正常虾池中浮游藻类H′和J值均明显高于发病虾池,蓝藻丰度则明显低于发病虾池。养殖末期M0组蓝藻丰度居各组最高水平而绿藻丰度则为最低,M250组则与之完全相反,绿藻丰度约为M0组的2倍之多,且M250组H′、R、J值均高于其余各组,此结果与该组对虾终体质量和养殖产量居于各组最高的结果一致。
3.4 虾-鲻混养在池塘藻相调控中的意义
尽管鲻偏好摄食硅藻,但也能适应不易消化的植物碎屑和蓝藻,有效降低海洋鱼类网箱下的沉积物有机质富集[39]。鲻鳃耙间距较小[39],可滤食20 μm左右的浮游植物,在鱼虾混养时可代替鲢的作用[40-41]。养殖末期鲻放养组绿藻高于M0,而蓝藻密度低于M0,表明鲻对浮游植物的滤食具有选择性,可有效控制蓝藻的增殖。M250组Chl-a质量浓度与浮游植物细胞丰度较差的相关性表明,该组浮游植物种类更替可能比其他组更为迅捷。
在水产养殖中常借助生态位调控来改变不良微藻的生长环境,利用种间竞争关系培养优良微藻以抑制有害微藻的大量滋生[42]。硅藻、金藻和绿藻是水生动物的天然优质饵料,其中绿藻在池塘水体净化方面又起着积极的作用。试验养殖池塘前期以硅藻为优势种群,绿藻次之,而养殖中后期,则以绿藻为主;但孔谦[10]的研究中单养池塘出现较多甲藻,而对虾-鲻混养池塘则出现较多金藻,其藻类组成更加多元化。
GILLES等[43]研究发现,小球藻可吸收罗非鱼[霍氏帚齿非鲫(Sarotherodon melanotheron heudelotii)]排泄的大部分氨,降低水中源自氨的毒性。张继平和郭照良[44]报道接种20个· L-1小球藻液后,水体中NO2-N降解效果极为显著(P<0.05)。因而,小球藻在养殖池塘中的快速繁殖有益于减轻对虾等养殖生物排泄及残饵分解产物对环境施加的压力。
试验结果表明,与对照组相比,放养适量密度的鲻能在养殖后期保持较高的绿藻尤其是小球藻密度,降低水体蓝藻丰度,有益于对虾池塘中后期水质的良好调控。养殖前中期,鲻密度较高组(尤其是M800)绿藻密度高、蓝藻密度低的特征较为明显;而到养殖后期(特别是末期)鲻密度略低的M250组的该特征更加明显。因此,网围分隔方式进行对虾与鲻混养时,可以尝试前期放入较高密度的鲻,中期适当分鱼降低密度的措施来维持对虾养殖期间较好的藻相分布特征[45]。
-
![]()
图 1 对虾-鲻围网分隔混养现场效果图
Figure 1. Real scene shooting of mesh enclosure isolated pond with polyculture of L.vannamei and M.cephalus
![]()
图 2 养殖期间叶绿素a质量浓度不同小写字母代表差异显著(P<0.05),下图同此
Figure 2. Concentration of Chl-a during the culture period Different small letters indicate significant difference (P < 0.05). The same case in the following figures.
![]()
图 3 养殖期间浮游植物生物量(A)和细胞丰度(B)
Figure 3. Biomass (A) and density (B) of phytoplankton during culture period
![]()
图 4 绿藻门(A)、蓝藻门(B)、硅藻门(C)、甲藻门(D)、裸藻门(E)和隐藻门(F)的优势度指数
Figure 4. Accumulated Simpson dominance indices of Chlorophyta (A), Cyanophyta (B), Bacillariophyta (C), Pyrrophyta (D), Euglenophyta (E) and Cryptophyta(F)
![]()
图 5 养殖期间绿藻门(A)和蓝藻门(B)绝对优势种
Figure 5. Absolute dominant species of Chlorophyta (A) and Cyanophyta (B) during culture period
![]()
图 6 香农多样性指数(A)、Margalef丰富度指数(B)及Pielou物种均匀度指数(C)
Figure 6. Shannon′s diversity index (A), Margalef richness index (B) and Pielou evenness index (C)
![]()
图 7 养殖期间水体中颗粒有机物(A)、颗粒无机物(B)和总悬浮颗粒物质量浓度(C)
Figure 7. Concentrations of particulate organic matter (A), particulate inorganic matter (B) and total particulate matter (C) in pond water during culture period
表 1 各处理组放苗及收获时间
Table 1 Stocking and harvest time in different treatments
处理组
treatment池塘
pond No.放苗日期stocking time 收获日期harvest time L M C L M C M0 504 05-18 ― 05-27 09-05 ― 09-05 M0 704 05-18 ― 05-27 09-12 ― 09-12 M0 1 205 05-18 ― 05-19 09-01 ― 09-01 M250 204 05-18 05-18 05-27 09-04 09-04 09-04 M250 805 05-28 05-26 05-27 09-06 09-06 09-06 M250 1 109 05-23 05-30 05-19 09-07 09-07 09-07 M500 705 05-18 05-21 05-27 08-29 08-29 08-29 M500 1 204 05-28 06-01 05-29 08-19 08-19 08-19 M500 104 05-18 05-18 05-27 09-15 09-15 09-15 M800 1 111 05-23 05-30 05-19 09-06 09-06 09-06 M800 706 05-18 05-20 05-27 08-20 08-20 08-20 M800 103 05-18 05-18 05-27 08-11 08-11 08-11 注:L、M、C分别代表凡纳滨对虾、鲻和胡子鲶;M0、M250、M500和M800代表鱼类放养数量(尾);后表同此
Note:L,M and C represent L.vannamei,M.cephalus and C.leather,respectively.M0, M250, M500 and M800 represent the fish stocking number.The same case in the following table. 下载: 导出CSV
下载: 导出CSV
表 2 各处理组对虾成活率、终体质量与产量
Table 2 Survival, final weight and farming yield of shrimps in different treatments
处理组
treatment成活率/%
survival终体质量/g
final weight产量/g·m-2
farming yieldM0 77.73±11.96 12.38±1.31 8.63±1.23 M250 69.70±8.43 14.30±2.37 9.05±2.34 M500 71.23±6.84 13.34±3.24 8.43±1.25 M800 74.93±4.78 11.02±1.11 7.41±0.53
下载: 导出CSV
表 3 池塘中浮游植物种类组成
Table 3 Composition of microalgae species in ponds
种类species 种类species 绿藻门Chlorophyta 蓝藻门Cyanophyta 小球藻属Chlorella 小球藻C.vulgaris 假鱼腥藻属Pseudanabaena 假鱼腥藻Pseudanabaena sp. 集球藻属Palmellococcus 集球藻Palmellococcus sp. 念珠藻属Nostoc 念珠藻Nostoc sp. 单针藻属Monoraphidium 单针藻Monoraphidium sp. 平裂藻属Merismopedia 小平裂藻M.minima 扁藻属Tetraselmis 扁藻T.Chui 细小平裂藻M.tenuissima 多芒藻属Golenkinia 多芒藻G.radiata 优美平裂藻M.elegans 集星藻属Actinastrum 集星藻Actinastrum sp. 色球藻属Chroococcus 膨胀色球藻C.turgidus 空星藻属Coelastrum 空星藻Coelastrum sp. 微小色球藻C.minutu 卵囊藻属Oocystis 细小卵囊藻O.pusilla 内生色球藻C.endophyticus 波吉卵囊藻O.borgei 粘连色球藻C.cohaerens 卵囊藻Oocystis sp. 湖沼色球藻C.limneticus 肾爿藻属Nephroselmis 肾爿藻Nephroselmis sp. 色球藻Chroococcus sp. 十字藻属Crucigenia 四足十字藻C.tetrapedia 小形色球藻C.minor 四角十字藻C.quadrata 易变色球藻C.varius 双月藻属Dicloster 双月藻Dicloster sp. 巨大色球藻C.giganteus 四棘藻属Treubaria 四棘藻T.triappendiculata 隐球藻属Aphanocapsa 细小隐球藻A.elachista 四角藻属Tetraedron 三角四角藻T.trigonum 束球藻属Coelosphaerium 圆胞束球藻C.naegelianum 塔胞藻属Pyramidomonas 娇柔塔胞藻P.delicatula 席藻属Phormidittmtertue 小席藻P.tentts 蹄形藻属Kirchneriella 蹄形藻K.lunaris 硅藻门Bacillariophyta 网球藻属Dictyosphaeria 网球藻D.cavernosa 小环藻属Cyclotella 小环藻Cyclotella sp. 微芒藻属Micractiai 微芒藻M.pusillum 卵形藻属Cocconeis 卵形藻Cocconeis sp. 纤维藻属Ankistrodesmus 纤维藻Ankistrodesmus sp. 舟形藻属Navicula 舟形藻Navicula sp. 镰形纤维藻A.falcatus 菱形藻属Nitzsehia 新月菱形藻N.closteztuma 小空星藻属Coelastrum 小空星藻C.microporum 三角藻属Triceratim 三角藻属Triceratim sp. 衣藻属Chlamydomonas 衣藻Chlamydomonas sp. 甲藻门Pyrrophyta 栅藻属Scenedesmus 四尾栅藻S.quadricauda 裸甲藻属Gymnodinium 裸甲藻G.aerucyinosum 斜生栅藻S.obliquus 多甲藻属Peridinium 多甲藻Peridinium sp. 二形栅藻S.dimorphus 角甲藻属Ceratium 角甲藻Ceratium sp. 被甲栅藻S.armatus 裸藻门Euglenophyta 二尾栅藻S.bicauda 裸藻属Euglena 鱼形裸藻E.pisciformis 栅藻Scenedesmus sp. 裸藻Euglena sp. 齿牙栅藻S.denticulatus 囊裸藻属Trachelomonas 囊裸藻Trachelomonas sp. 小刺栅藻S.microspina 隐藻门Cryptophyta 双对栅藻S.bijuba 隐藻属Cryptomonas 隐藻Cryptomonas sp. 黄藻门Xanthophyceae 卵形隐藻C.ovata 棘刺藻属Centritractus 棘刺藻Centritractus sp. 尖尾蓝隐藻C.acuta
下载: 导出CSV
表 4 叶绿素a质量浓度、生物量、浮游植物细胞丰度及悬浮物间的相关系数
Table 4 Pearson′s correlation coefficient of Chl-a concentration, biomass, phytoplankton density and suspended matter
处理组
treatment指标
index叶绿素a
Chl-a生物量
biomass细胞丰度
phytoplankton density处理组
treatment指标
index叶绿素a
Chl-a生物量
biomass细胞丰度
phytoplankton densityM0 生物量 0.581* M500 生物量 0.466 细胞丰度 0.485* 0.521* 细胞丰度 0.481* 0.674** TPM 0.704** 0.491* 0.524* TPM 0.568* 0.323 0.641** POM 0.867** 0.608** 0.691** POM 0.701** 0.399 0.554* PIM 0.333 0.228 0.200 PIM 0.322 0.182 0.534* M250 生物量 0.476* M800 生物量 0.277 细胞丰度 -0.052 0.323 细胞丰度 0.573* 0.166 TPM -0.479* -0.201 -0.228 TPM 0.302 0.053 0.474* POM 0.319 0.065 -0.145 POM 0.412 -0.039 0.133 PIM -0.643** -0.246 -0.206 PIM 0.101 0.081 0.450 注:* *表示在0.01水平上显著相关,*表示在0.05水平上显著相关
Note: * *. very significant at 0.01 level;*. significant at 0.05 level
下载: 导出CSV
-
[1] DASTRIDAR P G, MALLIK A, MANDAL N. Contribution of shrimp disease research to the development of the shrimp aquaculture industry: an analysis of the research and innovation structure across the countries[J]. Scientometrics, 2013, 97(3): 659-674. doi: 10.1007/s11192-013-0977-9
[2] TENDENCIA E A, BOSMA R H, VERRETH J A J. White spot syndrome virus (WSSV) risk factors associated with shrimp farming practices in polyculture and monoculture farms in the Philippines[J]. Aquaculture, 2011, 311(1): 87-93. https://xueshu.baidu.com/usercenter/paper/show?paperid=6daa1eea5a83a4e4f0ab7d3d279896c3&site=xueshu_se
[3] TEIXEIRA-LOPES M A, VIEIRA-GIRÃO P R N, FREIRE J E C, et al. Natural co-infection with infectious hypodermal and hematopoietic necrosis virus (IHHNV) and infectious myonecrosis virus (IMNV) in Litopenaeus vannamei in Brazil[J]. Aquaculture, 2011, 312(1): 212-216. https://www.researchgate.net/publication/229417018_Natural_co-infection_with_infectious_hypodermal_and_hematopoietic_necrosis_virus_IHHNV_and_infectious_myonecrosis_virus_IMNV_in_Litopenaeus_vannamei_in_Brazil
[4] LIU L P, HU Z X, DAI X L, et al. Effects of addition of maize starch on the yield, water quality and formation of bioflocs in an integrated shrimp culture system[J]. Aquaculture, 2014, 418/419: 79-86. doi: 10.1016/j.aquaculture.2013.10.005
[5] YUAN D, YI Y, YAKUPITIYAGE A, et al. Effects of addition of red tilapia (Oreochromis spp. ) at different densities and sizes on production, water quality and nutrient recovery of intensive culture of white shrimp (Litopenaeus vannamei) in cement tanks[J]. Aquaculture, 2010, 298(3): 226-238. https://xueshu.baidu.com/usercenter/paper/show?paperid=409ac2a148afa4068d547ac80e4da2e4&site=xueshu_se&hitarticle=1
[6] WHITFIELD A K, PANFILI J, DURAND J D. A global review of the cosmopolitan flathead mullet Mugil cephalus Linnaeus 1758 (Teleostei: Mugilidae), with emphasis on the biology, genetics, ecology and fisheries aspects of this apparent species complex[J]. Rev Fish Biol Fisher, 2012, 22(3): 641-681. doi: 10.1007/s11160-012-9263-9
[7] CASÉ M, LEÇA E E, LEITÃO S N, et al. Plankton community as an indicator of water quality in tropical shrimp culture ponds[J]. Mar Pol Bul, 2008, 56(7): 1343-1352. doi: 10.1016/j.marpolbul.2008.02.008
[8] BURFORD M. Phytoplankton dynamics in shrimp ponds[J]. Aquacult Res, 1997, 28(5): 351-360. doi: 10.1111/j.1365-2109.1997.tb01052.x
[9] 邱丽华, 吴进锋, 张汉华, 等. 海水池塘鲻鱼、斑节对虾混养的初步研究[J]. 湛江海洋大学学报, 2000, 20(4): 69-71. https://xueshu.baidu.com/usercenter/paper/show?paperid=dffb906957e8f9ca904ab4ea9b207836&site=xueshu_se [10] 孔谦. 凡纳滨对虾与鲻鱼混养中精养池的理化生物因子的研究[D]. 湛江: 广东海洋大学, 2010. https://xueshu.baidu.com/usercenter/paper/show?paperid=a2224990faf6c19646eda742ee710deb&site=xueshu_se&hitarticle=1 [11] 董双林, 介子林, 赵夕旦, 等. 对虾与滤食性鱼类分隔式混合养殖的方法: 中国, CN1736182[P]. 2006-02-22. https://xueshu.baidu.com/usercenter/paper/show?paperid=b019489a13d070eebd7e7c22a75d62da&site=xueshu_se [12] 虞为, 李卓佳, 朱长波, 等. 凡纳滨对虾池塘设置网箱养殖罗非鱼研究[J]. 广东农业科学, 2011, 38(15): 4-9. https://d.wanfangdata.com.cn/periodical/Ch9QZXJpb2RpY2FsQ0hJTmV3UzIwMjQxMTA1MTcxMzA0Eg9nZG55a3gyMDExMTUwMDIaCDFmaDM0bjll [13] 王吉桥, 李德尚, 董双林, 等. 鲈-中国对虾-罗非鱼混养的实验研究[J]. 中国水产科学, 2000, 7(4): 37-41. https://www.fishscichina.com/zgsckx/article/abstract/4291?st=article_issue [14] 孙军, 田伟. 2009年春季长江口及其邻近水域浮游植物——物种组成与粒级叶绿素a[J]. 应用生态学报, 2011, 22(1): 235-242. https://xueshu.baidu.com/usercenter/paper/show?paperid=abc080005c133372bac459e620c20a4a&site=xueshu_se&hitarticle=1 [15] 金相灿, 屠清瑛. 湖泊富营养化调查规范[M]. 2版. 北京: 中国环境科学出版社, 1990: 303-316. https://xueshu.baidu.com/usercenter/paper/show?paperid=f633a3a7aabd1b2ca44d669e3bb600ae&site=xueshu_se [16] 张觉民, 何志辉. 内陆水域渔业自然资源调查手册[M]. 北京: 农业出版社, 1991: 26-27. https://xueshu.baidu.com/usercenter/paper/show?paperid=6d65800fce421ed0afd788c25a57dfe8&site=xueshu_se&hitarticle=1 [17] 方志山. 综合水产养殖池悬浮颗粒物的动态变化研究[J]. 厦门大学学报: 自然科学版, 2007, 46(1), 131-134. doi: 10.3321/j.issn:0438-0479.2007.01.029 [18] 陈宇炜, 高锡云. 浮游植物叶绿素a含量测定方法的比较测定[J]. 湖泊科学, 2000, 12(2): 185-188. doi: 10.3321/j.issn:1003-5427.2000.02.015 [19] 雷衍之. 养殖水环境化学实验[M]. 北京: 中国农业出版社, 2006: 31-60. https://xueshu.baidu.com/usercenter/paper/show?paperid=aa98806ad2236b68a42111604e8403be&site=xueshu_se&hitarticle=1 [20] 彭聪聪, 李卓佳, 曹煜成, 等. 粤西凡纳滨对虾海水滩涂养殖池塘浮游微藻群落结构特征[J]. 渔业科学进展, 2011, 32(4): 117-125. https://xueshu.baidu.com/usercenter/paper/show?paperid=2f51de80e45bd4abbd85212409a11cd8&site=xueshu_se&hitarticle=1 [21] 闫家国, 黄洪辉, 齐占会, 等. 珠江口养殖池塘秋季浮游植物群落特征研究[J]. 广东农业科学, 2012, 39(16): 138-141. https://xueshu.baidu.com/usercenter/paper/show?paperid=1add618c5bb6dfbd8906620354e0da89&site=xueshu_se&hitarticle=1 [22] BABIN M, STRAMSKI D, FERRARI G M, et al. Variations in the light absorption coefficients of phytoplankton, nonalgal particles, and dissolved organic matter in coastal waters around Europe[J]. J Geo Res, 2003, 108(C7): 1-4. https://xueshu.baidu.com/usercenter/paper/show?paperid=ba6295b8e162f61906d5953851161e39&site=xueshu_se&hitarticle=1
[23] 陈金玲, 赖秋明, 苏树叶, 等. 精养虾池主要生态因子变化特点与相关性分析[J]. 南方水产科学, 2012, 8(4): 49-56. doi: 10.3969/j.issn.2095-0780.2012.04.008 [24] 粟丽, 朱长波, 张汉华, 等. 对虾池塘网箱养殖罗非鱼期间水体悬浮颗粒物的动态及对罗非鱼生长和存活的影响[J]. 中国水产科学, 2012, 19(2): 256-264. https://xueshu.baidu.com/usercenter/paper/show?paperid=1e600vw0s54y0cs0et5k0jq0fs476356&site=xueshu_se&hitarticle=1 [25] 申玉春, 熊邦喜, 叶富良, 等. 虾-鱼-贝-藻生态优化养殖及其水质生物调控技术研究[J]. 生态学杂志, 2005, 24(6): 613-618. https://xueshu.baidu.com/usercenter/paper/show?paperid=afe308d52ee8fb6db9384d518b529b25&site=xueshu_se&hitarticle=1 [26] 江敏, 余根鼎, 戴习林, 等. 凡纳滨对虾养殖塘叶绿素a与水质因子的多元回归分析[J]. 水产学报, 2010, 34(11): 1712-1718. https://xueshu.baidu.com/usercenter/paper/show?paperid=03acce35f120af708f893ba198f10b5c&site=xueshu_se&hitarticle=1 [27] 韦璐, 江敏, 余根鼎, 等. 凡纳滨对虾养殖塘叶绿素a与水质因子主成分多元线性回归分析[J]. 中国水产科学, 2012, 19(4): 620-625. https://xueshu.baidu.com/usercenter/paper/show?paperid=1g1b0jd0d27a0en0r05t0j7028079865&site=xueshu_se [28] LUCAS R, COURTIES C, HERBLAND A, et al. Eutrophication in a tropical pond: Understanding the bacterioplankton and phytoplankton dynamics during a vibriosis outbreak using flow cytometric analyses[J]. Aquaculture, 2010, 310(1): 112-121. https://xueshu.baidu.com/usercenter/paper/show?paperid=454537aa5549edd5167588495ea820c0&site=xueshu_se&hitarticle=1
[29] 杜胜蓝, 黄岁樑, 臧常娟, 等. 浮游植物现存量表征指标间相关性研究Ⅰ: 叶绿素a与生物量[J]. 水资源与水工程学报, 2011, 22(1): 40-44. https://xueshu.baidu.com/usercenter/paper/show?paperid=f77a4880f8f53e1c253b670727c7f4f9&site=xueshu_se&hitarticle=1 [30] 杜胜蓝, 黄岁樑, 臧常娟, 等. 浮游植物现存量表征指标间相关性研究Ⅱ: 叶绿素a与藻密度[J]. 水资源与水工程学报, 2011, 22(2): 44-49. https://xueshu.baidu.com/usercenter/paper/show?paperid=3b263897a239c0cc35bba9620c8bc1fa&site=xueshu_se [31] 谢立民, 林小涛, 许忠能, 等. 不同类型虾池的理化因子及浮游植物群落的调查[J]. 生态科学, 2003, 22(1): 34-37. https://xueshu.baidu.com/usercenter/paper/show?paperid=9761bdf1302b2400ce0cedd96c810315&site=xueshu_se&hitarticle=1 [32] SUN J, LIU D Y. The application of diversity indices in marine phytoplankton studies[J]. Acta Ocean Sin, 2003, 26(1): 62-75. https://xueshu.baidu.com/usercenter/paper/show?paperid=e6c22203026584fbb7f72001a0292109&site=xueshu_se&hitarticle=1
[33] 马建新, 郑振虎, 李云平, 等. 莱州湾浮游植物分布特征[J]. 海洋湖沼通报, 2002(4): 64-67. https://xueshu.baidu.com/usercenter/paper/show?paperid=1c5s0240b34s08s0241d0880m1610705&site=xueshu_se&hitarticle=1 [34] 李广楼, 陈碧鹃, 崔毅, 等. 莱州湾浮游植物的生态特征[J]. 中国水产科学, 2006, 13(2): 292-299. https://xueshu.baidu.com/usercenter/paper/show?paperid=c4911a3c1c2c563803744a3bf462af39&site=xueshu_se&hitarticle=1 [35] 刘孝竹, 李卓佳, 曹煜成, 等. 低盐度养殖池塘常见浮游微藻的种类组成, 数量及优势种群变动[J]. 南方水产, 2009, 5(1): 9-16. doi: 10.3969/j.issn.1673-2227.2009.01.002 [36] 王育红, 杨建敏, 张明亮, 等. 黄河三角洲海参养殖池塘微型浮游植物多样性研究[J]. 海洋与湖沼, 2013, 44(2): 415-420. https://xueshu.baidu.com/usercenter/paper/show?paperid=e76661e7d9ec7e7f2f051425229e9bce&site=xueshu_se&hitarticle=1 [37] 郭志勋, 张汉华, 冯娟, 等. 正常与发病对虾养殖池塘浮游藻种类组成和异养细菌数量比较[J]. 生态科学, 2011, 30(4): 369-374. https://xueshu.baidu.com/usercenter/paper/show?paperid=84e4b425e81ea540392057aec5e0230c&site=xueshu_se&hitarticle=1 [38] LUPATSCH I, KATZ T, ANGEL D L. Assessment of the removal efficiency of fish farm effluents by grey mullets: a nutritional approach[J]. Aquacult Res, 2003, 34(15): 1367-1377. doi: 10.1111/j.1365-2109.2003.00954.x
[39] HICKLING C F. A contribution to the natural history of the English grey mullets (Pisces, Mugilidae)[J]. J Mar Biol Ass UK, 1970, 50(3): 609-633. doi: 10.1017/S0025315400004914
[40] 陆开宏, 王扬才, 蔡惠凤. 2种摄藻鱼消化酶活性及消化器官组织形态比较研究[J]. 水利渔业, 2005, 25(5): 37-39. https://xueshu.baidu.com/usercenter/paper/show?paperid=4c4c9ddbec6e2e01386b6f34f9d21ad9&site=xueshu_se [41] CARDONA L. Microalgae selection by mullets (Mugil cephalus and Liza ramada) in Israeli semi-intensive fishponds[J]. Isr J Aquacult-Bamid, 1996, 48(3): 165-173.
[42] 张汉华, 李卓佳, 郭志勋, 等. 有益微生物对海水养虾池浮游生物生态特征的影响研究[J]. 南方水产科学, 2005, 1(2): 7-14. https://xueshu.baidu.com/usercenter/paper/show?paperid=1be5627337d9488d2531347ae2b5f151&site=xueshu_se&hitarticle=1 [43] GILLES S, LACROIX G, CORBIN D, et al. Mutualism between euryhaline tilapia Sarotherodon melanotheron heudelotii and Chlorella sp. : implications for nano-algal production in warmwater phytoplankton-based recirculating systems[J]. Aquacult Eng, 2008, 39(2): 113-121. https://www.semanticscholar.org/paper/Mutualism-between-euryhaline-tilapia-Sarotherodon-%E2%80%93-Gilles-Lacroix/4d8ccb9a97996e601b664c82360868105a9a4462
[44] 张继平, 郭照良. 小球藻对降低南美白对虾养殖水体中亚硝酸盐氮含量的研究[J]. 水产科学, 2006, 25(10): 517-519. https://xueshu.baidu.com/usercenter/paper/show?paperid=037ac6d42959d077d28af9412be27d00&site=xueshu_se&hitarticle=1 [45] 朱长波. 郭永坚. 颉晓勇, 等. 凡纳滨对虾-鲻网围分隔混奍模式下经济与生态效益评价[J]. 南方水产科学, 2014, 10(4): 1-8. doi: 10.3969/j.issn.2095-0780.2014.04.001 -
期刊类型引用(10)
1. 方民杰. 多营养层次养殖的海水池塘浮游植物群落特征. 中国农学通报. 2023(02): 135-141 .  百度学术
百度学术
2. 张雷鸣,原居林,倪蒙,刘梅,郭爱环,练青平,王海洋,顾志敏. 两种池塘养殖模式水质因子和浮游植物群落比较分析. 浙江农业学报. 2020(02): 317-326 . 百度学术
3. 陈思,李艺彤,蔡文贵,陈海刚,田斐,张林宝,张喆,郭志勋. 虾蟹混养池塘浮游植物群落结构的变化特征. 南方水产科学. 2020(03): 79-85 . 本站查看
4. 江天棋,张扬,姜亚洲,孙鹏,凌建忠,唐保军. 不同浮游植物叶绿素a提取方法的比较研究. 海洋渔业. 2020(04): 445-453 . 百度学术
5. 王旭娜,江敏,钟锐,缪一恒,吴涵长,余苗苗,吴丹. 凡纳滨对虾养殖池塘中浮游植物群落结构与水质因子的关系. 水产学报. 2018(11): 1787-1803 . 百度学术
6. 孙志伟,邱丽华,曹煜成,李卓佳,胡晓娟,苏浩昌,蔡卓平,文国樑. 蓝藻水华对水产养殖业影响的研究进展. 生态科学. 2017(01): 231-235 . 百度学术
7. 李俊伟,朱长波,颉晓勇,郭永坚,陈素文,常华,张宁,曾林,胡瑞萍. 池塘分区养殖系统对水质和浮游植物群落结构的影响. 生态科学. 2017(06): 114-124 . 百度学术
8. 高宏伟,李文香,王博涵,李晓春,白海锋,袁永锋,杨希. 长丰鲢夏花培育池塘浮游植物群落结构调查研究. 现代农业科技. 2015(21): 256-257 . 百度学术
9. 郭永坚,罗昭林,阴晓丽,李俊伟,朱长波,颉晓勇. 4种饲料投喂对鲻(Mugil cephalus)生长及鱼体组成的影响. 渔业科学进展. 2015(01): 48-53 . 百度学术
10. 粟丽,朱长波,陈素文. 混养罗非鱼对凡纳滨对虾养殖围隔中悬浮颗粒物和弧菌数量的影响. 南方水产科学. 2015(06): 19-26 . 本站查看
其他类型引用(3)






计量
- 文章访问数: 3425
- HTML全文浏览量: 168
- PDF下载量: 2202
- 被引次数: 13
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
