
Analysis of RAPD and mitochondrial Cyt b gene sequences of three cultured stocks of Epinephelus coioides from Guangdong Province
-
摘要:
斜带石斑鱼(Epinephelus coioides)为中国南方重要的海水增养殖经济鱼类,但过度捕捞和人工养殖的快速扩张导致了其自然资源衰退和遗传多样性降低。文章采用随机扩增多态DNA(RAPD)和线粒体细胞色素b(Cyt b)基因序列分析方法,对惠州、深圳和湛江3个斜带石斑鱼养殖群体进行了遗传多样性状况和遗传差异的研究。结果表明:1)相比较于线粒体Cyt b基因序列分析,RAPD技术在研究斜带石斑鱼种内遗传变异方面具有更高的灵敏性和多态性;2)相比较于其他已经报道的石斑鱼类,斜带石斑鱼的遗传多样性较低,RAPD检测到的多态位点百分率(P)为40.57%,究其原因,可能是受过度捕捞和人工养殖的影响;3)RAPD检测到的群体间遗传相似度(I)高达0.967 1~0.980 8,而Cyt b的核苷酸多样性仅为0.003 8;斜带石斑鱼养殖群体间的遗传差异显然很小,具有相似的遗传背景。文章还探讨了斜带石斑鱼线粒体Cyt b基因的特点,并针对目前斜带石斑鱼的养殖现状提出了相应保护遗传多样性的对策与建议。
Abstract:Epinephelus coioides is an important mariculture commercial fish in South China. In recent years, due to excessive fishing and rapid expansion of artificial breeding, the natural resources of E.coioides have declined and their genetic diversity has decreased. We applied random amplified polymorphic DNA(RAPD)and mitochondrial cytochrome b(Cyt b)gene sequences analyzing technique to three cultured stocks of E.coioides sampled from Huizhou, Shenzhen and Zhanjiang. The main results show that: 1)RAPD analysis might be more sensitive than mitochondrial Cyt b gene sequence to reveal genetic variations of E.coioides; 2)Compared with the other reported groupers, the genetic diversity of E.coioides might be affected by overfishing and aquaculture. The percentage of polymorphic loci was 40.57% according to RAPD analysis; 3)The genetic identity was 0.967 1~0.980 8 according to RAPD analysis and the nucleotide diversity of Cyt b was only 0.003 8. Small differentiation and similar genetic background were observed among E.coioides stocks. Furthermore, we explored the characteristics of mitochondrial Cyt b of E.coioides and put forward some suggestions and countermeasures for protecting genetic diversity of E.coioides.
-
Keywords:
- Epinephelus coioides /
- RAPD /
- mitochondrial Cyt b /
- genetic variation
-
珠江河口是南海北部最重要的河口型生态系统,是许多经济鱼类产卵、索饵和避难的场所,也是多种经济鱼类入海或溯河洄游的通道,渔业资源丰富且种类繁多[1-2]。近年来,随着粤港澳大湾区经济的不断发展,人类活动频繁,渔业资源被过度开发,鱼类资源量和种类多样性呈下降趋势[3]。为保护珠江河口鱼类资源,确保其可持续利用,亟需开展鱼类多样性的评估和监测。长期以来,鱼类多样性的研究主要是通过对传统的形态学特征进行比较作为分类依据,但由于鱼类的生长具有阶段性,传统的形态学方法对于鱼卵、幼鱼或形态特征被破坏的鱼类鉴定难度较大,即使有完整的成年标本,但遇到形态极其相似的物种,即便经验丰富的分类学家也很难识别[4]。因此,快速、准确的可模式化操作用于物种鉴定的方法显得尤为重要。
DNA条形码技术是一种利用生物体DNA中一段保守片段对物种进行区分的分子鉴定手段,经过近20年的实践和发展,已形成了一套完善的研究体系,能够快速准确地识别鉴定鱼类。而在DNA条形码技术中,DNA条形码的选择十分关键[5]。2005年,Ward[6]提出使用线粒体细胞色素氧化酶I(Cytochrome oxidase subunit I, COI) 基因5'端约652 bp的序列用于鉴定澳大利亚的207种鱼类,表明COI基因能够有效区分不同物种。随后该片段由于大小适宜、进化速度适中、相对保守且检测简单高效等特点,已成为鱼类鉴定的经典条形码。
近年来,DNA条形码技术不断发展并且与环境DNA (Environmental DNA) 结合产生了新型的环境DNA宏条形码 (Meta-barcoding) 技术,该技术由于非侵入性、无污染、检测范围广以及敏感性高等特点,在鱼类多样性研究中得到广泛使用[7-9]。环境DNA宏条形码技术就是从环境样本提取DNA、选取特定序列扩增并进行高通量测序,通过测序分析获得大量OTU序列 (Operational taxonomic units, OTUs),然后通过序列检索比对来鉴定环境中存在的目标生物类群物种组成[10]。该技术最重要的两个条件是选取一段合理的序列扩增,以及一个全面的检索数据库。但由于环境样本中DNA片段小,以及高通量测序长度的限定,就需要比较短的微型条形码。Miya等[11]开发了一套12S rDNA基因的环境DNA通用PCR引物,针对880种鱼类线粒体中具有的163~185 bp超可变区域。该微型条形码即使在只获得少量DNA的情况时也可以很容易地扩增,而且它在物种之间也有很高的差异,现广泛应用于鱼类多样性研究[12-13]。然而12S rDNA序列也并不是完美的,其长度相对于国际上通用的DNA条形码长度 (552~652 bp) 仍太短,可能在某些近缘物种间无法识别或者识别错误[14]。例如Wang等[15]根据12S rDNA序列将中华瞻星鱼 (Uranoscopus chinensis) 鉴定为日本瞻星鱼 (U. japonicus)。
不管是DNA条形码技术还是环境DNA宏条形码技术都十分依赖对比数据库的准确性和全面性[6,13]。目前,鱼类条形码数据库有NCBI (National Center for Biotechnology Information) 和BOLD (Barcode of Life Data),但由于上传门槛低,数据量冗余庞大,经常出现比对错误,制约了条形码技术的应用。因此,本研究选取南海典型河口珠江河口的鱼类作为研究对象,基于线粒体COI和12S rDNA基因构建珠江河口鱼类条形码数据库,并初步探讨其系统发育关系以及两种DNA条形码在珠江河口鱼类物种鉴定中的适用性,以期为珠江河口鱼类的群落动态监测以及物种多样性保护提供技术支持。
1. 材料与方法
1.1 样品采集及形态鉴定
在珠江河口设置采样断面,设置依据是尽可能覆盖整个珠江河口水域以及盐度范围,但由于珠江河口航道繁忙,工程建设多,以及部分海域存在建筑垃圾,所以拖网站位的选取受到一定限制。本研究共设置9个采样站位 (图1),其站位覆盖地区盐度范围为9~32.17,此外在盐度低于9的地区设置过站位,但这些地区基本处于航道,禁止底拖网。样本采集通过底拖网方式进行,采集时间为2020年4月、2020年9月以及2021年4月。样本冷冻运至实验室后,根据《中国鱼类系统检索》[16]进行形态学分类鉴定。随后取背部肌肉保存于95%乙醇中,于−20 ℃保存备用。
1.2 DNA提取与目的片段的扩增
剪取20~30 mg保存的鱼类肌肉,晾干酒精,用海洋动物组织基因组DNA提取试剂盒 (天根,中国) 提取基因组DNA。采用Ward等[6]设计的引物对COI基因片段进行扩增,引物序列为:Fish-F1:5'-TCAACCAACCACAAAGACATTGGCAC-3'和Fish-R1:5'-TAGACTT CTGGGTGGCCAAAGAATCA-3'。采用Miya等[11]设计的引物对12S rDNA基因片段进行扩增,引物序列为:MiFish-F:5'-GTCGGTAAAACTCGTGCCAGC-3'和MiFish-R:5'-CATAGTG GGGTATCTAATCCCAGTT-TG-3'。使用擎科生物Golden Star T6 Super PCR Mix (1.1×) 试剂盒,扩增体系 (总体积25 μL) 为:Mix预混液22 μL,正反向引物各1 μL,DNA模板1 μL。COI序列扩增反应程序:98 ℃预变性2 min;98 ℃变性10 s,56 ℃复性30 s,72 ℃延伸40 s,35个循环;72 ℃延伸5 min。12S rDNA序列扩增反应程序:98 ℃预变性2 min;98 ℃变性10 s,56 ℃复性30 s,72 ℃延伸20 s,35个循环;72 ℃延伸5 min。
PCR产物用2% TBE琼脂糖凝胶电泳检测,将条带清晰PCR产物送往北京六合华大基因科技有限公司进行测序。
1.3 数据库的构建
对照往年珠江河口鱼类资源调查文献[1,17-21],整理得到珠江河口鱼类名录,物种的有效学名以Fishbase数据库 (www.fishbase.org) 为准。为全面构建珠江河口鱼类DNA条形码数据库,未通过拖网采集到的鱼类种类从GenBank (http://www.ncbi.nlm.nih.gov/genbank/) 筛选并下载COI序列和12S rDNA序列,下载标准是优先下载具有全基因组的序列。
1.4 数据分析
通过DNAStar软件包中的SeqMan软件对测得的COI与12S rDNA序列进行人工校对和拼接[22]。校正后的序列经MitoFish、BOLD、GenBank数据库验证,当序列相似度大于99%时,才能定为同种,确保序列的准确性。运用MEGA-X软件将本研究所得序列与GenBank下载序列合并,分析序列特征,计算碱基组成[23];基于Kimura2-parameter (K2P) 双参数模型对种间遗传距离 (Interspecific genetic distance) 和种内遗传距离 (Intraspecific genetic distance) 进行计算,同时使用COI序列和12S rDNA序列根据种内遗传距离与种间遗传距离绘制遗传距离频数分布直方图[24]。采用ABGD (Automatic Barcode Gap Discovery) 软件将全部序列自动分组,相对间隙宽度值取X=0.8,其余参数全部默认[25]。使用PhyloSuit软件中的ModelFinder程序进行核苷酸序列的模型选择,选出最佳模型GTR+F+I+G4,然后使用PhyloSuit软件中的MrBayes进行贝叶斯系统发育树构建[26]。最后使用itol软件对系统树进行展示和标记[27]。站位图的底图来源于广东省标准地图服务子系统 (http://nr.gd.gov.cn/map/bzdt/),地图数据下载于全国地理信息资源目录服务系统 (http://www.webmap.cn/main.do?method=index),采用ArcGis10.6软件绘制后叠加而成。
2. 结果
2.1 序列库概况
通过底拖网采集珠江河口鱼类标本251尾,经过实验获得219条COI序列和247条12S rDNA序列,共鉴定出99种鱼,隶属于6目10科41属。然后从GenBank数据库下载鱼类165条COI序列和128条12S rDNA序列,共得到384条COI序列和375条12S rDNA序列,隶属于17目66科125属172种 [图2,附录A (详见http://dx.doi.org/10.12131/20210210 的资源附件)]。根据往年珠江河口鱼类资源调查文献初步统计出珠江河口鱼类188种,本研究鱼类占比91%。其中一半种类为鲈形目,占绝对优势,其次是鲽形目和鲱形目,共占比20%。其中包括棘头梅童鱼 (Collichthys lucidus)、风鲚 (Coilia mystus)、杜氏棱鳀 (Thryssa dussumieri) 等咸淡水鱼类优势种以及丽叶鲹 (Alepes djedaba)、带鱼 (Trichiurus lepturus)、银鲳 (Pampus argenteus) 等海水鱼类优势种,常见经济种如龙头鱼 (Harpadon nehereus)、花鰶 (Clupanodon thrissa)、皮氏叫姑鱼 (Johnius belangerii) 等也已全部包括[27]。此外在野外收集了软骨鱼纲的2个物种,条纹斑竹鲨 (Chiloscyllium plagiosum) 和日本燕魟 (Gymnura japonica),从GenBank数据库下载了软骨鱼纲1个物种,尖头斜齿鲨 (Scoliodon laticaudus)。
2.2 线粒体COI和12S rDNA序列的序列特征
172种鱼类384条COI序列经比对对齐,获得一致序列长度为614 bp,平均碱基交换/颠换值为1.29,碱基T、C、A、G的平均含量分别为29.17%、28.60%、24.03%、18.20%,A+T (53.20%) 含量均高于G+C (46.80%),表现出明显的碱基组成偏向性,该结果与海洋鱼类COI基因碱基组成特征一致[6]。172种鱼类375条12S rDNA序列经比对对齐后,长度为163~182 bp,平均碱基交换/颠换值为1.09,碱基T、C、A、G的平均含量分别为19.61%、23.97%、33.90%、22.52%,与COI序列一样,存在A+T (53.51%) 含量显著高于G+C (46.49%) 的现象。
2.3 种间与种内遗传距离
根据K2P遗传距离模型计算384条COI序列结果显示 (表1),172个种类COI序列的种内遗传距离为0~1.66%,平均为0.20%;种间遗传距离为2.16%~40.87%,平均为25.54%,种间平均遗传距离是种内的127.7倍。这一结果符合Hebert等[5]提出的“10倍规则”,即满足种间平均遗传距离值大于种内平均遗传距离10倍的标准可用于区分物种。基于375条12S rDNA序列,172种鱼类种内遗传距离为0~1.79%,平均为0.12%;种间遗传距离为0.59%~72.32%,平均为34.39%;种间平均遗传距离是种内的286.6倍。基于COI序列的遗传距离频数分布直方图能形成明显的条形码间隙,而基于12S rDNA序列的遗传距离频数分布直方图不能形成明显的条形码间隙,说明COI序列能对珠江河口鱼类物种进行区分鉴定,而12S rDNA序列在有些物种间存在区分困难的情况 (图3)。
表 1 基于COI和12S rDNA序列的种内与种间遗传距离Table 1. Genetic distance of intraspecies and interspecies based on mitochondrial COI and 12S rDNA sequences% 遗传距离
Genetic distanceCOI序列 COI sequence 12S rDNA序列 12S rDNA sequence 最小值
Minmum最大值
Maxium平均值
Mean最小值
Minmum最大值
Maxium平均值
Mean种内 Intraspecies 0 1.66 0.2 0 1.79 0.12 种间 Interspecies 2.16 40.87 25.54 0.59 72.32 34.39 ![]() 图 3 基于COI基因 (a) 和12S rDNA基因 (b) 的种内和种间K2P遗传距离的频率分布Figure 3. Frequency distributions of intraspecific and interspecific pairwise genetic K2P distance based on COI gene (a) and 12S rDNA gene (b)
图 3 基于COI基因 (a) 和12S rDNA基因 (b) 的种内和种间K2P遗传距离的频率分布Figure 3. Frequency distributions of intraspecific and interspecific pairwise genetic K2P distance based on COI gene (a) and 12S rDNA gene (b)2.4 系统发育关系
基于COI序列构建的贝叶斯系统发育树见图4,在物种较多的科中,石首鱼科、鲾科、舌鳎科、带鱼科、鲹科、鲻科可各自形成科的单系支,但在鲀科、鳀科、鰕虎鱼科、鲱科、狗母鱼科中个别物种存在互相交叉的现象,与传统分类存在一些分歧。在属水平上,绝大多数同属物种都能各自聚在一起,仅有小公鱼属 (Stolephorus),䱨属 (Terapon) 和细棘鰕虎鱼属 (Acentrogobius) 3个属的物种未能聚成独立的小分支。软骨鱼纲的3种物种可以形成一个单系支。当种内先验最大遗传距离P为1.29%时,384条DNA序列根据ABGD分析显示有172个分类单元,与本研究物种数量一致。
基于12S rDNA序列构建的贝叶斯系统发育树见图5,在物种较多的科中,带鱼科、舌鳎科、鳀科、鲾科、鲻科、鲀科可各自形成单系支,但在狗母鱼科、鰕虎鱼科、石首鱼科、鲱科、鲹科中聚类效果不明显;软骨鱼纲3种物种可形成一个单系支。在属的水平上,绝大多数同属的物种都能各自聚在一起,仅有细棘鰕虎鱼属、舌鳎属 (Cynoglossus)、蛇鲻属 (Saurida) 和兔头鲀属 (Lagocephalus) 4个属未聚成小分支。软骨鱼纲的3种物种可以形成一个单系支。当种内先验最大遗传距离P为1.29%时,375条DNA序列根据ABGD分析显示有166个分类单元,将带鱼和南海带鱼 (T. nanhaiensis) 聚为一个分类单元,大甲鲹 (Megalaspis cordyla) 和六带鲹 (Caranx sexfasciatus) 聚为一个分类单元,弓斑东方鲀 (Takifugu ocellatus)、星点东方鲀 (T. niphobles) 和黄鳍东方鲀 (T. xanthopterus) 聚为一个分类单元,翼红娘鱼 (Lepidotrigla alata) 和绿鳍鱼 (Chelidonichthys kumu) 聚为一个分类单元,乌鲹 (Parastromateus niger) 和马拉巴若鲹 (Carangoides malabaricus) 聚为一个分类单元,剩余161个由单个物种组成,占比93.6%。这些结果说明COI序列能很好地划分鱼类物种,而12S rDNA序列对于少数近缘物种无法有效划分。
3. 讨论
为了使DNA条形码技术和环境DNA宏条码技术在珠江河口鱼类多样性的研究中得到充分和有效的应用,本研究建立了珠江河口本地鱼类的COI和12S rDNA宏条形码数据库。而本地数据库的有效构建需到达到2个条件[27]:1) 数据库所含物种基本覆盖本地已出现的物种;2) 足够的核苷酸序列特异性来区分密切相关的物种[28]。
3.1 数据库的物种覆盖度
本研究获得172种鱼类物种序列,占珠江河口统计鱼类的91%。李因强[29]研究发现珠江河口鱼类优势种主要为棘头梅童鱼、凤鲚等,以及皮氏叫姑鱼、丽叶鲹等一些常见经济种,本研究已全部包括。剩余16种鱼类未在野外采集到样本,可能是由于本研究的采集方式为底拖网,该方式主要采集底栖鱼类,例如斑点马鲛 (Scomberomorus guttatus) 和革似鲹 (Scomberoides tol) 这种上层鱼类很难捕获[30]。另一方面,GenBank数据库和Bold数据库没有详细的数据可供下载,所以没有进行收集。此外这些鱼类在该地区基本属于偶见种、非经济种,对本研究影响不大。在今后的研究中将持续不断地完善数据库。
3.2 COI条形码在珠江河口鱼类物种鉴定中的适用性
目前已经有多项关于COI条形码在鱼类鉴定的报道。2021年罗纯等[31]使用COI序列研究中日沿海部分鱼类DNA条形码时,其种间与种内平均遗传距离比值为14.81,符合“10×”的阈值规则,可实现物种的高效鉴定;郜星晨和姜伟[32]在2020年使用COI序列建立三峡鱼类DNA条形码数据库时,其种间平均遗传距离为种内的9.38倍,表示COI基因可以作为三峡库区常见鱼类鉴定的有效条形码基因。
本研究中,COI序列种间平均遗传距离是种内的127.7倍,也十分符合“10×”的阈值规则。与上述研究相比,远大于其他研究中的遗传距离比值,这可能由于不同地区鱼类活动范围不同,基因交流不同,所以遗传距离有所差异[33-34]。还有可能就是本实验每种鱼类采样量不足,部分种类COI仅1条,未收集到物种内所有序列差异,导致种内平均遗传距离偏小。此外根据遗传距离频数分布直方图显示COI序列已形成明显的条形码间隙,同时根据ABGD正好将172种鱼类分成172个分类单元,所以COI序列在本研究中具有较好的物种鉴别率,可以满足珠江河口鱼类精准识别的需求。
3.3 12S rDNA条形码在珠江河口鱼类物种鉴定中的适用性
虽然12S rDNA序列种间平均遗传距离是种内的286.6倍,也十分符合“10×”的阈值规则。但是遗传距离直方图显示,种间遗传距离与种内遗传距离发生了重叠,未形成明显的条形码间隙。此外ABGD将一些近缘物种划分到了同一个分类单元,例如,本研究东方鲀属的3个物种根据ABGD划分为一个分类单元,且黄鳍东方鲀、星点东方鲀与弓斑东方鲀之间的遗传距离介于0.59%~1.2%,在GenBank上相似度高达99%以上,只有2个碱基的差别。因为基因会发生交换和重组,所以即使同一个物种间,12S rDNA序列有时也会存在几个碱基的差异,将导致该类型物种无法区分开来[35]。在Zou等[8]使用12S rDNA序列对南沙湿地的环境DNA研究中,出现了双斑东方鲀 (T. bimaculatus) 和铅点东方鲀 (T. alboplumbeus),与本研究东方鲀属的3个物种也仅有1~3个碱基的差异,所以很有可能会出现低估鱼类多样性的情况。此外在带鱼与南海带鱼两个亚种的区分上,ABGD也将两者划分为一个物种,且两者遗传距离为1.8%,仅有3个碱基的差别,说明12S rDNA序列可能无法对亚种进行区分。在本研究172个鱼类物种中,11个物种存在以上情况,占6.4%,说明12S rDNA序列还是可以用于大部分珠江河口鱼类的种类鉴定。但是在今后的研究中,区分一些近缘种类需特别注意,应当使用COI序列加以验证,并参考当地鱼类名录,调查是否出现过该物种而后加以确认。
综上所述,根据COI序列和12S rDNA序列建立的珠江河口鱼类DNA条形码数据库已经达到数据库建立的两个条件。同时,证明了COI条形码能很好地适用于珠江河口鱼类鉴定,并且论证了线粒体12S rDNA条形码适用于大部分珠江河口鱼类的种类鉴定,但是对少数亲缘关系近的物种可能难以区分,这可能会导致鱼类多样性的低估,在今后的研究中应特别注意此类物种。
-
![]()
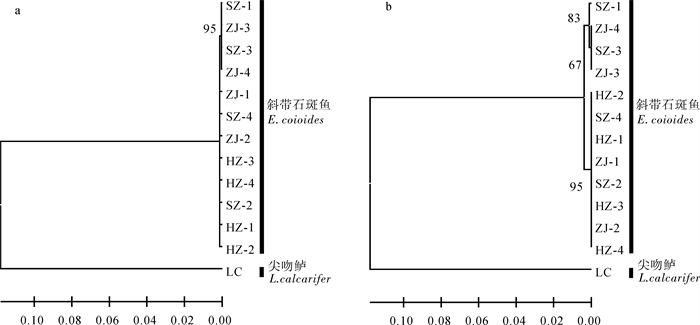
图 1 基于斜带石斑鱼线粒体Cyt b基因的NJ(a)和UPGMA(b)系统树
分枝节点处的数值为1 000次Bootstrap置信值
Figure 1. Phylogenetic tree of E.coioides based on partial mitochondrial Cyt b gene sequences by NJ (a)and UPGMA (b)methods
Bootstrap values (1 000 replicates)are shown at nodes.
表 1 RAPD和Cyt b引物序列
Table 1 Primer sequences of RAPD and Cyt b
RAPD引物序列(5′→3′)
RAPD primer sequence(5′→3′)Cyt b引物序列(5′→3′)
Cyt b primer sequence(5′→3′)S165-TGTTCCACGG S183-CAGAGGTCCC S241-ACGGACGTCA L14724-5′-CGAAGCTTGATATGAAAAACCATCGTTG-3′ S168-TTTGCCCGGT S190-ACCGTTCCAG S242-CTGAGGTCTC S178-TGCCCAGCCT S192-CTGGGTGAGT S245-TTGGCGGCCT S180-AAAGTGCGGC S193-GTCGTTCCTG S248-GGCGAAGGTT H15149-5′-AAACTGCAGCCCCTCAGAATGATATTTGTCCTCA-3′ S181-CTACTGCGCT S195-CAGTTCACGG S250-ACCTCGGCAC S182-CCTCTGACTG S200-TCTGGACGGA  下载: 导出CSV
下载: 导出CSV
表 2 斜带石斑鱼的遗传变异
Table 2 Genetic variation of E.coioides
群体
stock多态位点百分率(P)/%
percentage of polymorphic loci香农信息指数(i)
Shannon′s information index基因多样性(h)
gene diversity惠州 HZ 31.13 0.150 6 0.098 1 深圳 SZ 32.08 0.163 8 0.108 7 湛江 ZJ 28.30 0.142 9 0.094 8
下载: 导出CSV
表 3 基因多样性(对角线上方)和香农信息指数(对角线下方)的t检验
Table 3 T-test for gene diversity (above diagonal) and Shannon′s information index (below diagonal)
群体 stock 惠州群体 HZ stock 深圳群体 SZ stock 湛江群体 ZJ stock 惠州 HZ - 0.651 2 0.885 5 深圳 SZ 0.699 4 - 0.559 3 湛江 ZJ 0.819 4 0.545 6 -
下载: 导出CSV
表 4 遗传相似度(对角线上方)和遗传距离(对角线下方)
Table 4 Genetic identity (above diagonal) and genetic distance (below diagonal)
群体 stock 惠州群体 HZ stock 深圳群体 SZ stock 湛江群体 ZJ stock 惠州 HZ - 0.975 1 0.980 8 深圳 SZ 0.025 2 - 0.967 1 湛江 ZJ 0.019 4 0.033 5 -
下载: 导出CSV
表 5 斜带石斑鱼线粒体Cyt b基因的碱基组成
Table 5 Base composition of mitochondrial Cyt b gene of E.coioides
密码子 codon T C A G A+T 第一位点 the first site 24.8 24.1 25.5 25.5 54.3 第二位点 the second site 38.3 22.7 21.3 17.7 50.3 第三位点 the third site 22.3 45.1 30.5 2.1 59.6 平均 mean 28.5 30.6 25.8 15.1 52.8
下载: 导出CSV
-
[1] HEEMSTRA P C, RANDALL J E. Groupers of the world (Family Serranidae, Subfamily Epinephelinae): an annotated and illustrated catalogue of the grouper, rockcod, hind, coral grouper and lyretail species known to date[J]. FAO Fisheries Synopsis, 1993, 16(125): 42-48. https://www.ingentaconnect.com/contentone/umrsmas/bullmar/1995/00000056/00000001/art00028
[2] RUSSELL B C, HOUSTON W. Offshore fishes of the Arafura Sea[J]. Beagle, 1989, 6(1): 69-84. https://www.biodiversitylibrary.org/part/271273
[3] IUCN. IUCN red list categories and criteria[M]. Version 3.1. Gland and Cambridge: IUCN Species Survival Commission, 2001: 60-62. https://xueshu.baidu.com/usercenter/paper/show?paperid=bb12f246e77af3a0c9cf975b07fa1179&site=xueshu_se
[4] SADOVY Y. Regional survey for fry/fingerling supply and current practices for grouper mariculture: evaluating current status and long-term prospects for grouper mariculture in South East Asia. Final report to the Collaboration APEC grouper research and development network (FWG 01/99)[R]. Singapore: APEC, 2000: 5-8. https://www.researchgate.net/publication/242511231_Regional_survey_for_fryfingerling_supply_and_current_practices_for_grouper_mariculture_evaluating_current_status_and_long-term_prospects_for_grouper_mariculture_in_South_East_Asia
[5] 陈大玮, 邓利, 刘志刚, 等. 斜带石斑鱼抗菌肽hepcidin基因克隆及其成熟肽的原核融合表达[J]. 南方水产科学, 2011, 7(1): 1-7. doi: 10.3969/j.issn.2095-0780.2011.01.001 [6] FERAL J P. How useful are the genetic markers in attempts to understand and manage marine biodiversity[J]. J Exp Mar Biol Ecol, 2002, 268(2): 121-145. doi: 10.1016/S0022-0981(01)00382-3
[7] 林浩然. 重要海水养殖鱼类遗传多样性与种质基因组的研究[J]. 科技导报, 2004(9): 4-6. doi: 10.3321/j.issn:1000-7857.2004.09.002 [8] LIU Y G, CHEN S L, LI B F, et al. Analysis of genetic variation in selected stocks of hatchery flounder, Paralichthys olivaceus, using AFLP markers[J]. Biochem Syst Ecol, 2005, 33(10): 993-1005. doi: 10.1016/j.bse.2005.02.008
[9] 区又君, 吴勇, 李加儿, 等. 5种石斑鱼遗传差异的RAPD分析[J]. 南方水产, 2008, 4(3): 56-62. doi: 10.3969/j.issn.2095-0780.2008.02.009 [10] ANTORO C, NA-NAKOM U, KOEDPRANG W. Study of genetic diversity of orange-spotted grouper, Epinephelus coioides, from Thailand and Indonesia using microsatellite markers[J]. Mar Biol, 2006, 8(1): 17-26. doi: 10.1007/s10126-005-5026-0
[11] 蒙子宁, 杨丽萍, 吴丰, 等. 斜带石斑鱼、赤点石斑鱼RAPD和线粒体Cyt b基因序列变异分析[J]. 中山大学学报: 自然科学版, 2007, 46(1): 44-48. doi: 10.3321/j.issn:1672-3554.2007.01.010 [12] 蒙子宁, 杨丽萍, 吴丰, 等. 斜带石斑鱼养殖群体遗传多样性的RAPD分析[J]. 热带海洋学报, 2007, 26(2): 75-80. doi: 10.3969/j.issn.1009-5470.2007.02.008 [13] 骆剑, 黄宗文, 陈国华, 等. 南海斜带石斑鱼野生群体与人工繁殖群体的微卫星分析[J]. 热带生物学报, 2011, 2(2): 123-128. doi: 10.3969/j.issn.1674-7054.2011.02.007 [14] WILLIAMS J G K, KUBELIK A R, LIVAK K J, et al. DNA polymorphisms amplified by arbitrary primers are useful as genetic markers[J]. Nucleic Acids Res, 1990, 18(24): 6531-6533. https://pubmed.ncbi.nlm.nih.gov/1979162/
[15] WELSH J, MCCLELLAND M. Fingerprinting genomes using PCR with arbitrary primers[J]. Nucleic Acids Res, 1990, 18(24): 7213-7218. doi: 10.1093/nar/18.24.7213
[16] 杨慧荣, 赵会宏, 蒙子宁, 等. 赤眼鳟线粒体D-loop和Cyt b基因序列的对比分析[J]. 中山大学学报: 自然科学版, 2012, 51(5): 100-107. https://d.wanfangdata.com.cn/periodical/zsdxxb201205019 [17] 王培欣, 罗建仁, 白俊杰, 等. 斗鱼属鱼类亲缘关系的Cyt b基因序列和RAPD分析[J]. 动物学杂志, 2009, 44(5): 14-23. https://d.wanfangdata.com.cn/periodical/dwxzz200905003 [18] ZVYCHAINAIA E, DANIKIN A A, KHOI ODOVA M V, et al. Analysis of the variability of the control region and cytochrome b gene of mtDNA of Capreolus pygargus[J]. Izv Akad Nauk Ser Biol, 2011, 38(5): 511-517. doi: 10.1134/S1062359011050189
[19] SAMBROO J, FRITSCH E F, MANIATIS T. Molecular cloning: a laboratory manual[M]. 2nd ed. Cold Spring: Harbor Laboratory Press, 1989: 55-56. https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1454184/
[20] LIU Z J, CORDES J F. DNA marker technologies and their applications in aquaculture genetics[J]. Aquaculture, 2004, 238(1): 1-37. https://www.researchgate.net/publication/222393708_DNA_marker_technologies_and_their_applications_in_aquaculture_genetics
[21] BARDAKCI F, SKIBINSKI D O F. Application of the RAPD technique in tilapia fish: species and subspecies identification[J]. Heredity, 1994, 73(2): 117-123. doi: 10.1038/hdy.1994.110
[22] DINESH K R, LIM T M, CHUA K L, et al. RAPD analysis: an efficient method of DNA fingerprinting in fishes[J]. Zool Sci, 1993, 10(5): 849-854. https://pubmed.ncbi.nlm.nih.gov/7764373/
[23] JOHANSEN S, GUDDAL P H, JOHANSEN T. Organization of the mitochondrial genome of Atlantic cod, Gadus morhua[J]. Nucleic Acids Res, 1990, 18(3): 411-419. doi: 10.1093/nar/18.3.411
[24] GRAY M W. Origin and evolution of mitochondrial DNA[J]. Ann Rev Cell Biol, 1989, 5: 25-50. doi: 10.1146/annurev.cb.05.110189.000325
[25] 邵爱华, 朱江, 陈葵, 等. 暗纹东方鲀线粒体细胞色素b及其侧翼tRNA基因的克隆与序列分析[J]. 中国水产科学, 2005, 12(6): 675-681. doi: 10.3321/j.issn:1005-8737.2005.06.002 [26] 郑莲, 刘楚吾. 蜂巢石斑鱼随机扩增多态性DNA的初步研究[J]. 湛江海洋大学学报, 2002, 22(4): 14-18. doi: 10.3969/j.issn.1673-9159.2002.04.002 [27] PANTE M J R, GIERDE B, MCMILLAN I. Effect of inbreeding on body weight at harvest in rainbow trout (Oncorhynchus mykiss)[J]. Aquaculture, 2001, 192(2): 201-211. https://www.sciencedirect.com/science/article/pii/S0044848600004671
[28] 黎中宝, 方秀, 陈锦, 等. 大黄鱼(Pseudosciaena crocea)养殖群体遗传多样性的降低[J]. 海洋与湖沼, 2009, 40(4): 446-450. doi: 10.3321/j.issn:0029-814X.2009.04.009 -
期刊类型引用(11)
1. 王大伟,邢盈,蒋成宇,胡淼,赵金良. 放养密度对鳜幼鱼HPI轴激素、应激酶活力及呼吸频率的影响. 淡水渔业. 2025(02): 26-32 .  百度学术
百度学术
2. 何静怡,魏涯,岑剑伟,郝淑贤,陈胜军,黄卉,赵永强,王悦齐,杨少玲,林织. 基于梯度降温的草鱼暂养及有水保活运输技术. 食品科学. 2024(04): 271-278 . 百度学术
3. 何静怡,郑伟,黄卉,岑剑伟,赵永强,王田,魏涯,郝淑贤,杨少玲,陈琛. 不同温度、盐度条件对草鱼暂养及应激保活的影响. 大连海洋大学学报. 2024(04): 597-605 . 百度学术
4. 梁雪莹,郑晓婷,陈秋羽,谢静怡,董宏标,李勇,杨金龙,陈成勋,张家松. 模拟运输胁迫对牛蛙幼蛙生理机能的影响. 广东海洋大学学报. 2024(06): 118-126 . 百度学术
5. 刘浩,李洁,李亚军,康鹏天,张国维,邵东宏,王建福. 水体中泥沙含量对虹鳟生存、生理和体表微生物的影响. 中国水产科学. 2024(11): 1365-1374 . 百度学术
6. 唐忠林,张佳佳,周国勤,陈树桥,徐钢春,徐跑,强俊,王佩佩. 丁香酚对“优鲈3号”幼鱼运输水质及其血液、肌肉生理指标的影响. 水产科技情报. 2023(01): 44-52 . 百度学术
7. 王文雯,杨静茹,付正祎,于刚,马振华. 运输时间对高体鰤幼鱼应激、代谢、抗氧化和免疫的影响. 中国渔业质量与标准. 2023(02): 25-36 . 百度学术
8. 李哲,周珊珊,王好学,王嘉浩,陈璐,徐开达. 运输振荡对条石鲷幼鱼生理应激和水体总氨氮含量的影响. 水产科技情报. 2023(05): 327-332 . 百度学术
9. 范宏博,胡丁月,徐莉,孙浩,刘峰,刘春娥,张小栓. 运输密度和时间对单环刺螠保活运输品质的影响. 农业工程. 2023(10): 72-77 . 百度学术
10. 虞为,陈雪晴,杨育凯,张燕娃,黄小林,黄忠,李涛,马振华,吴洽儿,于刚,周传朋,林黑着. 饲料中添加雨生红球藻对尖吻鲈生长性能、抗氧化能力及免疫状态的影响. 南方水产科学. 2022(05): 46-54 . 本站查看
11. 陈旭,赵旺,陈明强,谭春明,于刚. 盐度胁迫对红娇凤凰螺耗氧率、排氨率以及免疫相关酶活性的影响. 南方水产科学. 2022(05): 160-165 . 本站查看
其他类型引用(4)





计量
- 文章访问数: 3478
- HTML全文浏览量: 123
- PDF下载量: 1706
- 被引次数: 15
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
