
Seasonal variation of biofouling communities on concrete
-
摘要:
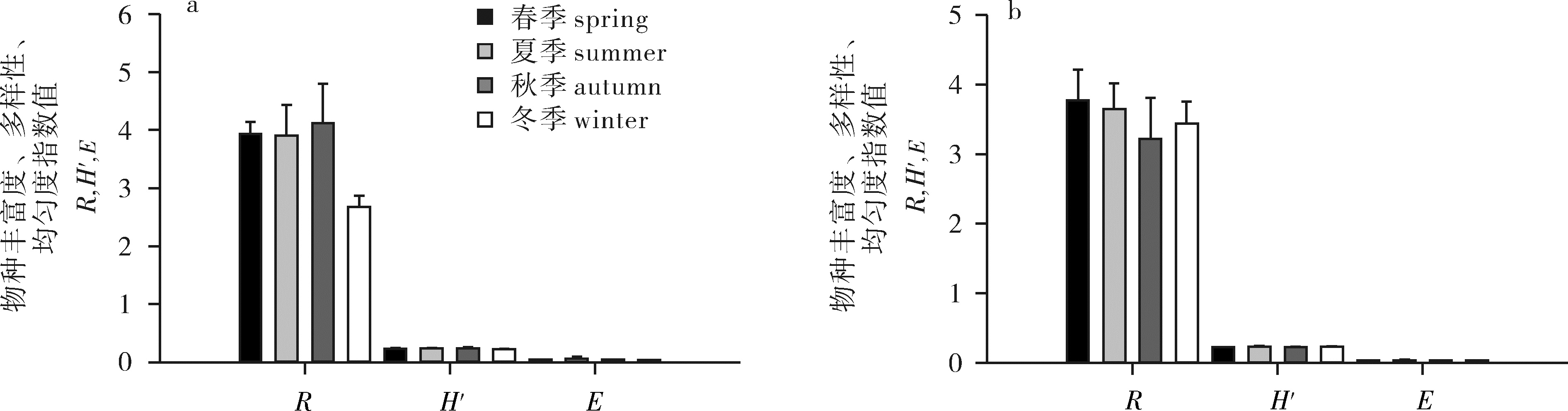
2008年分4个季节对深圳大亚湾混凝土礁体和铁质礁体上附着生物进行调查。结果显示,生物群落的种类组成存在季节差异。混凝土礁体秋季群落组成最丰富(60种),其次为夏季(58种)、春季(56种)和冬季(37种),其中仅23种为群落的共有种。不同季节群落的生物量为冬季﹥秋季﹥春季﹥夏季,而物种优势度(Y)差异不显著;不同季节的物种丰富度指数(R)为秋季>夏季>春季>冬季,物种多样性指数(H′)为夏季>秋季>春季>冬季,均匀度指数(E)为夏季>秋季>春季>冬季。铁质礁体春季群落组成最丰富(70种),其次是夏季(57种)、秋季(51种)和冬季(41种),其中仅17种为群落的共有种。不同季节群落的生物量为春季﹥秋季﹥夏季﹥冬季,而物种Y差异不显著;不同季节的物种R为春季>夏季>冬季>秋季,H′为冬季>夏季>秋季>春季,E为冬季>夏季>秋季>春季。然而,混凝土礁体和铁质礁体附着生物群落的R、H′和E季节变化不显著,且2种材质礁体相比,附着生物群落变化不明显。
Abstract:We investigated the biofouling communities on concrete and steel reefs in Daya Bay, Shenzhen, in May, July, October and December, 2008. Seasonal variation of species composition of biofouling communities was significant.On concrete reefs, 60 species were found in autumn, 58 in summer, 56 in spring and 37 in winter; only 23 species were found simultaneously in four seasons. The descending order of biomass of biofouling communities was winter>autumn>spring>summer. No significant difference in seasonal species dominance was found. The index of richness (R) was autumn>summer>spring>winter; the index of Shannon-Wiener (H′) was summer>autumn>spring>winter; the index of evenness (E) was summer>autumn>spring>winter. On steel reefs, 70 species were found in spring, 57 in summer, 51 in autumn and 41 in winter; only 17 species were found simultaneously in four seasons. The biomass of biofouling communities was spring>autumn>summer>winter. No significant difference in seasonal species dominance (Y) was found. The R value was spring>summer>winter>autumn; the H′ value was winter>summer>autumn>spring; the E value was winter>summer>autumn>spring. However, no significant difference in R, H′ and E values was found, neither was biofouling communities between concrete and steel reefs.
-
Keywords:
- seasonal variation /
- Daya Bay /
- biofouling communities /
- concrete reefs /
- steel reefs
-
广东鲂 (Megalobrama terminalis) 是一种江河洄游鱼类,在我国南方水系中占有重要的渔业生产地位[1-2]。我国南方水系复杂多样,并经历了多次河流袭夺,而气候变化导致的海平面波动是形成其特有河网系统的重要因素之一[3]。由于广东鲂对淡水环境依赖性较强,因此,海平面波动产生的地理隔离,对广东鲂种群的遗传结构产生了一定影响。Chen等[3]指出3个广东鲂地理遗传种群分别为珠江、漠阳江和海南岛万泉河种群,并发现广东鲂种群在不同的淡水栖息地中表现出明显的适应性分化。刘凯等[4]研究表明,受地理分布、食物来源、遗传特征和栖息环境等因素的叠加影响,鱼类的不同地理种群在肌肉营养成分上能够产生相对稳定的变异。然而,不同广东鲂地理种群肌肉营养成分是否存在显著差异尚不清楚。目前,对广东鲂研究多在于早期资源、资源捕捞量、性腺发育、繁殖策略、消化生理等方面[1-3,5-8],针对不同广东鲂种群肌肉营养成分和能量密度的研究尚未见报道。由于人类活动的不断加强 (如水利水电工程、航道治理、水污染、过度捕捞等),珠江野生广东鲂种群数量持续下降[2,7-8],漠阳江和万泉河广东鲂种群则呈现规模小、片段化分布特征,在其他陆河河流如榕江、鉴江、韩江中已难以监测到野生样本。因此,本研究测定了万泉河、漠阳江、西江广东鲂种群肌肉营养成分和能量密度,探究不同地理广东鲂种群肌肉营养成分差异,以期充实鱼类营养学和能量生态学研究材料,也为不同广东鲂的野生地理种群的分类保护和合理利用提供科学依据。
1. 材料与方法
1.1 样本采集
2020年6—7月分别于海南省琼海市万泉河琼海段 (QH, 110°27"36'E, 19°12"36'N)、广东省阳江市漠阳江段 (YJ, 111°42"2'E, 22°48"7'N) 以及广东省肇庆市珠江干流 (西江) 肇庆江段 (ZQ, 112°24"35'E, 23°5"24'N) 采集到150尾广东鲂,各采样点50尾 (雌、雄各25尾) 。采用哈希水质分析仪测量取样点的水温、盐度、溶解氧 (Dissolved oxygen, DO) 和pH,并测量样品的体长和体质量。采样站位的环境信息和样本生物学信息见表1。采用液氮快速冷冻样本,于–20 ℃冷冻保存,随后带回实验室−80 ℃保存。采集背部中后段肌肉 (每尾在相同位置采集20 g肌肉) 用于检测肌肉成分。各项指标由广东省质量监督食品检验站进行检测,各实验组均设置3个重复,每个重复含10尾样本 (背部肌肉捣碎)。
表 1 3个广东鲂地理种群环境信息和样本生物学信息Table 1. Basic environmental information, biological information of three M. terminalis populations指标
Index采样点 Sampling site 琼海 QH 阳江 YJ 肇庆 ZQ 环境指标 Environmental index 水温 Water temperature/℃ 30.0±0.2 29.2±0.4 28.6±0.3 盐度 Salinity 0.03±0.01 0.01±0.02 0.01±0.01 酸碱度 pH 7.7±0.3 7.9±0.3 7.8±0.3 溶解氧质量浓度 DO/(mg∙L−1) 6.8±0.2 6.7±0.2 7.0±0.2 生物学指标 Biological index 体长 Body length ($\overline { X}\pm { \rm {SD}} $)/mm 233±17.6 253±20.7 271±27.3 体质量 Body mass ($\overline { X}\pm { \rm {SD}} $)/g 354±19.4 424±33.1 521±30.1 1.2 肌肉营养成分和能量密度测定方法
样本水分测定采用GB 5009.3—2016;粗蛋白测定采用GB 5009.5—2016;脂肪测定采用GB 5009.4—2016;灰分测定采用GB 5009.4—2016;氨基酸测定采用GB T5009.124—2016;脂肪酸测定采用GB 5009.168—2016。采用电感耦合等离子体质谱仪 (7700 Series) 依据GB 5009—2016测定样本中钾 (K)、钙 (Ca)、钠 (Na)、磷 (P)、镁 (Mg)、锌 (Zn)、铁 (Fe)、铜 (Cu)、锰 (Mn) 等矿质元素含量。根据联合国粮农组织/世界卫生组织 (FAO/WHO) 提出和1991年中国预防医学科学院营养与食品卫生研究所提出的氨基酸评分模式,计算氨基酸评分 (Amino acid score, AAS)、化学评分 (Chemical score, CS) 和必需氨基酸指数 (Essential amino acid index, EAAI) [9-10]。Phillipson微量能量仪 (Gentry Instruments Inc., Aiken, South Carolina, USA) 测定能量密度。
1.3 数据分析
采用单因素方差分析 (One-way ANOVA) 检验不同广东鲂地理种群肌肉营养成分差异显著性。如差异显著,则采用多重比较方法比较平均数之间的差异,显著性水平为0.05。数据分析采用SPSS 19.0统计软件进行。实验数据均用“平均值±标准差 (
$\overline X \pm {\rm{SD}} $ )”表示。采用R (3.1.14) 对3个广东鲂种群肌肉生化分析结果进行主成分分析 (Principal component analysis, PCA)。2. 结果
2.1 一般营养成分和能量密度分析
万泉河广东鲂肌肉水分质量分数显著高于西江种群,而粗蛋白质质量分数则显著低于西江种群 (P<0.05,表2)。粗脂肪和灰分质量分数在3个地理种群中均无显著性差异。西江种群肌肉能量密度显著高于万泉河和漠阳江种群 (P<0.05)。
表 2 3个广东鲂地理种群肌肉中的一般营养成分和能量密度Table 2. Nutritional composition of muscle of three M. terminalis populations项目
Item广东鲂种群 M. terminalis population 琼海 QH 阳江 YJ 肇庆 ZQ 水分质量分数 Moisture mass fraction/% 79.0±0.5a 78.2±0.9ab 77.0±0.6b 粗蛋白质质量分数 Crude protein mass fraction/% 18.3±0.5b 19.1±0.7ab 20.2±0.5a 粗脂肪质量分数 Crude lipid mass fraction/% 1.2±0.1 1.1±0.1 1.3±0.1 粗灰分质量分数 Ash mass fraction/% 1.1±0.1 1.2±0.1 1.2±0.1 能量密度 Energy density/(kJ·g−1) 3.1±0.1b 3.2±0.2b 3.6±0.2a 注:同行不同上标字母表示差异显著 (P<0.05),下表同此。 Note: Different superscript letters within the same row indicate significant difference (P<0.05). The same case in the following tables. 2.2 氨基酸组成分析与评价
3个广东鲂地理种群共检测出18种常见氨基酸 (表3)。西江种群肌肉中的总氨基酸含量 (Total amino acids, TAA) 最高,漠阳江种群次之,万泉河种群最低。在必需氨基酸 (Essential aamino acid, EAA) 中,西江种群的赖氨酸和亮氨酸含量显著高于万泉河种群 (P<0.05),漠阳江种群介于两者之间。呈味氨基酸中,西江种群肌肉中的天冬氨酸、谷氨酸、甘氨酸和丙氨酸含量均显著高于万泉河种群 (P<0.05)。漠阳江种群肌肉中必需氨基酸/总氨基酸 (EAA/TAA) 最高,而西江种群最低。呈味氨基酸/总氨基酸 (DAA/TAA) 在3个广东鲂地理种群肌肉中无明显差异。芳香氨基酸/支链氨基酸 (BCAA/AAA) 在万泉河种群肌肉中最高,漠阳江种群次之,西江种群最低。将3个广东鲂地理种群肌肉的EAAI进行标准模式 (FAO/WHO) 及全鸡蛋蛋白质模式2种评价 (表3),分别计算出各EAA的AAS、CS和EAAI (表4)。万泉河、漠阳江和西江种群肌肉中的第一限制性氨基酸为蛋氨酸+半胱氨酸,第二限制性氨基酸为缬氨酸,其余各EAA的AAS均高于1;各EAA的CS与AAS结果保持一致。3个广东鲂地理种群肌肉中的EAAI达80以上,说明其氨基酸组成十分均衡。其中西江种群肌肉EAAI最高 (85.05)。
表 3 3个广东鲂地理种群肌肉氨基酸组成Table 3. Comparison of amino acid composition of muscles of three M. terminalis populations项目
Item广东鲂种群 M. terminalis population 琼海 QH 阳江 YJ 肇庆 ZQ 天冬氨酸* Asp 1.80±0.04b 1.82±0.07b 1.99±0.05a 苏氨酸# Thr 0.80±0.02 0.80±0.03 0.86±0.04 丝氨酸 Ser 0.72±0.02 0.73±0.03 0.80±0.04 谷氨酸* Glu 2.78±0.06b 2.68±0.10b 2.99±0.14a 脯氨酸 Pro 0.61±0.02 0.63±0.02 0.66±0.04 甘氨酸* Gly 0.83±0.04b 0.87±0.04ab 0.97±0.06a 丙氨酸* Ala 1.06±0.02b 1.09±0.03b 1.18±0.08a 缬氨酸#△ Val 0.85±0.02 0.87±0.03 0.92±0.04 蛋氨酸 Met 0.53±0.01 0.53±0.02 0.57±0.03 异亮氨酸#△ Ile 0.78±0.02 0.79±0.02 0.83±0.03 亮氨酸#△ Leu 1.41±0.03b 1.41±0.05ab 1.52±0.05a 半胱氨酸 Cys 0.11±0.07 0.19±0.03 0.43±0.15 酪氨酸◆ Tyr 0.62±0.02 0.61±0.02 0.66±0.02 苯丙氨酸# Phe 0.72±0.02 0.74±0.02 0.80±0.03 赖氨酸# Lys 1.73±0.05a 1.73±0.09ab 1.87±0.03b 组氨酸○ His 0.42±0.01 0.45±0.04 0.48±0.05 精氨酸○ Arg 1.07±0.04 1.07±0.04 1.17±0.07 色氨酸# Trp 0.27±0.04 0.35±0.03 0.37±0.05 氨基酸总量 TAA 17.11±0.58a 17.37±0.69a 19.07±0.57b 呈味氨基酸总量 DAA 6.47±0.06a 6.46±0.08a 7.13±0.07b 必需氨基酸/非必需氨基酸 EAA/NEAA/% 72.41 73.03 69.95 必需氨基酸/总氨基酸 EAA/TAA/% 38.34 38.51 37.60 半必需氨基酸/总氨基酸 SEAA/TAA/% 8.36 8.41 8.26 芳香氨基酸/支链氨基酸 BCAA/AAA/% 4.90 5.03 4.05 注:#. 必需氨基酸;○. 半必需氨基酸;*. 呈味氨基酸;△. 支链氨基酸;◆. 芳香氨基酸。 Note: #. Essential amino acid; ○. Semiessential amino acid; *. Delicious amino acid; △. Branched chain amino acid; ◆. Aromatic amino acid. 表 4 3个广东鲂地理种群肌肉氨基酸评价Table 4. Evaluation of essential amino acids composition of muscle of three M. terminalis populations评价模式
Evaluation method氨基酸
Amino acids广东鲂种群 M. terminalis population 琼海 QH 阳江 YJ 肇庆 ZQ 氨基酸评分 AAS 苏氨酸 Thr 1.09 1.05 1.07 缬氨酸 Val 0.94 0.92 0.92 色氨酸 Trp 1.53 1.91 1.91 异亮氨酸 Ile 1.06 1.03 1.03 亮氨酸 Leu 1.09 1.05 1.07 赖氨酸 Lys 1.73 1.66 1.70 苯丙氨酸 Phe+酪氨酸 Tyr 1.20 1.17 1.19 蛋氨酸 Met+半胱氨酸 Cys 0.82 0.78 0.80 化学评分 CS 苏氨酸 Thr 0.93 0.90 0.91 缬氨酸 Val 0.71 0.69 0.69 色氨酸 Trp 0.96 1.24 1.31 异亮氨酸 Ile 0.80 0.78 0.78 亮氨酸 Leu 0.90 0.86 0.88 赖氨酸 Lys 1.34 1.28 1.31 苯丙氨酸 Phe+酪氨酸 Tyr 0.81 0.78 0.80 蛋氨酸 Met+半胱氨酸 Cys 0.47 0.45 0.46 必需氨基酸指数 EAAI 83.31 83.37 85.05 2.3 脂肪酸组成分析
3个广东鲂地理种群肌肉中共检测出23种常见脂肪酸 (表5),其中包括7 种饱和脂肪酸 (Saturated fatty acid, SFA) 7种单不饱和脂肪酸 (Monounsaturated fatty acid, MUFA) 和9种多不饱和脂肪酸 (Polyunsaturated fatty acids, PUFA)。SFA中C14:0、C16:0和C22:0在万泉河种群肌肉中的含量显著高于漠阳江和西江种群 (P<0.05)。万泉河种群肌肉中MUFA总量显著低于西江和漠阳江种群 (P<0.05)。其中,C16:1、C18:1 n-9t、C18:1 n-9c漠阳江种群肌肉中含量最高,而在万泉河种群肌肉中含量最低。C22:1 n-9、C24:1在漠阳江种群肌肉中含量显著低于西江和万泉河种群。西江种群肌肉中二十二碳六希酸 (DHA) 含量最高,显著高于万泉河和漠阳江种群 (P<0.05)。PUFA在万泉河种群肌肉中含量最高,西江种群次之,漠阳江种群最低。
表 5 3个广东鲂地理种群肌肉脂肪酸组成Table 5. Comparison of fatty acids of muscles of three M. terminalis populations% 项目
Item广东鲂种群 M. terminalis population 琼海 QH 阳江 YJ 肇庆 ZQ 肉豆蔻酸 C14:0 5.36±0.85a 3.53±0.41b 3.40±0.32b 十五碳酸 C15:0 1.02±0.25 0.66±0.05 0.70±0.13 棕榈酸 C16:0 27.33±1.47 a 21.83±1.16 b 21.80±1.99 b 珠光脂酸 C17:0 2.03±0.35 0.99±0.18 1.13±0.33 硬脂酸 C18:0 11.34±1.79a 5.97±1.03b 8.14±1.65ab 花生酸 C20:0 0.29±0.05a 0.15±0.02b 0.21±0.07ab 花生酸 C22:0 0.30±0.08a 0.10±0.01c 0.17±0.01b ∑饱和脂肪酸 SFA 47.67±2.12a 33.23±2.85b 35.56±4.35b 肉豆蔻烯酸 C14:1 0.06±0.02 0.12±0.02 0.08±0.01 棕榈油酸 C16:1 6.21±1.22b 10.45±1.73a 8.10±0.52ab 顺-11-二十碳一烯酸 C20:1 2.32±0.36 1.85±0.08 2.06±0.15 顺-15-二十四碳一烯酸 C24:1 0.27±0.09a 0.06±0.01b 0.24±0.10a 反式油酸 C18:1 n-9t 0.21±0.05b 0.43±0.07a 0.35±0.07ab 油酸 C18:1 n-9c 16.73±3.87b 35.53±4.35a 31.80±3.65a 二十二碳一烯酸 C22:1 n-9 1.09±0.36a 0.11±0.03b 1.03±0.61a ∑单不饱和脂肪酸 MUFA 26.89±3.93a 48.54±5.20b 43.66±7.08b 亚油酸 C18:2 n-6c 4.30±0.64a 2.23±0.33b 1.59±0.64b α-亚麻酸 C18:3 n-3 5.54±0.86a 4.03±1.56ab 2.35±0.51b γ-亚麻酸 C18:3 n-6 0.14±0.02a 0.08±0.02ab 0.01±0.00b 顺,顺-11,14-二十碳二烯酸 C20:2 0.39±0.08 0.34±0.01 0.36±0.04 顺-11,14,17-二十碳三烯酸 C20:3 n-3 0.32±0.05 0.27±0.06 0.21±0.08 顺,顺,顺-8,11,14-二十碳三烯酸 C20:3 n-6 0.39±0.09a 0.22±0.03b 0.14±0.01c 花生四烯酸 C20:4 n-6 (ARA) 5.27±0.89a 2.94±0.42b 4.88±0.95a 二十碳五烯酸 C20:5 n-3 (EPA) 3.16±0.72 4.06±0.10 4.09±0.87 二十二碳六烯酸 C22:6 n-3 (DHA) 5.74±1.03b 3.95±0.13c 7.03±1.28a ∑多不饱和脂肪酸 PUFA 25.11±2.83a 18.04±2.47b 20.64±5.61ab 2.4 矿质元素组成分析
3个广东鲂地理种群肌肉中均含有丰富的矿质元素,其中K质量分数最高,Ca次之 (表6)。西江种群肌肉中K和Ca质量分数显著高于万泉河种群,而Na和Mg质量分数则显著低于万泉河种群 (P<0.05)。万泉河种群肌肉Zn质量分数显著高于漠阳江和西江种群,而Mn和Fe质量分数显著低于漠阳江和西江种群 (P<0.05)。
表 6 3个广东鲂地理种群肌肉矿质元素组成Table 6. Mineral element of muscle of three M. terminalis populationsmg∙kg−1 元素
Element广东鲂种群 M. terminalis population 琼海 QH 阳江 YJ 肇庆 ZQ 钾 K 3 340.05±105.36b 3 460.04±192.92ab 3 820.36±221.12a 钙 Ca 1 050.12±28.87 b 1 100.25±40.02 ab 1 200.11±34.64 a 钠 Na 487.34±58.96a 345.35±7.23b 385.57±30.66b 镁 Mg 298.65±3.79 303.05±2.65 332.31±5.51 磷 P 241.59±25.97 230.45±2.08 247.78±4.04 锌 Zn 6.47±0.15a 4.16±0.18b 3.82±0.17b 铁 Fe 3.06±0.13c 4.75±0.14b 6.43±0.18a 铜 Cu 0.12±0.00 0.12±0.01 0.13±0.01 锰 Mn 0.21±0.01c 0.38±0.02b 0.64±0.01a 2.5 3个广东鲂种群生化分析结果的主成分分析
综合3个广东鲂种群生化分析结果,并进行PCA。西江种群分布距均万泉河和漠阳江种群较远,万泉河种群和漠阳江种群相对较近。PCA共提取了2个主成分,对变异的累积贡献率为80.25%。其中主成分1的贡献率为50.75%,主成分2的为29.50% (图1)。
![]() 图 1 3个广东鲂种群肌肉生化成分主成分分析散点图Figure 1. Scattering diagram of principal components for biochemical analysis parameters of muscle of three M. terminalis populations
图 1 3个广东鲂种群肌肉生化成分主成分分析散点图Figure 1. Scattering diagram of principal components for biochemical analysis parameters of muscle of three M. terminalis populations3. 讨论
鱼类肌肉中蛋白质和脂肪含量是评价其营养价值的重要指标[11]。3 个广东鲂地理种群肌肉的粗蛋白质量分数 (18.3%~20.2%) 高于团头鲂 (Megalobrama amblycephala)、鲤 (Cyprinus carpio)、鲢 (Hypophthalmichthys molitrix)、鳙 (H. nobilis) 和草鱼 (Ctenopharyngodon idella),与翘嘴鲌 (Culter alburnus) 接近[12-13]。3个广东鲂地理种群肌肉粗脂肪质量分数 (1.1%~1.3%) 较团头鲂、翘嘴鲌、鲢、鳙、斑鳜 (Siniperca scherzeri) 等低[12-14],与常见的海水鱼类如牙鲆 (Paralichthys olivaceus)[15]、黄斑篮子鱼 (Siganus oramin)[16]和日本鳗鲡 (Anguilla japonica)[17]类似,表现出典型的低脂肪、高蛋白的特点。本研究发现,3个广东鲂种群肌肉生化PCA结果显示西江种群分布距万泉河和漠阳江种群较远,可能是由于栖息地环境因子以及饵料生物种类存在明显差异。本研究还发现,西江种群能量密度显著高于漠阳江和万泉河种群。能量密度被认为是衡量鱼体能量储备水平的重要指标,能直接反映鱼类发育状况以及对外界环境因子的适应性[18]。鱼类生殖洄游是主动的、定期定向的高耗能运动,且鱼体自身能量储备有限,因此鱼类洄游须尽可能地调节自身身体结构、能量储备和代谢能力以适应生殖洄游的需要[19-20]。3个广东鲂种群生殖洄游距离存在明显差异,可能是导致种群间肌肉中能源物质的积累程度不同的主要原因之一。有研究发现鱼类肌肉能量累积和消耗与其洄游能力密切相关[21-22]。
鱼类肌肉中蛋白质的营养价值由各种EAA含量和组成比例决定[23-24]。本研究显示,在3个广东鲂种群肌肉中谷氨酸含量均最高,谷氨酸作为一种重要呈味氨基酸,具有促进脑发育、治疗神经系统疾病等作用[25]。3个广东鲂种群肌肉中谷氨酸含量均高于团头鲂与翘嘴鲌[12]。西江种群肌肉中谷氨酸含量显著高于漠阳江和万泉河种群,表明西江种群肌肉较万泉河和漠阳江种群风味更佳。3个广东鲂种群肌肉中赖氨酸含量均较高,其中,西江种群肌肉中赖氨酸含量最高。赖氨酸是人体EAA之一,不仅具有提高蛋白质利用率和促进人体生长发育的作用,还可以增强免疫力、改善神经系统、预防骨质疏松[24,26]。在FAO/WHO提出的人体均衡蛋白需求理想模式中,EAA/NEAA>60%的蛋白质质量较好[27],3个野生广东鲂种群肌肉均属于良好的蛋白源。西江种群肌肉中的EAAI最高 (85.05),说明其肌肉中EAA组成最为平衡,蛋白质营养价值最高。肌肉中的脂肪酸含量是影响肌肉风味的重要因素之一[28]。本研究发现,西江和漠阳江种群肌肉均表现出MUFA的高占比。有研究表明,MUFA在调节人体脂质代谢方面具有重要的生理作用[29]。PUFA中DHA与EPA含量是评价鱼类营养成分的关键指标[30]。西江种群肌肉中的DHA含量显著高于漠阳江和万泉河种群,表明西江种群肌肉的脂肪质量较高。
矿物质元素是构成人体组织的重要成分,参与人体内多种物质的代谢和生理活动[31]。3个广东鲂种群肌肉中Na、K、Ca等常规矿质元素以及Fe、Zn、Cu、Mn等微量元素均有检出。K、Fe、Zn等矿质元素含量低于异齿裂腹鱼 (Schizothoraxo connori)[32]。Ca含量显著高于褐点石斑鱼 (Epinephelus fuscoguttatus) 和青石斑鱼 (E. awoara) 等多种海鱼[31]。西江种群肌肉中Fe含量显著高于漠阳江和万泉河种群,而Zn含量则显著低于万泉河种群。Fe具有造血功能和促进人体生长的作用等,Zn可以促进儿童智力的正常发育[33]。3个广东鲂地理种群肌肉多种微量元素含量差异显著,这可能是由于栖息水环境的差异所致。万泉河种群相对西江种群,其主要栖息水域为河口,盐度相对较高,易受潮汐影响。有研究发现淡水环境中Fe含量均显著高于海水,Zn含量明显低于海水环境[34-35]。因此,栖息地环境差异导致了Zn在万泉河种群肌肉中富集度更高,Fe和Mn则在西江种群肌肉中富集度更高。
-
![]()
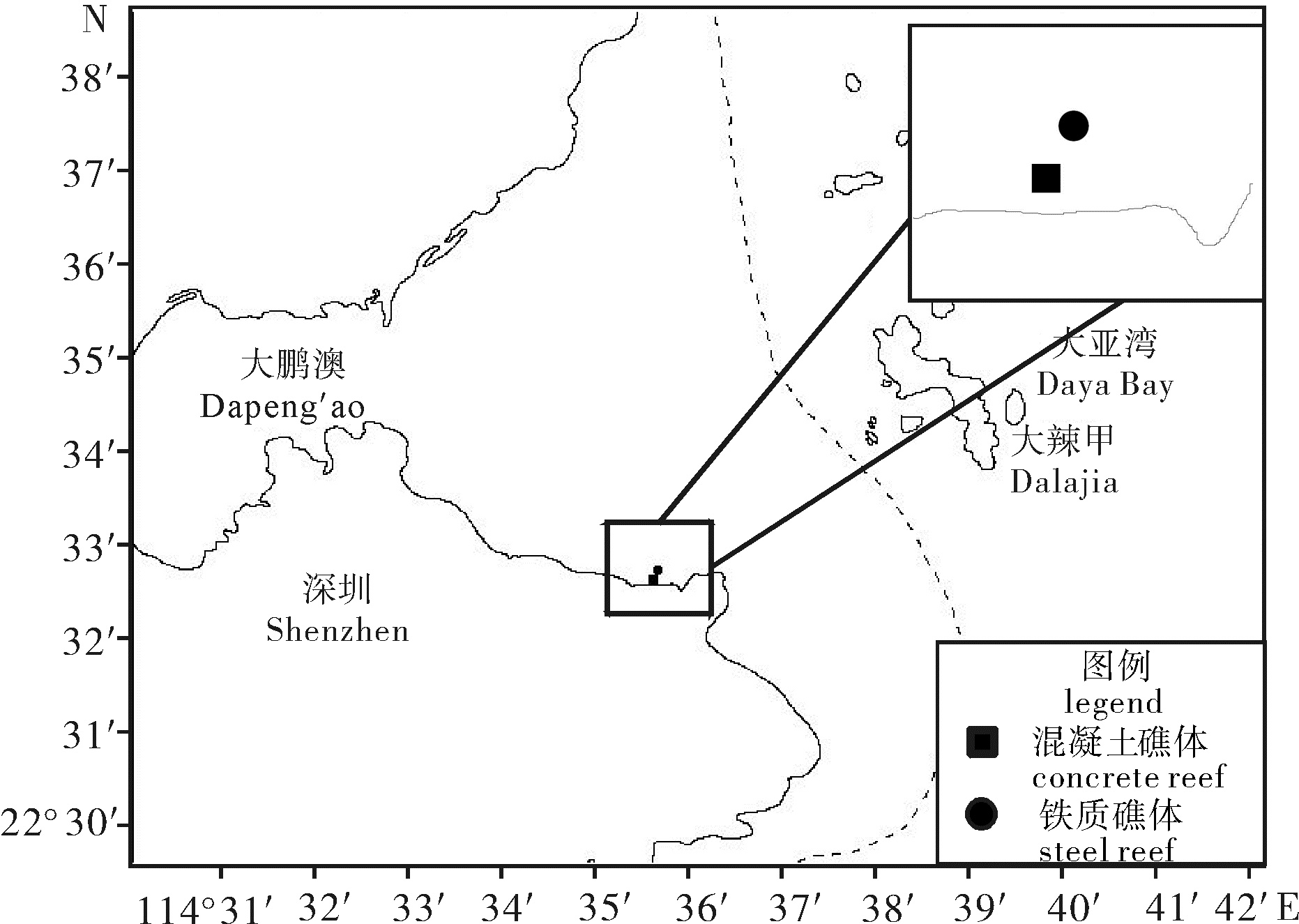
图 1 混凝土鱼礁和铁质鱼礁附着生物采样位置示意图
Figure 1. Sampling stations for biofouling communities on concrete and steel reefs
![]()
图 2 人工鱼礁混凝土礁体和铁质礁体附着生物群落多样性指数的季节变化
a.混凝土礁体;b.铁质礁体
Figure 2. Seasonal variation of diversity indexes of biofouling communities on concrete and steel reefs
a.concrete reef; b.steel reef
表 1 大亚湾混凝土鱼礁和铁质鱼礁上附着生物种类名录
Table 1 Species of fouling organisms on concrete and steel reefs, Daya Bay
种类
species混凝土礁体concrete reef 铁制礁体steel reef 春
spring夏
summer秋
autumn冬
winter春
spring夏
summer秋
autumn冬
winter海绵动物门 Spongla 岡田樽海绵 Sycon okadai + + 荔枝海绵 Tethya aurantium ++ 腔肠动物门 Coelenterata 水螅纲 Hydrozoa 筒状美螅 C.cylindrica + 总状真枝螅 Eudendrium racemosum ++ 双叉薮枝螅 Obelia dichotoma + 珊瑚纲 Anthozoa 石花虫 Telesto sp. + 海鸡头 Nephthea sp. + 侧花海葵 Anthopleura sp. + + ++ + + ++ + + 桂山厚丛柳珊瑚 Hicksonella guishanensis + 滑鞭柳珊瑚 Ellisella laevis + 刺鸡头 Dendronephthya sp. + + + 扁形动物门 Platyhelmintnes 外伪角涡虫 Pseudoceros exoptatus + + + + + + 柄涡虫Stylochus sp. + + + + 棘头动物门Acanthocephala 棘头虫Acanthocephalus + + + + + 环节动物门Annelida 多毛纲Polychaeta 华美盘管虫Semiserpula nanhaiensis ++ + + ++ ++ + 长须沙蚕N.loingior + + + + + + + 雾海鳞虫Halosydna nebulosa + + + ++ + 旋鳃虫Spirobranchus giganteus ++ + ++ ++ + ++ + ++ 须鳃虫C.tentaculata ++ ++ ++ ++ + + + 乳蛰虫Thelepus sp. ++ ++ + ++ ++ + 棒手拟隐鳞虫Hermadionella truncata ++ ++ + + + + 异足索沙蚕Lumbrineris heteropoda + + + ++ + + + 东海刺沙蚕N.donghaiensis + 珠须肌沙蚕Eunice antennata + + + 伪豆维虫Dorvillea pseudorubrovittata + + + + + + 麦缨虫Megalomma vesioulosum + 管缨虫Chone sp. ++ + + + + + 丝缨虫Hypsicomus phaeotaenia ++ + ++ + + ++ + + 多齿沙蚕N.multignatha ++ ++ ++ + ++ ++ ++ + 叶须虫P.laminosa ++ ++ + + ++ ++ + + 玛叶须虫P.malmgreni + + 渤海格鳞虫G.pohailnsis ++ ++ ++ ++ ++ ++ + ++ 亚洲哈鳞虫Harmothoe asiatica ++ ++ + ++ ++ + 背裂虫O.brunnea ++ ++ ++ + + ++ ++ + 尖锥虫Scoloplos armiger + 领襟松虫Lysidice collaris + 紫臭海蛹T.pupa + 软体动物门Mollusca 双神经纲Amphineura 网纹鬃毛石鳖Mopalia retifera + + + 腹足纲Gastropoda 史氏背尖贝Notoaomea schrencci + + + + + + 粗糙真蹄螺Euchelus scaber + + + ++ 梯螺Epitonium sp. + 黄口荔枝螺Thais luteostoma + + + + 蛎敌荔枝螺T.gradate + + + + 大沼螺Parafossarulus eximius + 三列笋螺Terebra triseriata + + 红带织纹螺Nassarius succinctus + 黑口滨螺Littorina malanostoma + 西格织纹螺N.siquinjorensis + 愚螺Amathina tricarinata + + + + + 节织纹螺Nassatius hepaticus + + + 珠带拟蟹守螺Cerithidea cingulata + + + ++ + ++ 美丽唇齿螺Engina pulchra + + + + + + 湖北钉螺Oncomelamia hupensis + + + + + 硬结原爱神螺Proterato callosa + + 疣荔枝螺T.clavigera + 杂色笔螺Mitra litterata + + + 丽核螺Pyrene bella ++ + + ++ ++ 瓣鳃纲Lamellibranhia 布纹蚶Barbation decussata + + 泥蚶Tegillarca granosa + + 双纹须蚶Barbatia bistrigata + + + + + + ++ 斑顶须蚶B.signata + + + + 翡翠贻贝P.viridis + + ++ ++ + ++ + 合浦珠母贝P.fucata martensi + 曲线索贻贝Hormomya mutabilis ++ ++ + 海月Placuma placenta + ++ + + + 翘鳞蛤Irus irus + + + + 变化短齿蛤Brachidontes variabilis ++ + ++ + 敦氏猿头蛤Chama dunkeri lischke + ++ ++ ++ + ++ ++ ++ 翘鳞猿头蛤C.lazarus linnarus + 草莓海菊蛤Sprnolylus fragum + 纹斑棱蛤T.liratum + + ++ + + + ++ + 斜纹心蛤Cardita leana + 铃蛤Jouannetia cumingi + 青蛤Cyclina sinensis + 文蛤M.meretrix + + + + 毛偏顶蛤Modiolus barbatus ++ ++ + + ++ + 角偏顶蛤M.metcalfei + 短偏顶蛤M.flavidus + 偏顶蛤M.elongatus + 方形蚶蛤Isognomon nucleus + + + + 细肋肌蛤Musculus mirandus ++ ++ + + + + + + 栉孔扇贝Chlamys sp. + + 华贵栉孔扇贝C.nobilis + 贻贝Mytilus edulis + + 光石蛏Lithophaga teres + + ++ + + + ++ 近江牡蛎O.rivularis + + 褶牡蛎O.plicatula + + + + + + + + 棘刺牡蛎O.echinata + + + + + + 马特海笋Martesia striata + + + + 宽壳全海笋B.dilatata + 嫁䗩 Cellana toreuma + 曲线索贻贝H.mutabilis + 西施舌Coelomaotra antiquata + 节肢动物门Arthropoda 蔓足亚纲Cirripedia +++ +++ 三角藤壶B.trigonus + +++ +++ +++ + +++ +++ +++ 红巨藤壶M.rosa ++ ++ ++ 端足类Amphipoda 中华蜾赢蜚Corophium sinense + + + 强壮板钩虾Stenothoe sp. +++ ++ +++ +++ ++ ++ 日本片钩虾Elasmopus japonicus ++ 尖额麦杆虫C.penantis + 大螯蜚Jassa sp. ++ ++ ++ ++ + 十足类Decapoda 鼓虾Alpheus sp. + + 东海铠甲虾Galathea balssi ++ + ++ + 锯额瓷蟹P.serratifrons ++ + + + ++ + + 光辉圆扇蟹Sphaeroxius nitidus + 双角互敬蟹Hyastenus diacanthus + 棘皮动物门Echinodermata 真蛇尾Ophiura sp. ++ ++ ++ ++ ++ ++ + ++ 近辐蛇尾Ophiactis affinis ++ 细雕刻肋海胆Temnotliurus toreumaticus ++ + + + ++ + + 海齿花Comanthus + 苔藓虫Bryozoa 宽松苔虫Caberea lata + + 多室草苔虫Bugula neritina + 被囊动物Ascidiacea 皱瘤海鞘S.plicata + + + ++ 冠瘤海鞘Styela canopus ++ ++ ++ + ++ ++ 乳突皮海鞘M.manhattensis ++ ++ ++ + ++ ++ ++ + 悉尼海鞘A.sydneiensis 注:+.<100个· m-2;+ +.<1 000个· m-2;+ + +.<10 000个· m-2
Note: +.<100 ind · m-2;+ +.<1 000 ind · m-2;+ + +.<10 000 ind · m-2 下载: 导出CSV
下载: 导出CSV
表 2 大亚湾混凝土鱼礁和铁质鱼礁上附着生物群落生物量
Table 2 Biomass of biofouling communities on concrete and steel reefs in Daya Bay
g·m-2 种类
species混凝土礁体concrete reef 铁制礁体steel reef 春
spring夏
summer秋
autumn冬
winter春
spring夏
summer秋
autumn冬
winter三角藤壶B.trigonus 12 027.33 5 257.67 7 983.67 6 726.00 20 503.33 6 990.00 4 261.67 175 092.67 褶牡蛎O.plicatula 12 329.00 9 487.33 13 389.33 21 132.33 4.67 17 610.67 1 288.67 4 496.67 翡翠贻贝P.viridis 3 448.00 1 439.33 8 463.00 9 125.33 10.35 4 373.33 13 541.33 2 094.00
下载: 导出CSV
表 3 大亚湾混凝土鱼礁和铁质鱼礁上附着生物群落优势种及其优势度
Table 3 Dominant species and dominance of biofouling communities on concrete and steel reefs in Daya Bay
优势种
dominant species优势度(Y) dominance 混凝土礁体concrete reef 铁制礁体steel reef 春
spring夏
summer秋
autumn冬
winter春
spring夏
summer秋
autumn冬
winter三角藤壶B.trigonus 0.198 3 0.118 4 0.134 4 0.106 1 0.289 9 0.152 6 0.081 9 0.080 5 真蛇尾Ophiura sp. 0.006 8 0.006 8 0.011 1 0.006 8 0.007 4 强壮板钩虾Stenothoe sp. 0.019 3 0.008 5 0.013 9 0.041 2 乳突皮海鞘M.manhattensis 0.006 4 0.006 2 0.010 4 渤海格鳞虫G.pohailnsis 0.007 6 0.006 7 0.006 6 叶须虫P.laminosa 0.006 6 0.008 1 须鳃虫C.tentaculata 0.006 2 0.010 3 背裂虫O.brunnea 0.008 2 0.010 8 翡翠贻贝P.viridis 0.006 6 锯额瓷蟹P.serratifrons 0.006 1 纹斑棱蛤T.liratum 0.008 7 红巨藤壶M.rosa 0.007 4 乳蛰虫Thelepus sp. 0.007 2 冠瘤海鞘S.canopus 0.007 6
下载: 导出CSV
-
[1] 黄宗国, 蔡如星. 厦门港附着生物生态的研究[J]. 厦门大学学报, 1961, 8(3): 221-250. https://xueshu.baidu.com/usercenter/paper/show?paperid=ae47398fdf77d072a2915af1b5f5367a&site=xueshu_se&hitarticle=1 [2] SMITH R I. Physiological and ecological problems of brackish waters[J]. Annu Biol Colloq, 1959, 20: 59-69. doi: 10.1007/BF02254908
[3] VERNBERG F J. Physiological ecology of estuarine organisms[M]. Columbia: University of South Carolina Press, 1975: 397. doi: 10.1007/978-94-017-8801-4_377
[4] ANNAPOLIS M. Woods hole oceanographic institute, marine fouling and its prevention[M]. Annapolis: United States Naval Institute, 1952: 388. https://xueshu.baidu.com/usercenter/paper/show?paperid=f61d02538da0bdda7c314c803bc49c10&site=xueshu_se
[5] 林传旭, 邹晓理, 王华接, 等. 广东碣石湾污损生物生态研究[J]. 海洋通报, 2011, 30(4): 287-293. https://xueshu.baidu.com/usercenter/paper/show?paperid=2188b56f6374ca6fe1644accd6fd4a62&site=xueshu_se [6] 严涛, 严文侠, 董钰, 等. 珠江口东南近海海区污损生物研究[J]. 海洋学报, 2003, 25(6): 117-125. doi: 10.3321/j.issn:0253-4193.2003.06.014 [7] 李勇, 洪洁漳, 李辉权. 珠江口竹洲人工鱼礁与相邻天然礁附着生物群落结构研究[J]. 南方水产科学, 2013, 2(9): 20-26. https://xueshu.baidu.com/usercenter/paper/show?paperid=e37bfa9a473c4781a6cabf789567a159&site=xueshu_se&hitarticle=1 [8] 黄宗国, 蔡如星. 海洋污损生物及其防除(上)[M]. 北京: 海洋出版社, 1984: 351. https://xueshu.baidu.com/usercenter/paper/show?paperid=2e5446e9b102f4725b115979224c100f&site=xueshu_se&hitarticle=1 [9] MEENAKUMARI B, NAIR N B. Ecology of fouling in Cochin Harbor[C]//6th International Congress on Marine Corrosion and Fouling: Marine Biology, 1984: 159-170. https://xueshu.baidu.com/usercenter/paper/show?paperid=264691701ee28fd5a007de52340bd443&site=xueshu_se
[10] SEMDES G W. Seasonal changes and fouling community interactions[M]//Marine biodeterioration and interdisciplinary study. Annapolis: United States Naval Institute, 1984: 155-160. https://www.semanticscholar.org/paper/Seasonal-Changes-and-Fouling-Community-Interactions-Smedes/0dce7de4b4c2b7b46d23ef6cda8bd823b749eb68
[11] 黄梓荣, 梁小芸, 曾嘉. 人工鱼礁材料生物附着效果的初步研究[J]. 南方水产, 2006, 2(1): 34-38. doi: 10.3969/j.issn.2095-0780.2006.01.007 [12] CALLOW M E. A world-wide survey of fouling on non-toxic and there anti-fouling[C]//6th International Congress on Marine Corrosion and Fouling: Marine Biology, 1984: 325-346.
[13] BOAVENTURA D, MOURA A, LEITAO F, et al. Macrobenthic colonization of artificial reefs on the southern coast of Portugal (Ancao, Algarve)[J]. Hydrobiologia, 2006, 555(1): 335-343. doi: 10.1007/s10750-005-1133-1
[14] HATCHER A M. Epibenthic colonization patterns on slabs of stabilized coal-waste in Pools Bay[J]. Hydrobiologia, 1998, 367: 153-162. doi: 10.1023/A:1003228100220
[15] MOURA A, BOAVENTURA D, CURDIA J, et al. Biomass production of early macrobenthic communities at the Faro/Ancao artificial reef (Portugal): effect of depth and reef layer[J]. Bull Mar Sci, 2006, 78(1): 83-92. https://xueshu.baidu.com/usercenter/paper/show?paperid=309305fa0e04891504c158beaa27d1d4&site=xueshu_se&hitarticle=1
[16] WALKER S J, SCHLACHER T A, SCHLACHER-HOENLINGER M A. Spatial heterogeneity of epibenthos on artificial reefs: fouling communities in the early stages of colonization on an East Australian shipwreck[J]. Mar Ecol, 2007, 28(4): 1-11. https://xueshu.baidu.com/usercenter/paper/show?paperid=17a90155db8a237cf0dcff38466fe6bc&site=xueshu_se&hitarticle=1
[17] SANDROCK S, SCHARF E M, MOHR T. The development of fouling communities on an artificial reef off Nienhagen[M]. 2011: 1-18.
[18] 方芳, 严涛. 南海污损生物研究的现状及展望[J]. 热带海洋学报, 2004, 23(1): 76-85. doi: 10.3969/j.issn.1009-5470.2004.01.011 [19] 李传燕, 黄宗国, 郑成兴, 等. 大亚湾人工鱼礁附着生物的初步研究[J]. 应用生态学报, 1991, 2(1): 63-69. doi: 10.3321/j.issn:1001-9332.1991.01.013 [20] 郑成兴, 黄宗国, 李传燕, 等. 大亚湾珍珠养殖场的污损生物[J]. 应用生态学报, 1991, 2(2): 146-152. doi: 10.3321/j.issn:1001-9332.1991.02.012 [21] 严颂凯, 黄宗国. 大亚湾码头桩柱的污损生物群落[J]. 海洋学报, 1992, 14(3): 115-120. https://xueshu.baidu.com/usercenter/paper/show?paperid=c0fc12d8b4e8d0e8cc5a167ac7bbbbb6&site=xueshu_se&hitarticle=1 [22] 曾呈奎, 张竣甫. 黄海西部底栖藻类记录的评论[J]. 海洋科学集刊, 1964, 6: 1-25. http://qdhys.ijournal.cn/hykxjk/ch/reader/view_abstract.aspx?file_no=19640601&flag=1 [23] 曾呈奎, 张竣甫. 黄海和东海的海藻区系[J]. 海洋与湖沼, 1959, 2(1): 43-52. https://xueshu.baidu.com/usercenter/paper/show?paperid=2aa88bdd365b52da40b14b70be82c4d2&site=xueshu_se&hitarticle=1 -
期刊类型引用(10)
1. 梁梓龙,解志龙,姜晓东,陈文彬,罗明,吴旭干. 中华绒螯蟹“长荡湖1号”选育第二代扣蟹阶段养殖性能研究. 水产科技情报. 2024(01): 8-14 .  百度学术
百度学术
2. 解志龙,姜晓东,范陈伟,荆晶,成永旭,吴旭干. 不同规格中华绒螯蟹成蟹的形态特征和组织系数比较. 上海海洋大学学报. 2024(01): 67-76 . 百度学术
3. 李凤璐,朱筛成,李嘉尧,成永旭. 黄河口盐碱地区稻蟹种养模式下中华绒螯蟹适宜放养密度的初探. 上海海洋大学学报. 2024(01): 53-66 . 百度学术
4. 梁梓龙,姜晓东,张光宝,吴旭干. 超大规格中华绒螯蟹繁育子代生长性能及养殖效果. 中国水产科学. 2024(02): 197-208 . 百度学术
5. 张光宝,姜晓东,陈文彬,周文全,罗明,吴旭干. 水草种植模式对全雄中华绒螯蟹成蟹养殖性能和经济效益的影响. 南方水产科学. 2023(02): 107-115 . 本站查看
6. 张光宝,姜晓东,庄振俊,陈文彬,罗明,成永旭,陈晓武,吴旭干. 中华绒螯蟹“长荡湖1号”新品系选育第二代(G2)生长、性腺发育和养殖性能的研究. 海洋与湖沼. 2023(05): 1444-1453 . 百度学术
7. 彭涛,张冬冬,姜晓东,罗明,陈文彬,成永旭,吴旭干. 中华绒螯蟹“长荡湖1号”奇数年群体选育第二代的生长性能和养殖效果评估. 中国水产科学. 2023(10): 1177-1187 . 百度学术
8. 殷乐,姜晓东,王海宁,陈瑜,吴旭干. 不同品系中华绒螯蟹土池生态育苗效果比较. 水产科技情报. 2022(04): 193-199 . 百度学术
9. 张冬冬,范陈伟,姜晓东,陈瑜,吴旭干. 中华绒螯蟹白壳和绿壳品系生殖性能、胚胎色泽和生化组成比较. 南方水产科学. 2022(03): 102-110 . 本站查看
10. 庄振俊,张冬冬,姜晓东,陈文彬,陈晓武,成永旭,吴旭干. 中华绒螯蟹“长荡湖1号”奇数年子一代的成蟹养殖性能评估. 海洋渔业. 2022(06): 747-758 . 百度学术
其他类型引用(1)


计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 11
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
