
Assessment of economic and ecological benefits for mesh enclosure isolated white shrimp-common mullet polyculture
-
摘要:
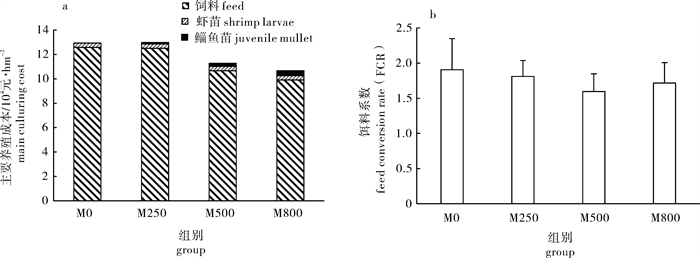
为探索新型对虾生态养殖模式,于池塘中构建围网开展了凡纳滨对虾(Litopenaeus vannamei)-鲻(Mugil cephalus)混养试验。鲻密度依次为0尾(M0,对照组)、250尾(M250)、500尾(M500)和800尾(M800)。结果显示,鲻放养组的饵料系数均比对照组低,但组间差异不显著(P>0.05)。M250组对虾产值(30.30×104元·hm-2)及总产值(31.02×104元·hm-2)为各组最高,M800组对虾产值(20.96×104元·hm-2)及总产值(23.30×104元·hm-2)最低。试验结束后,放养较多鲻的处理组(M500和M800)的总有机碳(TOC)有约20 mg·L-1幅度的下降,而M0和M250则变动极小,但组间差异不显著(P>0.05)。养殖末期鲻放养组的亚硝态氮(NO2-N)质量浓度均高于对照组M0,且M250组NO2-N及氨氮(NH4-N)质量分数居于各组最高(P>0.05)。试验表明,在池塘围网混养少量鲻(M250)时对虾产量最高,而混养较多鲻(M500和M800)时饲料成本较为节约且水质指标较好。采取养殖前期多放鱼,后期适当分鱼以降低密度的措施,可能更有利于虾塘经济与生态效益的提高。
Abstract:In order to explore a new ecological culture mode for shrimps, we developed a white shrimp (Litopenaeus vannamei)-common mullet (Mugil cephalus) polyculture system in shrimp ponds. The stocking density of common mullets for each treatment was 0 (M0), 250 (M250), 500 (M500) and 800 (M800), respectively, in which M0 was the control without mesh enclosure inside the pond. The results show that the feed conversion ratios (FCR) of treatments with common mullets were all lower than the FCR of the control, but no significant difference was found among treatments (P>0.05). Treatment M250 had the highest shrimp production value (¥30.30×104 yuan·hm-2) and gross production value (¥31.02×104 yuan·hm-2), while treatment M800 had the lowest shrimp production value (¥20.96×104 yuan·hm-2) and gross production value (¥23.30×104 yuan·hm-2), though there were no significant differences among treatments (P>0.05). TOC value in treatments with more common mullets (M500 and M800) decreased by about 20 mg·L-1 at final culture stage but changed very little in treatments M0 and M250, and no significant difference was found among treatments (P>0.05). Treatments with common mullets had higher concentration of NO2-N than the control at final culture stage, and M250 had the highest NO2-N and NH4-N concentrations among all treatments. The results indicate that the polyculture system with fewer common mullets (M250) may obtain better shrimp production, while it is more feed-saving and has better water quality control with more fish stocking (M500 and M800). The economic and ecological benefits might be improved by stocking more fish at early culture stage and decreasing the fish stocking density at later culture stage with this mesh enclosure isolated white shrimp-common mullet co-culture system.
-
Keywords:
- shrimp /
- mullet /
- mesh enclosure /
- polyculture /
- water quality /
- economic benefits
-
铜藻Sargassum horneri(Turn)属褐藻门马尾藻属,为北太平洋西部特有的暖温带性大型海藻,生长在低潮带深沼中或大干潮线深至4 m处的岩石上[1]。铜藻不仅是海藻工业的优质原料,还被广泛应用到医药、食品、饲料和有机肥料等方面,具有较高的商业价值。铜藻具有生长快速和生物量巨大的特点,因而被海洋生态学家列为重建海底藻场和实施海洋生态修复的重要物种之一[2]。目前,关于铜藻的报道多集中在对其繁殖、生理和生态方面的研究[2-4]。中国是一个海藻养殖大国,从暖温带到亚热带,每年鲜海藻产量高达400万t,铜藻作为一个潜在的人工栽培海藻物种,相应的胁迫生理研究目前是空白,而幼苗时期对各种不良环境条件的反应和相应处理措施通常是实现一种海藻人工栽培所必须要了解的。
叶绿素荧光技术(chlorophyll fluorescence measurements)是20世纪80年代崛起的一种有效、操作简单、对受试对象无伤害的快速光合生理测定技术,被广泛应用在陆地作物和海藻胁迫生理的研究领域[5]。该技术可以在不破损活体样品(甚至不发生任何接触)的情况下用来了解海藻光合生理状态,判断其转化光能的效率,测定电子传递的速率,估计其受环境条件胁迫(如温度、光照等)的程度。因此,叶绿素荧光技术不仅可以应用于基础光合作用研究领域,而且可以应用到植物生态生理学领域。2007~2008年在全人工条件下,采用同步受精技术实现了大规模铜藻苗种培育。此项研究的主要目的是利用叶绿素荧光测量技术了解全人工铜藻幼苗在不同温度、光照强度和盐度等胁迫条件下光合效率的表现,从而为今后苗种的大规模栽培提供直接技术支持。
1. 材料与方法
1.1 试验材料
2007~2008年在中国科学院海洋研究所海洋生物种质库种质培养室80 L(65 cm×45 cm×35 cm)PP水箱中人工培育的铜藻幼苗,按照大小相近原则随机挑选健康藻体,质量0.039±0.05 g,藻体叶片4~6片,最长叶长1.78±0.02 cm,主茎长度约0.3 cm。
1.2 试验方法
用Mini-PAM光量子分析仪(Walz Effeltrich,德国产)测量叶绿素荧光参数,包括PSII最大荧光量子产量(Fv/Fm)和即时叶绿素荧光量子产量(Fm′)。测量经温度、室外太阳直射光以及高、低盐度处理样品的Fv/Fm值,基本方法参照PANG等[6],将藻体分别置于9 cm培养皿中,培养皿中加入20 mL PES(provasli enriched seawater),用载玻片(76.2 mm×25.4 mm×1.2 mm)盖压藻体,从而保证测量时,光源和藻体距离恒定。测量前首先将样品暗适应20 min。测量时PAM光纤探头的末端垂直对准藻体基部。测量室内消减的太阳光条件下培养藻体的Fm′的测量方法参考图 1。将5棵藻体用橡皮筋分别固定在图 1所示的5个斜面装置下部平台上,藻体基部对准斜面小孔,保证PAM光纤末端以固定的距离对准藻体基部,5个重复的斜面装置并排放到水箱中,藻体没于水面下5 cm。固定的距离可以保持5个重复测量的一致性。除特殊说明均用光度计(SAMPO-LX1330A)记录光照强度,紫外强度计(TN-2340)记录紫外光强度,溶解氧电导仪记录温度。
![]() 图 1 叶绿素荧光量子产量测量装置Figure 1. Design for the measurement of chlorophyll fluorescence quantum yields
图 1 叶绿素荧光量子产量测量装置Figure 1. Design for the measurement of chlorophyll fluorescence quantum yields1.3 试验设计
1.3.1 温度试验
试验1设置3个温度(25、30和35℃)和室温对照(18℃)。20棵铜藻分别投入2 L烧杯,每烧杯5棵,加入1 600 mL PES,放入25±0.5、30±0.5和35±0.5℃光照培养箱(宁波江南仪器厂)和室温培养架上(18±0.5℃),通气培养。盐度30.8,光强90~100 μmol photons · m-2 · s-1,除特殊说明此试验光照周期均为光照12 h · d-1。分别测量在胁迫培养1 h后和在对照条件下恢复24 h后的Fv/Fm。试验2设置3个温度(26、28和30℃)和室温对照(18℃)。20棵铜藻分别投入2 L烧杯,每烧杯5棵,加入1 600 mL PES,放入26±0.5、28±0.5和30±0.5℃光照培养箱和室温培养架上,通气培养10 d后,在对照条件下恢复培养24 h,每天更换培养液。盐度29.8~31.9,光照强度同试验1。每天测量Fv/Fm,记录试验个体的初始和最终重量(g)。试验3设置3个温度(22、24和26℃)和室温对照(18℃)。20棵铜藻分别投入2 L烧杯,每烧杯5棵,加入1 600 mL PES,放入22±0.5、24±0.5和26±0.5℃光照培养箱和室温培养架上,通气培养。10 d后恢复培养24 h,每天更换PES。光照强度同试验1。每天测量Fv/Fm,记录初始和最终重量(g)和最长叶长(cm)。
1.3.2 光照胁迫
(1) 室外实验。将铜藻幼苗固定在长塑料板(57.5 cm×9.8 cm×1 cm)上,置于室外水箱中(200 cm×57 cm×50 cm),藻体没于水下约5 cm,在太阳直射光下培养1、2、3、4、5和6 h后分别取藻体5棵,测量Fv/Fm,全程记录直射光和紫外线强度(280~320 nm),之后,室内通气恢复培养24 h,光强98 μmol photons · m-2 · s-1,测量Fv/Fm。室外水箱水温变化范围17.5~22.9℃。(2)室内消减的太阳光胁迫(直射太阳光的70%,不含紫外线)。试验在室内80 L翻滚式充气培养PP水箱(82 cm×40 cm×30 cm)中进行,室内可接受70%日光照射[6]。将5棵藻体用橡皮筋分别固定在图 1所示的5个斜面装置中,藻体基部对准斜面小孔,保证PAM光纤末端对准藻体基部,将装置并排放到水箱中,藻体没于水面下5 cm处,从7: 00~18: 00每1 h测量藻体的即时荧光产量(Fm′)。用球形光度计(Biospherical,美国产)在线记录室内光强。水箱水温17.6℃,盐度30.6。
1.3.3 盐度胁迫
(1) 高盐胁迫。试验设置3个盐度(30、45和60),其中盐度30作为对照。60棵铜藻分别投入2 L烧杯,每烧杯20棵,加入1 600 mL PES,培养架上(18±0.5℃)通气培养。光强90~100 μmol photons · m-2 · s-1。处理30 min,1、3和6 h后,每个处理随机取样5棵,测量Fv/Fm。恢复培养24 h后测Fv/Fm。(2)低盐胁迫。试验设置4个盐度(30、21、9和0),30作为对照。80棵铜藻分别投入2 L烧杯,每烧杯20棵,加入1 600 mL PES,培养架上18±0.5℃通气培养。光强90~100 μmol photons · m-2 · s-1。处理30 min,1、3和6 h后,每个处理取样5棵,测量Fv/Fm。恢复培养24 h后测Fv/Fm。
1.4 数据处理
试验数据经One-Way ANOVA统计处理,P < 0.05表示显著差异。
2. 结果
2.1 温度试验
2.1.1 温度试验1
和对照相比,35℃胁迫1 h铜藻的Fv/Fm降到对照的63%,经24 h恢复仅为4.96%,个别参数甚至降到0(图 2)。原本黄褐色的叶片明显萎焉苍白,个别叶片甚至脱落,短时间35℃高温会明显不可逆转的破坏铜藻幼苗光合系统。
![]() 图 2 3种温度条件下处理1 h及修复24 h后铜藻幼苗的Fv/Fm(n=5)Figure 2. Optimal chlorophyll fluorescence quantum yields (Fv/Fm)of seedlings of S.horneri after exposure to three temperatures for 1 h respectively and the corresponding recovered values after 24 h
图 2 3种温度条件下处理1 h及修复24 h后铜藻幼苗的Fv/Fm(n=5)Figure 2. Optimal chlorophyll fluorescence quantum yields (Fv/Fm)of seedlings of S.horneri after exposure to three temperatures for 1 h respectively and the corresponding recovered values after 24 h2.1.2 温度试验2
和对照相比,10 d内经28℃处理铜藻幼苗的Fv/Fm明显下降(P < 0.05),且叶片明显萎缩,但24 h恢复后的Fv/Fm同对照相比无显著差异。30℃处理后以及经过24 h恢复后的Fv/Fm同对照相比有显著差异(P < 0.05) (表 1);28℃处理组生长速度仅为0.83% · d-1,30℃处理组生长速度呈负值,为-0.88% · d-1 (表 2)。可以看出在此试验条件下铜藻幼苗生长的温度上限为28℃。
表 1 3种温度条件下跟踪铜藻幼苗10 d, 起始和24 h修复后的Fv/Fm(n=5)Table 1. Initial and final optimal chlorophyll fluorescence quantum yields (Fv/Fm) of the seedlings of S.horneri after exposed to three temperatures for 10 d and the corresponding recovered values after 24 hMeans±SD 时间time 18℃(对照) (control) 26℃ 28℃ 30℃ 初始值initial value 0.73±0.01 0.73±0.01 0.72±0.02 0.72±0.01* 第10天tenth day 0.73±0.01 0.72±0.01 0.66±0.02* 0.55±0.02* 恢复24 h 24 h recovery 0.73±0.01 0.72±0.01 0.68±0.02 0.53±0.02* 注:*. 与对照相比有显著差异(P < 0.05)
Note:*. statistically significant with the initial control (P < 0.05)表 2 3种温度条件下处理10 d后铜藻幼苗的相对生长速度(n=5)Table 2. Relative growth rates in weights of S.horneri seedlings after exposure to three temperatures for 10 days18℃(对照) (control) 26℃ 28℃ 30℃ 起始平均重量/g initial average weight 0.046±0.01 0.045±0.01 0.046±0.01 0.046±0.01 最终平均重量/g ultimate average weight 0.072±0.02 0.058±0.01* 0.050±0.01* 0.043±0.05* 相对生长速度/%·d-1 relative growth rate 4.48 2.56 0.83 -0.88 注:*.与对照相比有显著差异(P < 0.05)
Note:*.statistically significant with the initial control (P < 0.05)2.1.3 温度试验3
22、24和26℃处理10 d的幼苗的鲜重增长速度为4.53,5.99和2.59% · d-1,叶长增长的速度为2.32,2.04和0.52% · d-1,22和24℃处理的最终平均重量和叶长同对照相比有显著差异(P<0.05)(表 3)。因此,在此试验条件下,铜藻幼苗最适生长温度是22~24℃。
表 3 3种温度条件下处理10 d后铜藻幼苗的生长速度(n=5)Table 3. Relative growth rates in weights and the longest leaves of S.horneri seedlings after exposure to three temperatures for 10 days18℃(对照) (control) 22℃ 24℃ 26℃ 起始平均重量/g initial averaged weight 0.036±0.001 0.038±0.001 0.038±0.003 0.038±0.004 最终平均重量/g ultimate averaged weight 0.05±0.007 0.06±0.006* 0.069±0.006* 0.049±0.001 相对生长速率(重量)/%·d-1 relative growth rates in weights 3.85 4.53 5.99 2.59 起始平均最长叶长/cm initial averaged length of the longest leaves 1.88±0.08 1.76±0.05 1.86±0.13 1.67±0.11 最终平均最长叶长/cm ultimate averaged length of longest leaves 2.18±0.08 2.22±0.19 2.28±0.21 1.76±0.2* 相对生长速率(最长叶长)/%·d-1 relative growth rates in length 1.48 2.32 2.04 0.52 注:*. 与对照相比有显著差异(P < 0.05)
Note:*. statistically significant with the initial control (P < 0.05)2.2 光胁迫
2.2.1 室内消减的太阳光胁迫(70%太阳光,不含紫外线)
Fm′随光强增加而降低,上午9: 00~10: 00,最高光强320 μmol photons · m-2 · s-1,Fm′最低降到0.4左右(图 3)。随着光强降低,Fm′逐渐恢复,到16: 00时基本恢复到上午7: 00时的水平(图 4)。
![]() 图 3 室内水箱全天光照变化由QSPL-2100光度计测量获得Figure 3. Changes in light intensities above the indoor tanksMeasured by the QSPL-2100 photometer
图 3 室内水箱全天光照变化由QSPL-2100光度计测量获得Figure 3. Changes in light intensities above the indoor tanksMeasured by the QSPL-2100 photometer![]() 图 4 藻幼苗在图 3光照培养条件下即时荧光产量的变化(n=5)Figure 4. Fluorescence quantum yield measured in the presence of actinic light (Fm′) of S.horneri seedlings under reduced ambient solar irradiance as illustrated in Fig. 3
图 4 藻幼苗在图 3光照培养条件下即时荧光产量的变化(n=5)Figure 4. Fluorescence quantum yield measured in the presence of actinic light (Fm′) of S.horneri seedlings under reduced ambient solar irradiance as illustrated in Fig. 32.2.2 室外光胁迫
8: 30开始将铜藻幼苗暴露在室外水箱中仅1 h,Fv/Fm就降到0.36,随着光强和紫外强度的增强,Fv/Fm急剧下降,11: 30(最高1 490 μmol photons · m-2 · s-1和最高紫外光4 720 μW · cm-2)时降到0.11(图 5、图 6和图 7),仅为同一时间室内对照水箱的15%左右,随着光强和紫外光的降低稍有回升。经过24 h室内恢复后,处理1 h幼苗的Fv/Fm基本恢复到对照水平,超过1 h处理幼苗的Fv/Fm虽都有不同程度的恢复(图 7),但是幼苗叶片明显萎焉。
![]() 图 5 室外水箱光照变化由SAMPO-LX1330光度计测量获得Figure 5. Changes in photosynthetic active radiation (PAR) above the outdoor tanksMeasured by the SAMPO-LX1330 photometer
图 5 室外水箱光照变化由SAMPO-LX1330光度计测量获得Figure 5. Changes in photosynthetic active radiation (PAR) above the outdoor tanksMeasured by the SAMPO-LX1330 photometer![]() 图 6 室外水箱紫外线强度变化由TN-2340紫外线强度计测量获得Figure 6. Changes in UV radiation (280~380 nm) above the outdoor tanksMeasured by the TN-2340 UV light meter
图 6 室外水箱紫外线强度变化由TN-2340紫外线强度计测量获得Figure 6. Changes in UV radiation (280~380 nm) above the outdoor tanksMeasured by the TN-2340 UV light meter![]() 图 7 暴露在室外水箱以及24 h室内恢复后铜藻幼苗的Fv/Fm以室内水箱同一时刻铜藻的Fv/Fm作对照 (n=5)Figure 7. Optimal chlorophyll fluorescence quantum yields (Fv/Fm)of seedlings of S.horneri after exposure to direct solar irradiation in outdoor tank and the corresponding recovered values after 24 h indoor cultureFv/Fm of S.horneri in indoor tank at the same time was used as the control (n=5)
图 7 暴露在室外水箱以及24 h室内恢复后铜藻幼苗的Fv/Fm以室内水箱同一时刻铜藻的Fv/Fm作对照 (n=5)Figure 7. Optimal chlorophyll fluorescence quantum yields (Fv/Fm)of seedlings of S.horneri after exposure to direct solar irradiation in outdoor tank and the corresponding recovered values after 24 h indoor cultureFv/Fm of S.horneri in indoor tank at the same time was used as the control (n=5)2.3 盐胁迫
2.3.1 高盐胁迫
随着盐度增加,处理时间的延长,幼苗的Fv/Fm明显降低,24 h恢复后,即使在盐度60处理6 h的Fv/Fm也恢复到正常水平(P>0.05)(表 4)。藻体色泽无变化。表明幼苗能够耐受较高渗透压变化。
表 4 3个盐度条件下处理0.5、1, 3和6 h以及恢复24 h的铜藻的Fv/Fm(n=5)Table 4. Optimal chlorophyll fluorescence quantum yield (Fv/Fm) of seedlings of S.horneri after exposure to three different salinities for 0.5, 1, 3 and 6 h, respectively, and the recovered Fv/Fm after 24 hMeans±SD 盐度salinity 处理treatment 0.5 h 1 h 3 h 6 h 30 处理后测量 0.75±0.01 0.74±0.01 0.73±0.01 0.73±0.01 修复24 h后 0.72±0.01 0.72±0.02 0.72±0.01 0.72±0.01 45 处理后测量 0.69±0.02 0.68±0.02 0.68±0.01 0.68±0.02 修复24 h后 0.73±0.04 0.73±0.01 0.74±0.02 0.70±0.01 60 处理后测量 0.64±0.04* 0.7±0.02 0.66±0.02 0.61±0.01* 修复24 h后 0.72±0.01 0.72±0.02 0.69±0.02 0.68±0.02 注:*. 与对照相比有显著差异(P < 0.05)
Note:*. statistically significant with the initial control (P < 0.05)2.3.2 低盐胁迫
随着盐度降低和处理时间的延长,幼苗的Fv/Fm均呈下降趋势。盐度0处理0.5,1,3和6 h的Fv/Fm分别为对照组的95.77%,78.38%,50.00%和36.49%,经24 h恢复后藻体的Fv/Fm分别为对照组的97.18%,76.71%,44.44%和28.38%,因此,即使在淡水中处理1 h,幼苗的Fv/Fm基本可恢复(P>0.05),幼苗依然健康,色泽无变化;盐度9和21在6 h以内处理以及恢复的Fv/Fm分别为对照组的98.65%和90.54%,和对照组相比虽然有所降低,但不明显(P>0.05)(表 5)。结果表明,渗透压的降低对铜藻的影响不大,与羊栖菜(Sargassum fusiforme)的结果相似,但羊栖菜在淡水中处理6 h,Fv/Fm也可基本完全恢复[6]。
表 5 4种盐度条件下处理0.5, 1, 3和6 h以及24 h恢复后的铜藻幼苗的Fv/Fm(n=5)Table 5. Optimal chlorophyll fluorescence quantum yield (Fv/Fm) of seedlings of S.horneri after exposure to four salinities for 0.5, 1, 3 and 6 h, respectively, and the recovered Fv/Fm after 24 hMeans±SD 盐度salinity 处理treatment 0.5 h 1 h 3 h 6 h 30 处理后测量 0.72±0.01 0.74±0.01 0.72±0.01 0.74±0.01 修复24 h后 0.71±0.01 0.73±0.03 0.72±0.02 0.74±0.01 21 处理后测量 0.71±0.01 0.74±0.03 0.72±0.02 0.73±0.01 修复24 h后 0.71±0.01 0.73±0.02 0.73±0.01 0.67±0.02 9 处理后测量 0.71±0.01 0.71±0.02 0.68±0.03 0.67±0.02 修复24 h后 0.71±0.01 0.72±0.02 0.66±0.06* 0.60±0.02* 0 处理后测量 0.68±0.01 0.58±0.01* 0.36±0.07* 0.27±0.20* 修复24 h后 0.69±0.01 0.56±0.01* 0.32±0.10* 0.21±0.10* 注:*. 与对照相比有显著差异(P < 0.05)
Note:*. statistically significant with the initial control (P < 0.05)3. 讨论
叶绿素荧光作为光合作用能量转换的天然探针,含有极其丰富的光合作用信息,因易受逆境胁迫影响,广泛用于研究和探测逆境对植物光合作用生理的影响[7-8]。Fv/Fm表示PSII最大光量子产量,即PSII原初光能转换效率,是衡量PSII完整性的指标,SCHANSKER等[9]发现,当Fv/Fm大于0.44时,PSII活性随着Fv/Fm的降低而降低,当小于0.44时就完全失去活性,表现出反应中心的破坏。PSII反应中心失活分为PSII反应中心可逆失活和PSII反应中心遭到破坏[10]。
通过测量胁迫环境条件下铜藻幼苗的叶绿素荧光产量的变化,结果表明,35℃的高温在1 h已经对铜藻的PSII造成了不可逆转的损伤,幼苗可耐受的温度上限为28℃,不耐较强室外光胁迫,此试验最高1 490 μmol photons · m-2 · s-1和最高紫外光4 720 μW · cm-2已经对铜藻造成致命伤害,室内消减的太阳光320 μmol photons · m-2 · s-1虽然能对铜藻造成光抑制,但可恢复。铜藻具有较强耐受高盐(盐度60处理6 h)和淡水能力(盐度0处理1 h)。同为马尾藻属、马尾藻科,羊栖菜却能耐受40℃的高温长达6 h,室外强光胁迫(1 800 μmol photons · m-2 · s-1)和低盐胁迫(淡水6 h)[6]。此试验的环境因子都是影响海洋藻类生长的关键因素。
目前,国内外对高等植物生理胁迫的研究较多,但对大型海藻的研究较少[7, 11-12]。在高等植物中,有关高温胁迫机制的研究较为成熟。大量研究表明,PSII是最易热敏感的色素蛋白复合体之一[13-14],其活性在高温时受到抑制。还有报道称,类囊体膜对高温非常敏感,高温也会导致类囊体膜结构的改变,此种改变是由D1蛋白结构的改变所导致,高温使LHCII(捕光色素蛋白复合体II)与PSII反映中心脱离,与高等植物相比,大型藻类中含有藻体蛋白,最复杂的藻体蛋白位于类囊体膜的表面。高温会导致此类大型藻类PSII失活,并且使藻胆体的(PBS)从PSII反应中心脱离出来,还会导致藻胆蛋白降解[15]。由于Fv/Fm反映的是暗适应状态下光系统II的量子产量,强光导致Fv/Fm的不可逆说明,光抑制直接作用于光系统II的反应中心(包括天线色素)。照光后光系统II的量子产量降低,不仅是因为部分光系统II反应中心关闭,也是因为热耗散的增加,长时间暴露在室外光下幼苗的Fv/Fm明显降低,说明PSII光合能力下降,几乎所有吸收来的光能都被转化为热能,而没被转化为光合能[7, 16]。铜藻幼苗受室内消减太阳光可逆抑制的能力与海带(Laminaria japonica)、孔石莼(Ulva pertusa)、掌状红皮藻(Palmaria palmata)相同。
在此项研究中,铜藻幼苗最适生长温度为22~24℃,结果符合自然种群幼苗出现时间基本在夏季早期的观察[2]。由于铜藻的商业化栽培或者大规模重建铜藻藻场还没有实现,相关研究基本是空白,此项研究的结果提供了有价值的科学数据。另外,附生杂藻的去除是海藻人工栽培过程中一大难题。附生杂藻同要培养的海藻竞争生存空间、营养和光照[6]。在海带苗种培育过程中,每天都要用大量的海水冲洗苗帘来去除附生杂藻。在紫菜(Porphyra spp.)养殖中,随着潮汐变化,苗网每天都会干出一段时间,杂藻从而得到有效抑制(生产实践)。在铜藻人工苗种的人工培育过程中,根据此项研究可以采用自来水短暂浸泡的方法来有效地消除一些对渗透压敏感的杂藻种类,如石莼属(Ulva spp.)、多管藻属(Polysiphonia spp.)和水云属(Ectocarpus spp.)中的种类。采用叶绿素荧光技术检测海藻光合生理变化的最大优点是快速、简便、灵敏、以及对受测试的藻体无损伤[6],可以预测,该种技术将被广泛地应用在海藻人工栽培的研究中。
-
![]()
图 1 对虾-鲻围网分隔混养现场效果图
Figure 1. Real scene shooting of mesh enclosure isolated white shrimp-common mullet pond
![]()
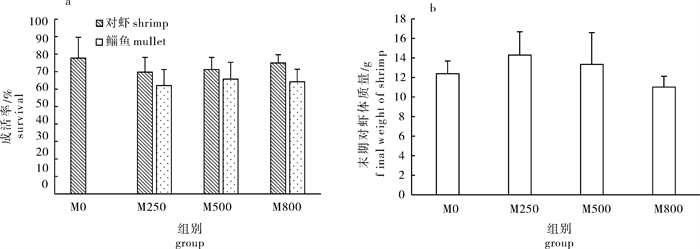
图 2 养殖结束后鱼、虾成活率(a)和对虾体质量(b)
Figure 2. Survival of shrimps and mullets (a)and final weight of shrimps (b)
![]()
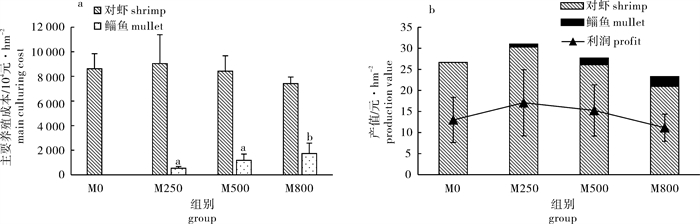
图 4 鱼、虾养殖产量(a)和产值(b)
各处理组间不同字母标注表示差异显著(P < 0.05)
Figure 4. Farming yield (a) and production value (b) of shrimps and mullets
Values with different superscripts are significantly different (P < 0.05).
![]()
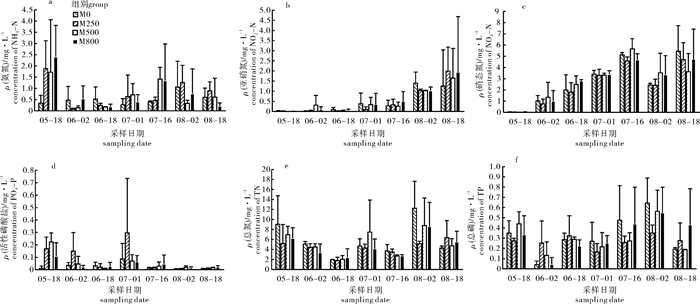
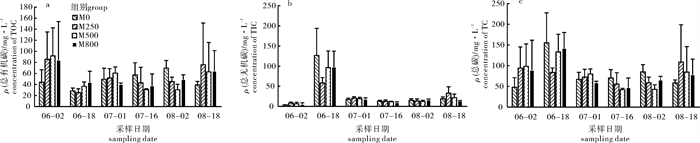
图 5 水体中氨氮(a)、亚硝态氮(b)、硝态氮(c)、活性磷酸盐(d)、总氮(e)和总磷(f)质量浓度
Figure 5. Concentrations of ammonium (a), nitrite (b), nitrate (c), dissolved inorganic phosphorus (d), total inorganic nitrogen (e) and total inorganic phosphorus (f) in water
![]()
图 6 水体中总有机碳(a)、总无机碳(b)、总碳(c)质量浓度
Figure 6. Concentrations of total organic carbon (a), total inorganic carbon (b) and total carbon (c) in water
![]()
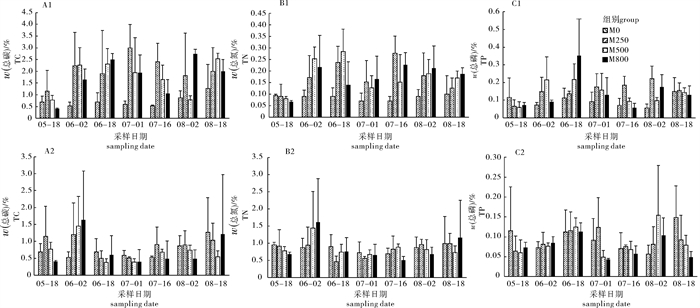
图 7 围网内、外沉积物总碳(内:A1,外:A2),总氮(内:B1,外:B2),总磷(内:C1,外:C2)浓度
Figure 7. Contents of total carbon (inside: A1, outside: A2), total nitrogen (inside: B1, outside: B2) and total phosphorus (inside: C1, outside: C2) of mesh enclosure
表 1 各处理组放苗及收获时间
Table 1 Stocking time and harvest time for different treatments
处理组
treatment池塘
pond No.对虾 L.vannamei 鲻 M.cephalus 放苗日期
stocking time收获日期
harvest time养殖周期/d
culture period放苗日期
stocking time收获日期
harvest time养殖周期/d
culture periodM0 504 2012-05-18 2012-09-05 110 ― ― ― 704 2012-05-18 2012-09-12 117 ― ― ― 1 205 2012-05-18 2012-09-01 106 ― ― ― M250 204 2012-05-18 2012-09-04 109 2012-05-18 2012-09-04 109 805 2012-05-28 2012-09-06 101 2012-05-26 2012-09-06 103 1 109 2012-05-23 2012-09-07 107 2012-05-30 2012-09-07 100 M500 705 2012-05-18 2012-08-29 103 2012-05-21 2012-08-29 101 1 204 2012-05-28 2012-08-19 83 2012-06-01 2012-08-19 80 104 2012-05-18 2012-09-15 120 2012-05-18 2012-09-15 120 M800 1 111 2012-05-23 2012-09-06 106 2012-05-30 2012-09-06 99 706 2012-05-18 2012-08-20 94 2012-05-20 2012-08-20 92 103 2012-05-18 2012-08-11 85 2012-05-18 2012-08-11 85  下载: 导出CSV
下载: 导出CSV
-
[1] DASTIDAR P G, MALLIK A, MANDAL N. Contribution of shrimp disease research to the development of the shrimp aquaculture industry: an analysis of the research and innovation structure across the countries [J]. Scientometrics, 2013, 97(3): 659-674. doi: 10.1007/s11192-013-0977-9
[2] 彭卫正, 张建强, 郑真龙, 等. 珠三角地区南美白对虾养殖的主要病害防治及养殖管理建议[J]. 广东饲料, 2012 (4): 47-48. doi: 10.3969/j.issn.1005-8613.2012.04.021 [3] 吕华当. 寸虾"偷死" 潮袭击珠三角[J]. 海洋与渔业, 2011(7): 22-23. doi: 10.3969/j.issn.1672-4046.2011.07.014 [4] 邱丽华, 吴进锋, 张汉华, 等. 海水池塘鲻鱼, 斑节对虾混养的初步研究[J]. 湛江海洋大学学报, 2000, 20(4): 69-71. doi: 10.3969/j.issn.1673-9159.2000.04.015 [5] LUPATSCH I, KATZ T, ANEGL D L. Assessment of the removal efficiency of fish farm effluents by grey mullets: a nutritional approach [J]. Aquac Res, 2003, 34(15): 1367-1377. doi: 10.1111/j.1365-2109.2003.00954.x
[6] 孔谦. 凡纳滨对虾与鲻鱼混养中精养池的理化生物因子的研究[D]. 湛江: 广东海洋大学, 2010. 10.7666/d.y1804038 [7] 雷衍之. 养殖水环境化学实验[M]. 北京: 中国农业出版社, 2006: 31-60. [8] 杨柳燕, 赵兴青, 肖琳, 等. 沉积物总氮、总磷联合测定分析方法: 中国, CN1869656 [P]. 2006-11-29. https://wenku.baidu.com/view/2de246fa5d0e7cd184254b35eefdc8d377ee14b1?fr=xueshu [9] 查广才, 周昌清, 黄建荣, 等. 低盐度养殖的凡纳滨对虾体长和体重的增长规律[J]. 水产学报, 2006, 30(4): 489-494. doi: 10.3321/j.issn:1000-0615.2006.04.009 [10] 刘少英, 朱长波, 文国樑, 等. 凡纳滨对虾在半集约化土池养殖模式下的生长特性分析[J]. 广东农业科学, 2012, 39(9): 9-13. doi: 10.3969/j.issn.1004-874X.2012.09.003 [11] 虞为, 李卓佳, 王丽花, 等. 对虾单养养和对虾-罗非鱼混养试验围隔水质动态及产出效果的对比[J]. 中国渔业质量与标准, 2013, 3(2): 89-97. http://www.aquaticjournal.com/article/id/655f7b96-4c8f-40bc-bb43-068a545669d2 [12] 裴宇. 凡纳滨对虾与金钱鱼混养对池塘生态因子及能量收支的影响[D]. 湛江: 广东海洋大学, 2012. 10.7666/d.Y2215296 [13] 胡伟国. 南美白对虾与斑点叉尾混养试验[J]. 科学养鱼, 2008 (8): 38-38. https://wenku.baidu.com/view/7916cd38bd64783e09122bc0?fr=xueshu [14] 郭泽雄. 高位池南美白对虾与鲻鱼混养技术初探[J]. 科学养鱼, 2004 (3): 32-33. http://qikan.cqvip.com/Qikan/Article/Detail?id=9303578 [15] JOYNI M J, KURUP BM, AVNIMELECH Y. Bioturbation as a possible means for increasing production and improving pond soil characteristics in shrimp-fish brackish water ponds [J]. Aquaculture, 2011, 318(3/4): 464-470. doi: 10.1016/j.aquaculture.2011.05.019
[16] PHAN-VAN M, ROUSSEAU D, PAUW N D. Effects of fish bioturbation on the vertical distribution of water temperature and dissolved oxygen in a fish culture-integrated waste stabilization pond system in Vietnam [J]. Aquaculture, 2008, 281(1): 28-33. https://www.sciencedirect.com/science/article/pii/S0044848608003098
[17] 杨大佐. 刺参养殖池塘有机碳周年变化的初步研究[J]. 科技致富向导, 2011 (20): 108-109. https://www.nstl.gov.cn/paper_detail.html?id=4bb9130572dafc65e48041d4c792151a [18] 赵文, 董双林, 李德尚, 等. 盐碱池塘围隔生态系统的悬浮物结构及有机碳库储量[J]. 生态学报, 2002, 22(12): 2133-2140. doi: 10.3321/j.issn:1000-0933.2002.12.017 [19] 余根鼎, 江敏, 邢斌, 等. 凡纳滨对虾养殖塘主要有机污染指标相关性分析[J]. 淡水渔业, 2010, 40(2): 67-70. doi: 10.3969/j.issn.1000-6907.2010.02.013 [20] 游奎, 马甡. 对虾养殖生态系中有机碳的初步研究[J]. 青岛海洋大学学报: 自然科学版, 2002, 32(1): 51-55. doi: 10.3969/j.issn.1672-5174.2002.01.023 [21] 王丽花, 曹煜成, 李卓佳. 溶藻细菌控藻作用及其在对虾养殖池塘中的应用前景[J]. 南方水产科学, 2012, 8(4): 76-82. doi: 10.3969/j.issn.2095-0780.2012.04.012 [22] 彭聪聪, 李卓佳, 曹煜成, 等. 虾池浮游微藻与养殖水环境调控的研究概况[J]. 南方水产, 2010, 6(5): 74-80. doi: 10.3969/j.issn.1673-2227.2010.05.012 [23] 曹煜成, 李卓佳, 杨莺莺, 等. 浮游微藻生态调控技术在对虾养殖应用中的研究进展[J]. 南方水产, 2007, 3(4): 70-73. doi: 10.3969/j.issn.2095-0780.2007.04.012 [24] 张汉华, 李卓佳, 郭志勋, 等. 有益微生物对海水养虾池浮游生物生态特征的影响研究[J]. 南方水产, 2005, 1(2): 7-14. doi: 10.3969/j.issn.2095-0780.2005.02.002 [25] 张振东, 王芳, 董双林, 等. 草鱼、鲢鱼和凡纳滨对虾多元化养殖系统结构优化的研究[J]. 中国海洋大学学报: 自然科学版, 2011, 41(7/8): 60-66. doi: 10.3969/j.issn.1672-5174.2011.07.010 [26] MARCEL P M, LUIS R M C, MARCO P C, et al. Shrimp polyculture: a potentially profitable, sustainable, but uncommon aquacultural practice [J]. Rev Aquac, 2010, 2(2): 73-85. doi: 10.1111/j.1753-5131.2010.01023.x
[27] MUANGKEOW B, IKEJIMA K, POWTONGSOOK S, et al. Effects of white shrimp, Litopenaeus vannamei (Boone), and Nile tilapia, Oreochromis niloticus L. , stocking density on growth, nutrient conversion rate and economic return in integrated closed recirculation system[J]. Aquaculture, 2007, 269(1): 363-376. doi: 10.1016/j.aquaculture.2007.04.002
[28] JASPE C J, CAIPANG C M A, ELLE B J G. Polyculture of white shrimp, Litopenaeus vannamei and milkfish, Chanos chanos as a strategy for efficient utilization of natural food production in ponds [J]. Anim Biol Anim Husband, 2011, 3(2): 96-104. https://www.semanticscholar.org/paper/Polyculture-of-white-shrimp%2C-Litopenaeus-vannamei-a-Jaspe-Caipang/28d710e5c512a087138b67517b6b9efaab1107b0
[29] 董双林, 介子林, 赵夕旦, 等. 对虾与滤食性鱼类分隔式混合养殖的方法: 中国, CN1736182[P]. 2006-02-22. http://www.xjishu.com/zhuanli/01/200510044224.html [30] 赵裕青, 唐兴本, 潘海军. 池塘设隔离网混养南美白对虾与鲤鱼技术[J]. 科学养鱼, 2012 (2): 33-33. https://d.wanfangdata.com.cn/periodical/Ch9QZXJpb2RpY2FsQ0hJTmV3UzIwMjQxMTA1MTcxMzA0Eg1reHl5MjAxMjAyMDIwGghwcXptYjN0NA%3D%3D [31] SUN W M, DONG S L, JIE Z L, et al. The impact of net-isolated polyculture of tilapia (Oreochromis niloticus) on plankton community in saline-alkaline pond of shrimp (Penaeus vannamei) [J]. Aquac Int, 2011, 19(4): 779-788. doi: 10.1007/s10499-010-9394-8








计量
- 文章访问数: 3567
- HTML全文浏览量: 187
- PDF下载量: 1821
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
