
Statolith morphology of purpleback flying squid Sthenoeuthis oualaniensis in the central South China Sea
-
摘要:
根据2012年9月~10月于南海中部海域采集的鸢乌贼(Sthenoteuthis oualaniensis)样本90尾,对其耳石形态特征及计量学参数进行了初步研究。结果表明,耳石样本具有明显的背区、侧区、吻区和翼区结构,背区小,侧区和翼区宽大,吻区长窄。南海中部海域鸢乌贼按照耳石形态的差异可分为3个不同的群体。耳石总长(total statolith length,TSL)、翼区长(wing width,WL)、最大宽(maximum width,MW) 与胴长之间呈对数函数关系(P < 0.05),吻区长(rostrum length,RSL)、背侧区长(dorsolateral length,DLL)与胴长之间呈线性函数关系(P < 0.05)。随着胴长的增加,耳石绝对尺寸逐渐增大,相对尺寸逐渐减小,耳石的形态结构比例基本不变,即背侧区长、侧区长、吻区长、翼区长同耳石总长之比基本保持一致,分别为48.05%、60.05%、35.44%和67.59%,其中侧区长、翼区长与耳石总长的比值随胴长增大而增加的趋势较为明显。
Abstract:We studied the morphology, microstructure and demographic parameters of statolith of Sthenoeuthis oualaniensis from 90 individuals captured in the central South China Sea during September to October, 2012. The morphologic results show that the statolith had great lateral dome and wing dome, small dorsal dome and long narrow rostrum dome. S.oualaniensis in the central South China Sea could be devided into three populations according to their statolith morphology. The total statolith length (TSL), wing length (WL), maximum width (MW) were logarithmically related to the mental length (ML) (P < 0.05);while the rostrum length (RSL) and dorsolateral length (DLL) were linearly related to the ML(P < 0.05). The absolute size and relative size of statolith gradually increased and decreased with increasing ML, respectively. The ratios of DLL, LDL, RSL and WL to TSL almost remained the same, being 48.05%, 60.05%, 35.44% and 67.59%, respectively; and the ratios of LDL and WL to TSL were relatively higher.
-
鸢乌贼(Sthenoteuthis oualaniensis)隶属枪形目、柔鱼科、鸢乌贼属,广泛分布于印度洋、太平洋的热带和亚热带海域,印度洋西北部海域和南海分布数量较多[1-2]。南海鸢乌贼渔业的研究是中国维护南海主权突出存在的重要砝码,对加强中国南海鸢乌贼的研究具有特殊的政治意义[3]。中国台湾和东南亚渔业开发中心(SEAFDEC)1998年~1999年与1998年~2001年相继在南海开展鸢乌贼资源调查,分别估算南海鸢乌贼的生物量为150×104 t和113.2×104 t [4-5]。中国于2003年~2005年对印度洋西北海域的鸢乌贼资源进行调查研究[6-9],初步了解其资源量和生长繁殖等信息。然而,目前针对南海鸢乌贼种群生物学的研究较少[10-11],尤其是对南海鸢乌贼耳石形态特征的研究未见报道。
耳石是头足类信息的良好载体,其形态特征可作为种类鉴定特征,其生长轮纹可用以年龄估计、推算孵化日期和生长率等[12-19]。笔者根据2012年秋季采集于南海中部海域的鸢乌贼样本,观测分析耳石形态特征,寻找可以表征耳石形态特征的主成分,分析南海中部海域鸢乌贼可能存在的种群及其分布情况,探讨鸢乌贼耳石形态结构变化与个体生长的关系,获得各特征参数与胴长之间的最适模型,以期了解南海中部海域鸢乌贼种群结构与地理分布,为进一步研究南海鸢乌贼年龄与生长积累基础资料。
1. 材料与方法
1.1 材料
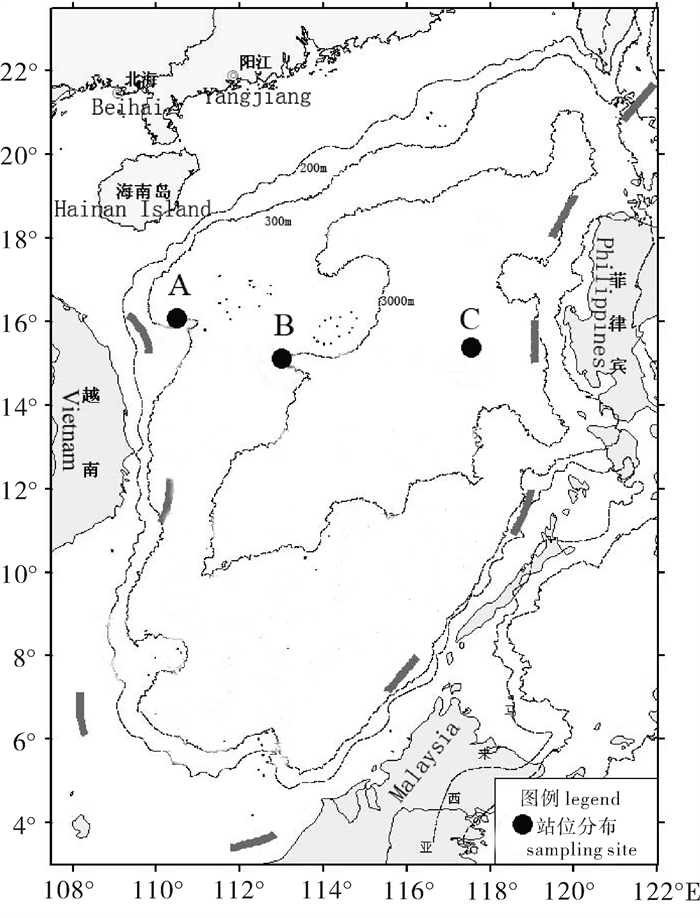
鸢乌贼样本采集于2012年9月~10月,采集海域为109°40′~117°31′E, 14°59′~16°20′N(图 1),采集方法为灯光罩网作业(主尺度为281.60 m×80.18 m,网口网目尺寸为35 mm,网囊最小网目尺寸为17 mm)。对南海中部海域3个站位样本进行研究。其中A站位(109°40′E,16°20′N)随机抽取样本30尾,B站位(112°29′E,14°53′N)随机抽取样本30尾,C站位(117°32′E,14°59′N)随机抽取样本30尾。样本冷冻保存运回实验室。此次检测鸢乌贼样本共90尾,胴长范围为78~193 mm,纯体质量范围为13.74~258.03 g。
![]() 图 1 南海中部海域鸢乌贼采集站位示意图Fig. 1 Sampling sites of S.oualaniensis in the central South China Sea
图 1 南海中部海域鸢乌贼采集站位示意图Fig. 1 Sampling sites of S.oualaniensis in the central South China Sea1.2 方法
1.2.1 耳石形态测量
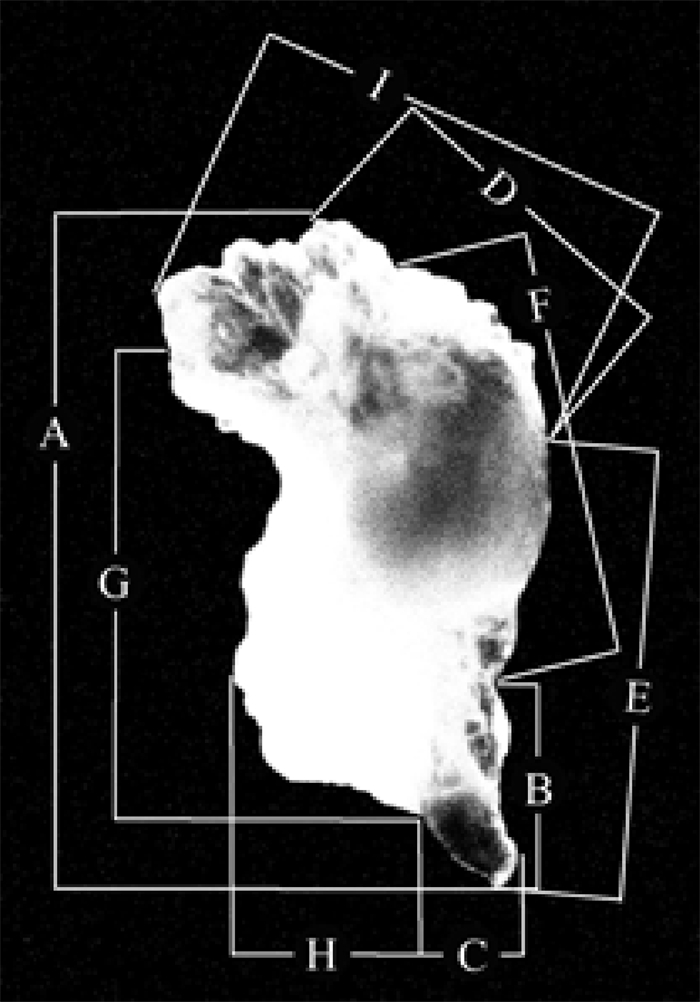
耳石置于Olympus生物显微镜(物镜×10、×40、×100,目镜×10)下拍照,分别测量耳石各形态参数值,分别包括耳石总长(total statolith length,TSL)、吻区长(rostrum length,RSL)、吻区宽(rostrum width,RSW)、背侧区长(dorsolateral length,DLL)、腹外侧长(ventrolteral length,VLL)、侧区长(lateral dome length,LDL)、翼区长(wing length,WL)、翼区宽(wing width,WW)和最大宽度(maximum width,MW)[20](图 2)。
![]() 图 2 鸢乌贼耳石形态参数示意图A.耳石总长;B.吻区长;C.吻区宽;D.背侧区长;E.腹外侧长;F.侧区长;G.翼区长;H.翼区宽;I.最大宽度Fig. 2 Morphometric measurement of S.oualaniensisA. total statolith length, TSL; B. rostrum length, RSL; C. rostrum width, RSW; D. dorsolateral length, DLL; E. ventrolteral length, VLL; F. lateral dome length, LDL; G. wing length, WL; H. wing width, WW;I. maximum width, MW
图 2 鸢乌贼耳石形态参数示意图A.耳石总长;B.吻区长;C.吻区宽;D.背侧区长;E.腹外侧长;F.侧区长;G.翼区长;H.翼区宽;I.最大宽度Fig. 2 Morphometric measurement of S.oualaniensisA. total statolith length, TSL; B. rostrum length, RSL; C. rostrum width, RSW; D. dorsolateral length, DLL; E. ventrolteral length, VLL; F. lateral dome length, LDL; G. wing length, WL; H. wing width, WW;I. maximum width, MW1.2.2 数据处理方法
利用SPSS 13.0统计软件对9个耳石形态参数进行主成分分析,分析耳石主要形态参数各比值(主要形态参数值与胴长之比)与胴长的关系;对耳石9个形态参数进行聚类分析,讨论样本的种群结构和分布情况;利用函数模型拟合耳石形态特征参数值与胴长的关系。
2. 结果
2.1 耳石形态及种群分布
2.1.1 耳石形态特征
鸢乌贼样本中各生物学特征参数值分别为:胴长78~193 mm;纯质量13.74~258.03 g。耳石样本具有明显的背区、侧区、吻区和翼区结构,背区小,侧区和翼区宽大,吻区长窄。耳石各形态参数值分别为TSL 616~1 243 μm;RSL 60~478 μm;MW 423~783 μm;LDL 478~731 μm;DLL 314~691 μm;VLL 334~894 μm;WL 366~940 μm;RSW 171~454 μm;WW 294~582 μm。
对上述9个形态参数进行主成分分析(表 1),结果显示,第一、第二、第三、第四和第五主成分解释形态参数的贡献率分别为64.36%、10.67%、8.71%、4.74%和4.46%,累计约为90%。第一主成分中除WL形态参数之外,各形态参数负载系数均在0.7以上;第二主成分中形态参数WL、RSL、DLL、LDL、VLL负载系数较大,均在0.3以上;第三主成分中形态参数WL、RSL、LDL负载系数较大,均在0.3以上;第四主成分中形态参数RSW、LDL负载系数较大,均在0.3以上;第五主成分中形态参数RSW、DLL负载系数较大,均在0.3以上。
表 1 鸢乌贼耳石9个形态参数5个主成分负荷值和贡献率Table 1 Loading and contribution rates of five principal components for nine morphometric measurement of statolith from S.oualaniensis形态参数
morphometric measurement主成分 principal component 1 2 3 4 5 耳石总长/μm TSL 0.897 0.192 -0.107 -0.278 -0.017 吻区长/μm RSL 0.778 0.412 -0.351 -0.059 -0.198 翼区长/μm WL 0.475 0.536 0.675 0.166 -0.017 最大宽度/μm MW 0.880 -0.133 0.088 0.054 -0.126 背侧区长/μm DLL 0.804 -0.364 0.038 0.176 -0.310 腹外侧长/μm VLL 0.849 0.328 -0.244 0.001 0.243 侧区长/μm LDL 0.752 -0.348 0.305 -0.331 0.271 吻区宽/μm RSW 0.810 -0.174 -0.167 0.412 0.311 翼区宽/μm WW 0.890 -0.239 0.060 -0.067 -0.140 贡献率/% contribution rate 64.36 10.67 8.71 4.74 4.46 2.1.2 种群分布
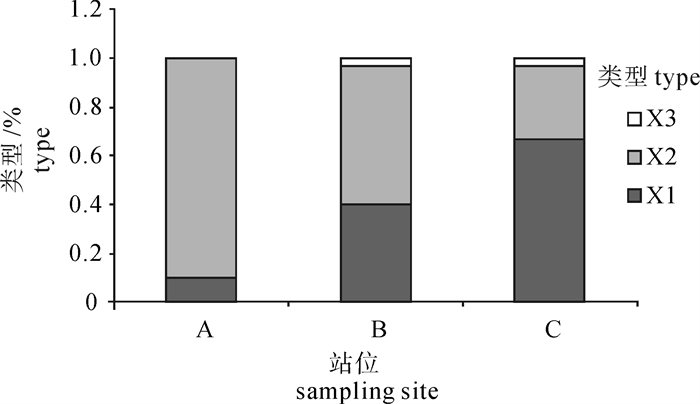
依据耳石的9个形态特征的研究,对南海中部海域鸢乌贼进行聚类分析,发现主要聚为3支。根据其所采集的站位分布,发现X1型鸢乌贼主要分布于C站位,随着经度的变化,自东向西逐渐减少;X2型鸢乌贼在A站位分布较高,随着经度的变化,自西向东逐渐减少;X3型鸢乌贼只在B和C站位零星分布(图 3)。
![]() 图 3 依据耳石形态结构的鸢乌贼种群在南海中部海域的分布Fig. 3 Distribution of S.oualaniensis populations by statolith morphology in the central South China Sea
图 3 依据耳石形态结构的鸢乌贼种群在南海中部海域的分布Fig. 3 Distribution of S.oualaniensis populations by statolith morphology in the central South China Sea2.2 耳石的生长
2.2.1 耳石绝对大小的变化
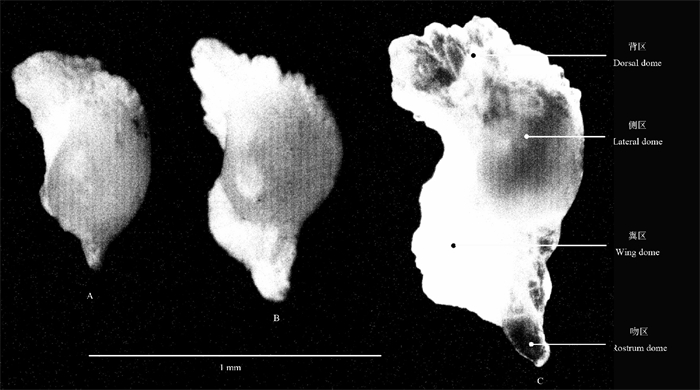
鸢乌贼个体胴长在78~100 mm范围的耳石,其结构简单,背区和侧区较大但分割不明显,吻区小,翼区不明显。随着个体的增大,当胴长达到100~193 mm范围时,其耳石各区不断增长;成体的耳石具有明显的背区、侧区、翼区和吻区,其中背区、侧区和翼区较大,吻区长窄(图 4)。
![]() 图 4 鸢乌贼耳石形态变化示意图A.耳石总长626 μm;B.耳石总长819 μm;C.耳石总长1 086 μmFig. 4 Morphological variation of statolith from S.oualaniensisA. TSL 626 μm; B. TSL 819 μm; C. TSL 1 086 μm
图 4 鸢乌贼耳石形态变化示意图A.耳石总长626 μm;B.耳石总长819 μm;C.耳石总长1 086 μmFig. 4 Morphological variation of statolith from S.oualaniensisA. TSL 626 μm; B. TSL 819 μm; C. TSL 1 086 μm2.2.2 耳石相对大小的变化
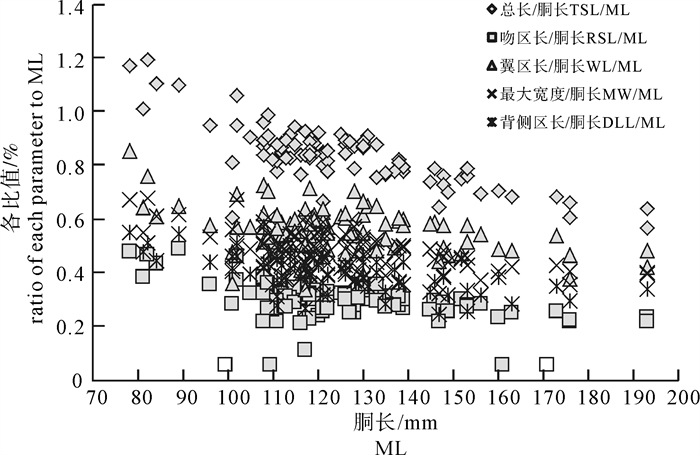
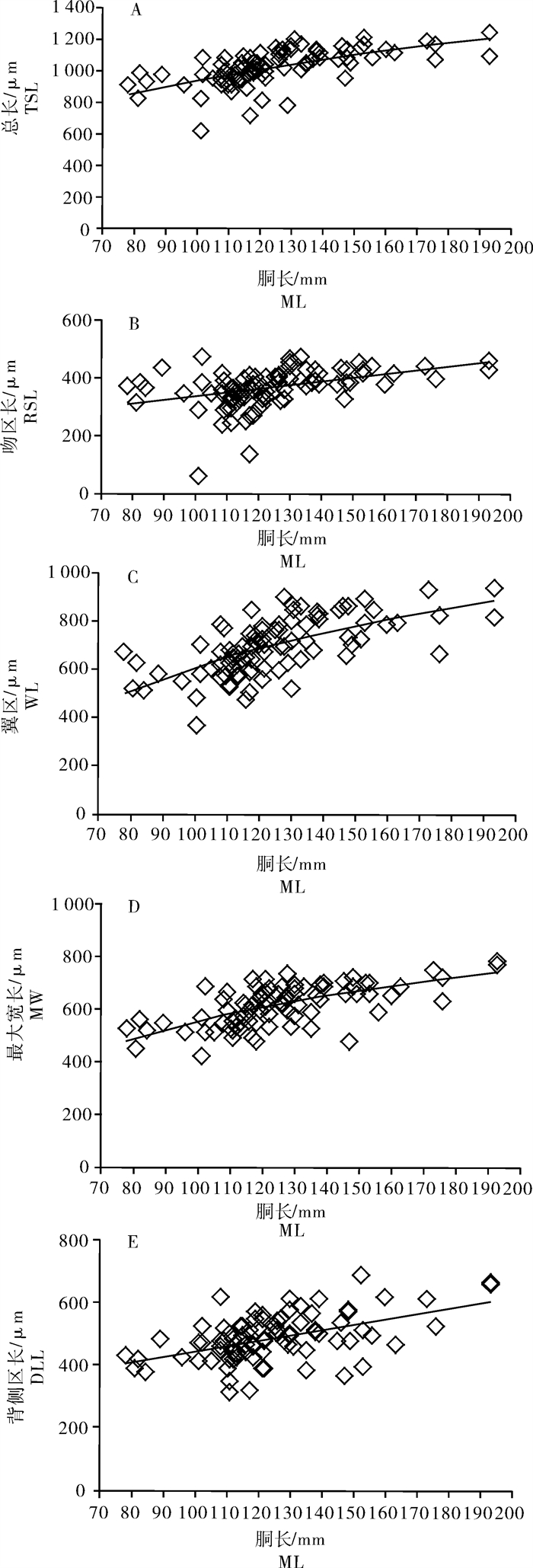
随着鸢乌贼个体胴长的不断增大,TSL/ML之比值的下降趋势最为明显,而RSL/ML、WL/ML、MW/ML、DLL/ML之比值的下降趋势略缓,变化趋势较为一致(图 5)。统计分析显示,鸢乌贼生长过程中其耳石的相对尺寸随个体胴长的增大而逐渐减小。TSL、RSL、WL、MW、DLL与胴长的比值分别由胴长78 mm时的1.172%、0.480%、0.861%、0.676%、0.553%减小为胴长193 mm时的0.568%、0.239%、0.487%、0.400%、0.341%。
![]() 图 5 鸢乌贼耳石总长/胴长、吻区长/胴长、翼区长/胴长、最大宽度/胴长、背侧区长/胴长的值与胴长关系Fig. 5 Relationship between ratios of TSL/ML, RSL/ML, WL/ML, MW/ML, DLL/ML and ML of S.oualaniensis
图 5 鸢乌贼耳石总长/胴长、吻区长/胴长、翼区长/胴长、最大宽度/胴长、背侧区长/胴长的值与胴长关系Fig. 5 Relationship between ratios of TSL/ML, RSL/ML, WL/ML, MW/ML, DLL/ML and ML of S.oualaniensis2.2.3 主要形态特征参数值与胴长的关系
综合性指标TSL、RSL、WL、MW、DLL与胴长之间分别以指数函数、线性函数、幂函数和对数函数进行模拟。结果显示,南海中部海域鸢乌贼耳石的TSL、WL、MW与胴长呈对数函数关系(P < 0.05);RSL、DLL与胴长呈线性函数关系(P < 0.05)。关系式分别为:
TSL=395.47ln(ML)-877.54 (R2=0.386 4,n=90) (图 6-A)
![]() 图 6 鸢乌贼耳石总长、吻区长、翼区长、最大宽度、背侧区长与胴长关系A.总长/胴长;B.吻长/胴长;C.翼长/胴长;D.最大宽/胴长;E.背侧区长/胴长Fig. 6 Relationship between TSL, RSL, WL, MW, DLL and ML of S.oualaniensisA.TSL/ML; B.RSL/ML; C.WL/ML; D.MW/ML; E.DLL/ML
图 6 鸢乌贼耳石总长、吻区长、翼区长、最大宽度、背侧区长与胴长关系A.总长/胴长;B.吻长/胴长;C.翼长/胴长;D.最大宽/胴长;E.背侧区长/胴长Fig. 6 Relationship between TSL, RSL, WL, MW, DLL and ML of S.oualaniensisA.TSL/ML; B.RSL/ML; C.WL/ML; D.MW/ML; E.DLL/MLRSL=1.282 4×ML+207.7 (R2=0.179 4,n=90) (图 6-B)
WL=434.82ln(ML)-1396.5(R2=0.416 4,n=90) (图 6-C)
MW=292.37ln(ML)-794.89 (R2=0.424 9,n=90) (图 6-D)
DLL=1.728 6×ML+274.1 (R2=0.270 4,n=90) (图 6-E)
式中TSL、RSL、WL、MW、DLL分别为耳石总长、吻区长、翼区长、最大宽度、背侧区长,单位为μm;ML为胴长,单位为mm。模拟结果中,吻区长与胴长关系图中(图 6-B),胴长分别为101 mm和117 mm的样本,其吻区长分别为59.99 μm和134.64 μm。这2个样本吻区长数据远小于其他样本的吻区长数据,经检查发现这2个样本的吻区较小,处于耳石形态生长早期(图 3-A)。
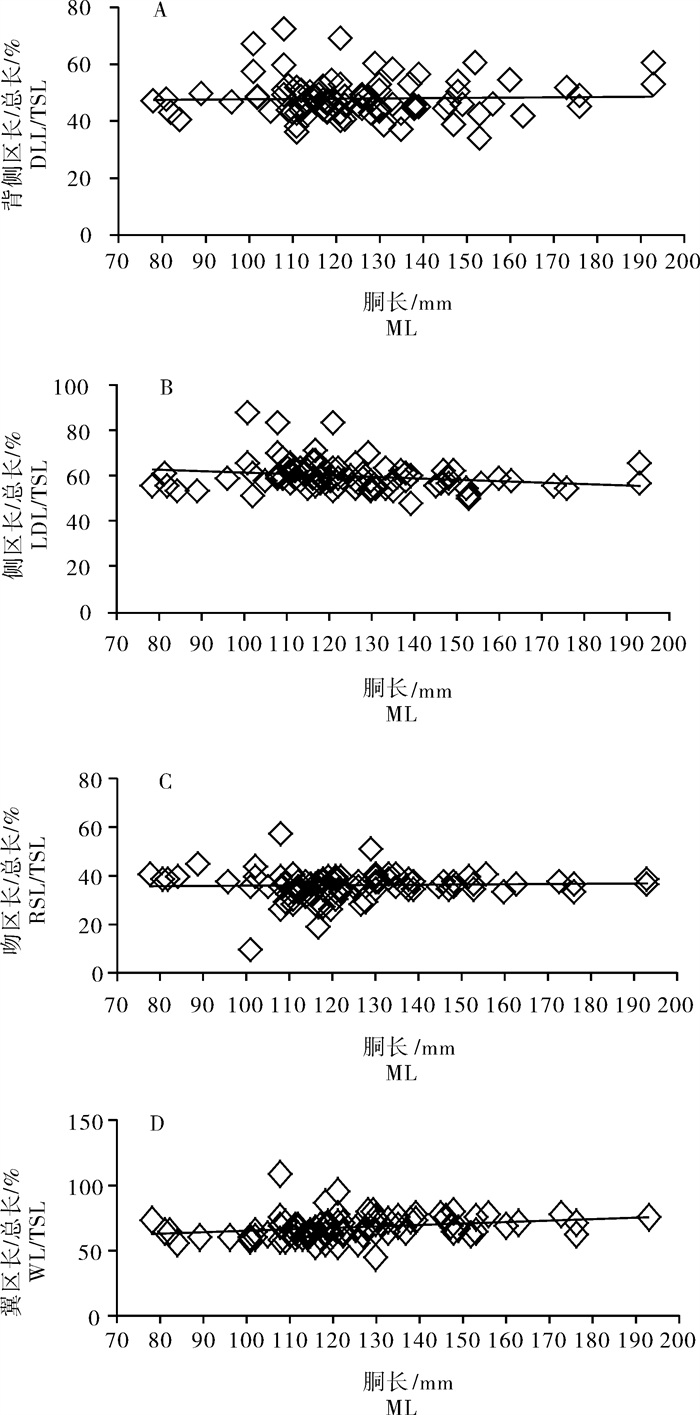
2.2.4 耳石各区形态参数相对长度与胴长的关系
由图 7-A~D可看出,耳石各区DLL、LDL、RSL、WL同TSL之间比值随胴长的增加而增加的趋势不是很明显,其平均值分别为48.05%、60.05%、35.44%和67.59%。其中,DLL/TSL、RSL/TSL值随胴长增加呈平缓趋势(δDLL/TSL:0.003 0;δRSL/TSL:0.003 4);LDL/TSL值随胴长增加呈下降趋势(δLDL/TSL:0.004 0);而WL/TSL值随胴长增加呈增长趋势(δWL/TSL:0.002 4)。
![]() 图 7 鸢乌贼耳石背侧区长/总长、侧区长/总长、吻区长/总长、翼区长/总长与胴长的关系A.(背侧区长/总长)/胴长;B.(侧区长/总长)/胴长;C.(吻区长/总长)/胴长;D.(翼区长/总长)/胴长Fig. 7 Relationship between DLL/TSL, LDL/TSL, RSL/TSL, WL/TSL and mantle length of S.oualaniensisA.(DLL/TSL)/ML; B.(LDL/TSL)/ML; C.(RSL/ TSL)/ML; D.(WL/TSL)/ML
图 7 鸢乌贼耳石背侧区长/总长、侧区长/总长、吻区长/总长、翼区长/总长与胴长的关系A.(背侧区长/总长)/胴长;B.(侧区长/总长)/胴长;C.(吻区长/总长)/胴长;D.(翼区长/总长)/胴长Fig. 7 Relationship between DLL/TSL, LDL/TSL, RSL/TSL, WL/TSL and mantle length of S.oualaniensisA.(DLL/TSL)/ML; B.(LDL/TSL)/ML; C.(RSL/ TSL)/ML; D.(WL/TSL)/ML3. 讨论
鸢乌贼耳石生长早期,其结构简单,呈水滴状,背区和侧区较大,吻区小,翼区尚未形成[11]。ARKHIPKIN等[21]利用耳石研究鸢乌贼发现,其稚鱼期结束年龄为100~110 d,此时ML约100 mm。随个体生长,其耳石各区日趋明显;成体的耳石具有明显的背区、侧区、翼区和吻区,其中背区、侧区和翼区较大,吻区长窄。耳石的形态特征能反映其所栖息水层的信息:生活于水体底层种类其耳石较大,吻区前端短平、翼区窄;生活于水体上层种类其耳石较小,吻区前端长窄、翼区宽[22-24]。鸢乌贼栖息于上层水体,印度洋西北海域鸢乌贼的耳石具有长窄的吻区和宽大的翼区[9],南海中部海域鸢乌贼的耳石亦具有长窄的吻区和宽大的翼区。南海中部海域和印度洋西北海域的鸢乌贼耳石外部形态特征的这种相似,说明鸢乌贼的耳石外部形态与其栖息的水层有关系。头足类耳石的生长,温度是最为主要的因素[2]。
NESIS[25]将鸢乌贼分为大型群、中型单轴群、中型双轴群、小型群、微型群,后3个种群出现在太平洋。刘必林[2]根据孵化期,将印度洋西北海域鸢乌贼分为春生群、夏生群和秋生群。根据耳石形态参数,可以将南海中部海域鸢乌贼分为3个不同的种群。X1型鸢乌贼自西向东逐渐增多,X2型鸢乌贼自西向东逐渐减少,X3型鸢乌贼零星分布;X3型鸢乌贼耳石具有不甚发达的侧区、背区、翼区,吻区短小,确认其为鸢乌贼耳石生长的早期阶段。而南海海域鸢乌贼种群及分布的情况,则有待进一步的研究。
南海海域和印度洋海域的鸢乌贼耳石研究结果相同之处是在鸢乌贼个体发育过程中,随着个体胴长的增长,其耳石的绝对尺寸逐渐增加,相对尺寸则逐渐减小;而两研究结果的不同之处是笔者研究中鸢乌贼耳石形态参数TSL、WL、MW与胴长之间呈对数函数关系,形态参数RSL、DLL与胴长之间呈线性函数关系,而印度洋西北海域鸢乌贼耳石主要形态参数TSL、RSL、WL、MW、DLL与胴长之间呈明显的幂函数关系[9],通过比较,推测造成这种不同结果的原因可能是印度洋西北海域鸢乌贼的采集方法为钓捕作业,难以钓获胴长小于150 mm个体[9];南海海域的鸢乌贼采集方法为灯光罩网作业,胴长范围为78~193 mm。
刘必林等[9]研究印度洋西北海域鸢乌贼耳石发现,相对于耳石总长的背侧区长、侧区长和翼区长分别维持42%、57%和75%。南海中部海域鸢乌贼耳石各区形态参数相对长度与胴长的关系说明,其耳石生长较为均匀,无明显的滞后或快速生长期,相对于耳石总长的背侧区长、侧区长和翼区长分别维持48%、60%和68%。2个海域鸢乌贼耳石生长均较为均匀,而影响耳石生长最主要的因素是温度,说明2个海域鸢乌贼生活时期温度均无急剧的变化;而两海域鸢乌贼耳石WL/TSL值均高于其他值,说明鸢乌贼耳石生长过程中翼区表现出较高的生长率。
笔者研究对鸢乌贼耳石外部形态特征进行了初步的描述与分析,但样本分布海域较小,采集样本个体大小均小于200 mm。在后续鸢乌贼轮纹与年龄的研究中将扩大样本分布范围,以补充和完善南海鸢乌贼耳石形态特征、轮纹与年龄的研究。耳石包含头足类重要的生态信息,温度、盐度、饵料等因素均会影响耳石的形成[20]。结合鸢乌贼样本在时间和空间上的分布,系统地开展鸢乌贼耳石形态特征、轮纹与年龄、微化学等方面的研究。
-
![]()
图 1 南海中部海域鸢乌贼采集站位示意图
Figure 1. Sampling sites of S.oualaniensis in the central South China Sea
![]()
图 2 鸢乌贼耳石形态参数示意图
A.耳石总长;B.吻区长;C.吻区宽;D.背侧区长;E.腹外侧长;F.侧区长;G.翼区长;H.翼区宽;I.最大宽度
Figure 2. Morphometric measurement of S.oualaniensis
A. total statolith length, TSL; B. rostrum length, RSL; C. rostrum width, RSW; D. dorsolateral length, DLL; E. ventrolteral length, VLL; F. lateral dome length, LDL; G. wing length, WL; H. wing width, WW;I. maximum width, MW
![]()
图 3 依据耳石形态结构的鸢乌贼种群在南海中部海域的分布
Figure 3. Distribution of S.oualaniensis populations by statolith morphology in the central South China Sea
![]()
图 4 鸢乌贼耳石形态变化示意图
A.耳石总长626 μm;B.耳石总长819 μm;C.耳石总长1 086 μm
Figure 4. Morphological variation of statolith from S.oualaniensis
A. TSL 626 μm; B. TSL 819 μm; C. TSL 1 086 μm
![]()
图 5 鸢乌贼耳石总长/胴长、吻区长/胴长、翼区长/胴长、最大宽度/胴长、背侧区长/胴长的值与胴长关系
Figure 5. Relationship between ratios of TSL/ML, RSL/ML, WL/ML, MW/ML, DLL/ML and ML of S.oualaniensis
![]()
图 6 鸢乌贼耳石总长、吻区长、翼区长、最大宽度、背侧区长与胴长关系
A.总长/胴长;B.吻长/胴长;C.翼长/胴长;D.最大宽/胴长;E.背侧区长/胴长
Figure 6. Relationship between TSL, RSL, WL, MW, DLL and ML of S.oualaniensis
A.TSL/ML; B.RSL/ML; C.WL/ML; D.MW/ML; E.DLL/ML
![]()
图 7 鸢乌贼耳石背侧区长/总长、侧区长/总长、吻区长/总长、翼区长/总长与胴长的关系
A.(背侧区长/总长)/胴长;B.(侧区长/总长)/胴长;C.(吻区长/总长)/胴长;D.(翼区长/总长)/胴长
Figure 7. Relationship between DLL/TSL, LDL/TSL, RSL/TSL, WL/TSL and mantle length of S.oualaniensis
A.(DLL/TSL)/ML; B.(LDL/TSL)/ML; C.(RSL/ TSL)/ML; D.(WL/TSL)/ML
表 1 鸢乌贼耳石9个形态参数5个主成分负荷值和贡献率
Table 1 Loading and contribution rates of five principal components for nine morphometric measurement of statolith from S.oualaniensis
形态参数
morphometric measurement主成分 principal component 1 2 3 4 5 耳石总长/μm TSL 0.897 0.192 -0.107 -0.278 -0.017 吻区长/μm RSL 0.778 0.412 -0.351 -0.059 -0.198 翼区长/μm WL 0.475 0.536 0.675 0.166 -0.017 最大宽度/μm MW 0.880 -0.133 0.088 0.054 -0.126 背侧区长/μm DLL 0.804 -0.364 0.038 0.176 -0.310 腹外侧长/μm VLL 0.849 0.328 -0.244 0.001 0.243 侧区长/μm LDL 0.752 -0.348 0.305 -0.331 0.271 吻区宽/μm RSW 0.810 -0.174 -0.167 0.412 0.311 翼区宽/μm WW 0.890 -0.239 0.060 -0.067 -0.140 贡献率/% contribution rate 64.36 10.67 8.71 4.74 4.46  下载: 导出CSV
下载: 导出CSV
-
[1] ROPER C F E, SWEENY M J, NAUEN C E. An annotated and illustrated catalogue of species of interest to fisheries [J]. FAO Fish Synop, 1984, 125(3): 157-181. https://xueshu.baidu.com/usercenter/paper/show?paperid=c4720a3ead8c8bc3524f59e266de718b&site=xueshu_se
[2] 刘必林. 利用耳石微结构研究印度洋西北海域莺乌贼的年龄和生长[D]. 上海: 上海水产大学, 2006. https://d.wanfangdata.com.cn/thesis/Y918043 [3] 范江涛, 冯雪, 邱永松, 等. 南海鸢乌贼生物学研究进展[J]. 广东农业科学, 2013, 40(23): 122-128. doi: 10.3969/j.issn.1004-874X.2013.23.030 [4] 张引. Fisheries acoustic studies on the purpleback flying squid resource in the South China Sea [D]. 台北: 台湾大学, 2005. https://scholars.lib.ntu.edu.tw/entities/publication/1da30842-effa-4e39-9f9b-c8e6bab96dfe [5] SEAFDEC. Report of the first meeting of the regional advisory committee on fisheries management in Southeast Asia [R]. Bangkok: Southeast Asian Fisheries Development Center, 2009.
[6] 叶旭昌, 陈新军. 印度洋西北海域鸢乌贼生物学特性初步研究[J]. 上海水产大学学报, 2004, 13(4): 316-322. doi: 10.3969/j.issn.1004-7271.2004.04.006 [7] 田思泉, 钱卫国, 陈新军. 印度洋西北海域鸢乌贼渔获量、渔获率和脱钩率的初步研究[J]. 上海水产大学学报, 2004, 13(3): 224-229. doi: 10.3969/j.issn.1004-7271.2004.03.007 [8] 陈新军, 钱卫国. 印度洋西北海域鸢乌贼资源密度分布的初步分析[J]. 上海水产大学学报, 2004, 13(3): 218-223. doi: 10.3969/j.issn.1004-7271.2004.03.006 [9] 刘必林, 陈新军, 钟俊生. 印度洋西北海域鸢乌贼耳石的形态特征分析[J]. 上海水产大学学报, 2008, 17(5): 604-609. https://d.wanfangdata.com.cn/periodical/shscdxxb200805016 [10] 张宇美, 颜云榕, 卢伙胜, 等. 西沙群岛海域鸢乌贼摄食与繁殖生物学初步研究[J]. 广东海洋大学学报, 2013, 33(3): 56-64. doi: 10.3969/j.issn.1673-9159.2013.03.010 [11] 张鹏, 杨吝, 张旭丰, 等. 南海金枪鱼和鸢乌贼资源开发现状及前景[J]. 南方水产, 2010, 6(1): 68-74. doi: 10.3969/j.issn.1673-2227.2010.01.012 [12] RADTKE R L. Chemical and structural characteristics of statoliths from the short-finned squid Illex illecebrosus [J]. Mar Biol, 1983, 76(1): 47-54. doi: 10.1007/BF00393054
[13] ARKHIPKIN A I, BJORKE H. Ontogenetic changes in morphometric and reproductive indices of the squid Gonatus fabricii (Oegopsida, Gonatidae) in the Norwegian Sea [J]. Pol Biol, 1999, 22(6): 357-365. doi: 10.1007/s003000050429
[14] DURHOLTZ M D, LIPINSKI M R. Influence of temperature on the microstructure of statoliths of the thumbstall squid Lolliguncula brevis [J]. Mar Biol, 2000, 136(6): 1029-1037. doi: 10.1007/s002270000298
[15] ARKHIPKIN A I, BJORKE H. Statolith shape and microstructure as indicators of ontogenetic shifts in the squid Gonatus fabricii (Oegopisda, Gonatidae) from Norwegian Sea [J]. Pol Biol, 2000, 23(1): 1-10. doi: 10.1007/s003000050001
[16] YATSU A, MORI J. Early growth of the autumn cohort of neon flying squid, Ommastrephes bartramii, in the North Pacific Ocean [J]. Fish Res, 2000, 45(2): 189-194. doi: 10.1016/S0165-7836(99)00112-5
[17] ARKHIPKIN A I. Statolith as "black boxes" (life recorders) in squid [J]. Mar Freshw Res, 2005, 56(5): 573-583. doi: 10.1071/MF04158
[18] CHEN X J, LU H J, LIU B L, et al. Age, growth and population structure of jumbo flying squid, Dosidicus gigas, based on statolith microstructure off the Exclusive Economic Zone of Chilean waters[J]. J Mar Biol Assoc UK, 2011, 91 (1): 229-235. doi: 10.1017/S0025315410001438
[19] LIU B L, CHEN X J, CHEN Y, et al. Age, maturation, and population structure of the Humboldt squid Dosidicus gigas off the Peruvian Exclusive Economic Zones [J]. Chin J Oceanol Limnol, 2013, 31(1): 81-91. doi: 10.1007/s00343-013-2036-z
[20] 刘必林, 陈新军, 陆化杰, 等. 头足类耳石[M]. 北京: 科学出版社, 2011: 37. [21] ARKHIPKIN A I, MIKHEEV A. Age and growth of the squid (Sthenoteuthis pteropus) (Oegopsida: Ommastrephidae) from the Central-East Atlantic[J]. J Exp Mar Biol Ecol, 1992, 163(2): 261-276. doi: 10.1016/0022-0981(92)90054-E
[22] CLARKE M R. The cephalopod statolith: an introduction to its form [J]. J Mar Biol Assoc UK, 1978, 58(1): 701-712. doi: 10.1017/S0025315400041345
[23] ARKHIPKIN A I, BIZIKOV V A. Statolith in accelerometers of squid and cuttlefish [J]. Ruthenica, 1998, 8: 81-84. https://www.semanticscholar.org/paper/Statoliths-in-Accelerometers-of-squid-and-Arkhipkin-Bizikov/6f4653b9d2fa5de3acaf3a02801a3706e4ea5a7b
[24] ARKHIPKIN A I. Towards identification of the ecological life style in nektonic squids using statolith morphometry [J]. J Mollus Stud, 2003, 69(3): 171-178. doi: 10.1093/mollus/69.3.171
[25] NESIS K. Cephalopods of seamounts and submarine ridges [M]. OKUTANI T, O′DOR R K, KUBODERA T. Recent advances in cephalopod fisheries biology. [S. l. ]: [s. n. ], 1993: 365-373. https://oceanrep.geomar.de/38052/
-
期刊类型引用(10)
1. 晏磊,李杰,张鹏,杨炳忠,王腾. 基于GAM模型的南海鸢乌贼CPUE时空分布及其与环境因子的关系. 海洋通报. 2021(02): 217-223 .  百度学术
百度学术
2. 许莎莎,唐峰华,任慧敏,李治洪,何利军. 基于Cytb基因的东印度洋鸢乌贼系统发育关系和种群遗传结构. 上海海洋大学学报. 2021(06): 970-980 . 百度学术
3. 朱凯,王雪辉,杜飞雁,张鹏,邱永松. 南海中南部鸢乌贼中型群与微型群形态指标的分析比较. 中国海洋大学学报(自然科学版). 2019(01): 43-54 . 百度学术
4. 江艳娥,陈作志,林昭进,邱永松,张鹏,方展强. 南海海域鸢乌贼中型群与微型群渔业生物学比较. 水产学报. 2019(02): 454-466 . 百度学术
5. 李波,阳秀芬,邱星宇,王锦溪,周倍合,谢嘉仪,颜云榕. 基于耳石形态特征的南海鸢乌贼群体判别分析. 广东海洋大学学报. 2019(02): 58-66 . 百度学术
6. 范江涛,张俊,冯雪,陈作志. 南沙海域鸢乌贼渔场与海洋环境因子的关系. 上海海洋大学学报. 2019(03): 419-426 . 百度学术
7. 金岳,陈新军. 中国近海头足类基础生物学研究进展. 海洋渔业. 2017(06): 696-712 . 百度学术
8. 王雪辉,邱永松,张鹏,杜飞雁. Natural mortality estimation and rational exploitation of purpleback flying squid Sthenoteuthis oualaniensis in the southern South China Sea. Chinese Journal of Oceanology and Limnology. 2017(04): 902-911 . 百度学术
9. 江艳娥,方展强,林昭进,张鹏,陈作志. 南海鸢乌贼耳石微量元素研究. 南方水产科学. 2016(04): 71-79 . 本站查看
10. 朱凯,王雪辉,张鹏,杜飞雁,邱永松. 南海南部鸢乌贼中型群与微型群形态学差异及其判别分析. 热带海洋学报. 2016(06): 82-88 . 百度学术
其他类型引用(5)


计量
- 文章访问数: 3383
- HTML全文浏览量: 212
- PDF下载量: 1994
- 被引次数: 15
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
