
Acoustic monitoring on fish resources in Xijiang section of Pearl River during first closed fishing season
-
摘要:
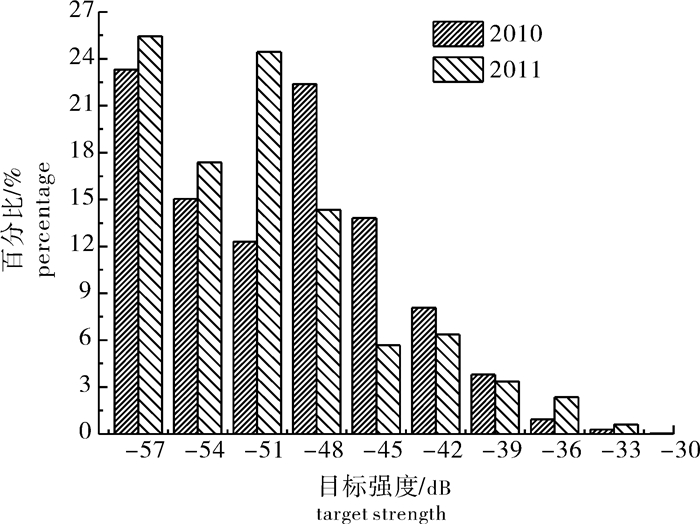
2011年珠江开始实行禁渔期制度。该研究采用Simrad EY60鱼探仪,分别于2010年5月19日和2011年5月22日对珠江西江段进行了走航式探测。结果显示,珠江禁渔期期间西江段鱼类资源密度增加,2010年和2011年鱼类平均密度分别为0.051 3尾·m-3和0.106 8尾·m-3,根据渔获物调查数据,推算增幅分别为广东鲂(Megalobrama terminalis)47.85%、赤眼鳟(Squaliobarbus curriculus)18.54%、鲮(Cirrhina moitorella)10.87%和鳊(Parabramis pekinensis)6.25%;两年西江段深水区域鱼类密度均较大,尤其是水底结构为凹形区域,鱼类有明显的聚集现象;浅水区域且底部较平整的水域,鱼类分布分散且密度较低;垂直分布方面,鱼类主要分布4~10 m中下层水层。珠江禁渔期的实施,在一定程度上保护了鱼类的产卵亲体繁殖,对鱼类资源数量的补充起到了增殖作用,同时在很大程度上护养了鱼类的自然生长,起到了资源增重作用。
Abstract:Closed fishing season was initially implemented in Pearl River in 2011. We used a Simrad EY60 echo-sounder with 120 kHz split-beam transducer to survey the Xijiang section of Pearl River. The results show that during the closed season, the density of fish resources was increasing, and the average fish densities were 0.051 3 ind·m-3 and 0.106 8 ind·m-3 in 2010 and 2011, respectively. According to the data of catches, the increases were deduced as follows: Megalobrama terminalis 47.85%, Squaliobarbus curriculus 18.54%, Cirrhina moitorella 10.87% and Parabramis pekinensis 6.25%. The deep water had higher fish density than shallow water, especially that fish obviously aggregated in concave region. The fish mainly distributed at depth of 4~10 m. The implementation of closed fishing season, to some extent, protects breeding of spawning stocks and supplements fish resources as well as maintenances natural growth of fishes.
-
Keywords:
- fish resources /
- closed fishing /
- acoustic assessment /
- Pearl River
-
近年来,广西海水养殖业得到了长足发展,海水养殖业的快速发展不仅改善了人们的食物结构,丰富了城乡居民的菜篮子,而且对发展渔区经济,增加渔农民收入发挥了巨大的作用,已成为农业经济增长的一个新亮点。
1. 海水养殖业现状
广西海岸线长1 595 km,沿岸岛屿约700个,岛屿岸线长600余km。潮间带滩涂面积10万hm2,适宜水产养殖的面积约6.7万hm2。20 m等深线以内浅海面积65万hm2,适宜水产养殖的面积约57万hm2。水域滩涂生物资源丰富,盛产对虾Penaeus、锯缘青蟹Scylla serrata、马氏珠母贝Pinctada martensii、近江牡蛎Crassostrea rivularis、文蛤Meretrix meretrix及赤点石斑鱼Epinephelus akaara、真鲷Pagrosomus major、二长棘鲷Parargyrops edita、银鲳Pampus argenteus等主要经济种类,发展海水养殖业条件十分优越。从20世纪90年代开始,在“以养为主”的方针指导下,实施包括旨在发展海水养殖在内的“种子工程”和“万水工程”等经济战略下,坚持面积、产量、质量和效益并重,海水养殖业得到快速、稳步发展。
1.1 养殖规模不断扩大
2004年,全区海水养殖面积达6万多hm2,是1994年的3.6倍。其中陆基池塘养殖约2万hm2,滩涂养殖3万多hm2,占近期可开发滩涂总面积的45%;浅海开发约1万hm2,占适宜开发利用面积的3%。充分发挥资源优势,潮上带、潮间带和浅海开发同步发展,养殖热潮有增无减。大批养殖户通过发展养殖生产走上了致富之路。
1.2 养殖品种呈多元化发展
为适应市场需要,在稳定常规品种养殖的同时,扩大名优品种养殖。养殖品种发展到30多个,主要有对虾(长毛对虾Penaeus penicillatus、斑节对虾P.monodon、日本对虾P.japonicus、南美白对虾P.vannamei)、文蛤、近江牡蛎、马氏珠母贝、泥蚶Tegillarca granosa、海湾扇贝Argopecten irradians、栉江珧Pinna pectinata Linnaeus、大獭蛤Lutria maxima、中华乌塘鳢Bostrichthys sinensis、大弹涂鱼Boleophthalmus pectinirostris、真鲷、美国红鱼Sciaemops ocellatus、赤点石斑鱼、红鳍笛鲷Lutjanus erythropterus、真鲷、黄鳍鲷Sparus lctus、黑鲷S.macrocephalus、鲈鱼Lateolabrax japonicus、军曹鱼Rachycentron canadum、鲻鱼Mugil cephalus等。养殖品种的多样化和优质化丰富了消费市场,满足了人们生活需求。
1.3 养殖模式向高产高效方向发展
养殖模式主要推广混养、轮养、立体吊养、网箱养殖、深水礁体养殖以及对虾高位池养殖、珍珠深水育珠技术、贝类增养殖等养殖模式。海水养殖由岸边、滩涂养殖向浅海养殖推进,由单一养殖向立体养殖的深度和广度发展,逐渐形成鱼、虾、蟹贝全面发展的格局。
1.4 养殖向规模化、基地化、专业化发展
形成了以对虾、珍珠、近江牡蛎、文蛤、泥蚶、名贵鱼类6大主导产品的海水养殖基地。1.3 hm2以上的规模养殖户达712户,面积6 200 hm2,占养殖总面积的10%;其中6.7 hm2以上的规模养殖户有75户。北海工厂化养殖基地面积已达30万m2,10 m等深线内深水养殖大獭蛤、栉江珧等面积达4 000 hm2;钦州建成了广西最大的近江牡蛎养殖基地,其中七十二泾近江牡蛎浮筏串式高密度养殖近1 500 hm2,形成集观光、生产于一体的海洋牧场。
1.5 种苗生产数量增长迅猛
2004年育出各种鱼、虾110亿尾,比1994年增长了9倍。对虾育苗场近100家,育苗水体达10万m3,年育苗量109亿尾,完全能满足广西对虾养殖生产的需要。近江牡蛎自然采苗2.5亿只,不但满足广西养殖需要而且部分苗种运销区外。
2. 存在问题
2.1 养殖水环境污染日趋严重
随着沿海地区经济的迅速发展,城市毗连的海域、港口、海湾、河流入海口,岸线海域的水质已受到不同程度的污染,并有继续发展的趋势。工业废水、城市生活污水和大量富含有机质、无机氮和磷及有机农药的农业污水随泾流进入近海水域,致使养殖水质恶化,严重地影响养殖种类的生存和生长。另一方面是养殖自身水环境的污染,养殖过程中的残饵、化肥、消毒药品等,特别是残饵中的蛋白质、氮和磷未经科学处理就直接排放到海区水环境中,易引起海水的富营养化,滋生病原;大量新增加的养殖设施使养殖区及其毗邻水域场发生改变,养殖设施的屏障效应使流速降低,影响了营养物质的输入和污物的输出,使陆源污染物得不到及时的稀释扩散,滞留在养殖水域。
2.2 养殖资源开发得不到有效控制
渔业管理虽然推行了养殖证制度,但养殖开发利用仍然处于无序状态。养殖水域滩涂规划滞后,养殖规模的盲目扩大与海区养殖容量相对有限的问题仍严重。目前海水养殖规模大小大都是人为盲目决定,或由市场单一引导,缺乏有科学依据的政策性宏观调控,养殖生产往住一哄而上。近江牡蛎、马氏珠母贝养殖近年来连续发生死亡,2005年早造的对虾养殖发病率超过五成。海上近江牡蛎筏吊养殖过密,也将直接危害到一些名贵物种的繁衍生长,发展无序化的弊病,已显端倪。
2.3 养殖品种种质退化[1]
近年来,水产种苗培育偏重数量、不讲质量;不重视选育,由于近亲繁殖,种质退化严重,导致养殖性能严重衰退,生长减缓,个体小型化,抗逆性减弱。缺少高产抗逆的优良品种和高产值养殖对象,优质种总量比例小,产量、质量提高缓慢。
2.4 局部区间养殖品种结构失衡
在不同生态类型海区养殖中,普遍存在养殖种类结构不合理现象,池塘对虾、滩涂贝类养殖中,养殖种类或类群单一问题由来已久。养殖生物量严重超过环境负荷,进行掠夺式养殖。结构单一的密集养殖,使生态系统能量和物质由于超支而贫乏,造成循环过程紊乱和生态失衡,致使某些污损、赤潮和病原生物异常发生,极易引发病害的发生和流行。
2.5 养殖病害爆发、流行日趋严重
虾病流行,贝类发病减产,形势严峻。暴发性、流行性疾病频繁发生,病毒性疾病类型愈来愈多,非病原性疾病愈来愈突出,病原耐药性越来越强,水产养殖病害不断发生、蔓延。养殖生态、疾病与环境和营养的关系研究不够,短期内难以提出普遍有效的技术措施,使生产难以持续增长。
2.6 养殖科技含量偏低
从业者养殖技术参差不齐,科技应用水平偏低。科研手段落后,科技不能满足生产快速发展的要求,许多领域滞后于生产。在水产养殖发展前途的重大关键技术方面,科技的先导作用不能充分发挥。良种引进更新换代步伐不快,科研所必需的仪器设备不能及时配置,致使试验手段无法更新,长期处于落后状态等等,影响了科学研究的深度和精度。
2.7 养殖产品质量亟待提高
国际、国内消费市场对水产品的质量安全性都有了很高的认识和要求,而目前的养殖生产环境、产品质量标准、投入品及药残监控等方面与国际市场的要求尚有一定差距,提高养殖产品的食用安全性、使产品更好地进入国内外市场显得格为迫切。
2.8 渔业基础资料缺乏
由于科研经费的缺乏,制约了渔业资源和渔业生态环境的基础性调查。缺乏常规性、基础性的调查和监测数据,渔业统计资料不规范、不完整,严重影响了渔业决策的科学依据和理论基础,致使生产带有一定的盲目性,难以科学、合理、有效地利用海洋渔业资源。
3. 海水养殖业可持续发展的对策
3.1 加强养殖环境整治,树立环境保护与持续发展观念
加强海洋环境的保护,要用法律手段来控制和治理养殖水域污染,处理好养殖发展与保护资源和生态环境的关系,保证养殖资源的合理和永续利用。对于陆源污染的治理,要重点放在污染形成的前端,增加污水和废渣的处理能力,控制污染物入海量。要用法律手段对重点排污河流、临海城市和直接排污入海的大、中企业进行重点管理,规定其污水处理率和排放达标率,以控制工业废水、生活污水等的排放,要改变人们长期以来形成的,只知开发利用、不知保护海洋的观念。调整优化养殖结构,改进养殖技术,提高饲料质量,科学养殖,养殖污水经过净化处理后再排放等措施,把水产养殖自身污染降低到最低程度。
3.2 加强养殖容量研究,合理地布局养殖结构[1, 2]
加强对养殖容量研究,科学地确定各养殖品种的养殖负荷量,制订科学的养殖规划、养殖布局,确定适宜合理的养殖规模。调整现有的区域结构、品种结构和养殖方式等。调整养殖布局,将过度密集的近岸养殖转移到外海,大力发展深水抗风浪网箱及深水养鱼。把以前在潮间带及水面上进行的养殖转移到水面以下,发展海珍品底播养殖及沉箱养殖。合理搭配养殖品种,实施立体生态养殖,提高水域利用率。营造海底森林,大力发展海洋牧场、增殖渔业、人工鱼礁、围海养殖,形成深海海洋牧场产业带、外海抗风浪网箱养殖产业带、近海筏式养殖产业带、潮间带海珍品养殖产业带、滩涂池塘养殖产业带、观光及休闲渔业产业带、陆上工厂化养殖产业带等优势产业带,拓展新的发展空间,增加新的发展活力。
3.3 实施良种工程,实现养殖对象良种化[3]
加快水产原良种繁育基地建设,完善水产苗种供应体系。做好种苗提纯复壮,培养性状好、抗病力强的品种,实现养殖良种化。通过引进、繁育、提纯、复壮,开发一大批生产性状优良、经济价值较高的名优新品种。增加名特优新品种比重,从而调整、优化养殖品种结构。抓好苗种生产许可证的年审和新证换发工作,并加大水产苗种生产及流通的监管和执法检查力度,促使苗种生产和销售健康、有序地进行。
3.4 贯彻以防为主的原则,加强病害防治体系建设
随着水产养殖向高密度、高产方向发展,防治病害将越来越重要,要重视和加强病害防治工作。贯彻以防为主的原则,科学养殖,减少病害发生。应从长远来规划水产养殖的发展基地,制定相应的环保措施。组织科研力量,对重大流行病实行多单位联合攻关,提高科技应变能力,满足生产需要。健全基层水产科技推广网络,发挥基层科技人员的作用,尤其要强化镇级技术干部的培训,发挥其骨干作用。做好种苗的检疫及渔用药物的监检工作,严格把关;积极研究和探索生态防治技术,从水质调控人手,防治水产动物的疾病。
3.5 实施科技兴渔战略,提高养殖业综合水平[4]
利用国内外现有科研成果,迅速转化为生产力,提高水产养殖的科技含量。依靠科技进步,推广优良品种和先进养殖技术。加强技术培训,开展多种形式的养殖技术培训班、现场调查会、交流会,提高养殖业从业人员的养殖技术。
3.6 加强生产环境控制和监督管理,建立完整的质量保证体系
加强对苗种、饲料、渔药等生产投入品的监督管理,确保投入品的质量。推广健康养殖技术,推进无公害水产养殖基地建设。建立健全渔业质量标准体系,积极推行HACCP、欧盟等国际通行的水产品质量标准。抓好无公害养殖技术推广,全面推行无公害生产。逐步推行养殖场注册、养殖生产日志、渔药使用记录、产品标识、苗种检疫、环境监测、药残抽捡等制度。
3.7 加强渔业基础理论研究,为合理开发养殖资源提供依据
加强渔业资源调查和监测及渔业环境监测,制订科学的渔业规划、渔业布局,确定适宜合理的渔业规模,走科技兴渔和科技管理结合的道路,做到渔业资源与海区水交换能力、渔业作业模式和技术与品种结构间的平衡协调发展,实现海洋渔业资源的可持续开发利用。
3.8 实施依法治渔,严格规范养殖生产行为
加大渔业法制建设,认真贯彻《渔业法》及涉渔法律法规,大力推行养殖许可证制度、苗种生产许可证制度,《水产品质量管理条例》等法规及标准。组织编制“水产养殖发展规划”、“养殖区划布局规划”,对养殖生产进行宏观调控和指导。水产养殖的立项上马建立严格的审批制度,养殖项目要进行“生态成本”的核算,做到未雨绸缪,达到经济发展、资源节约与环境保护的和谐和统一。同时加强对从业人员的岗位技术培训,制订水产养殖的准入制度、持证生产制度和苗种检疫制度等,走绿色养殖之路,走法制化的水产养殖之路。
-
![]()
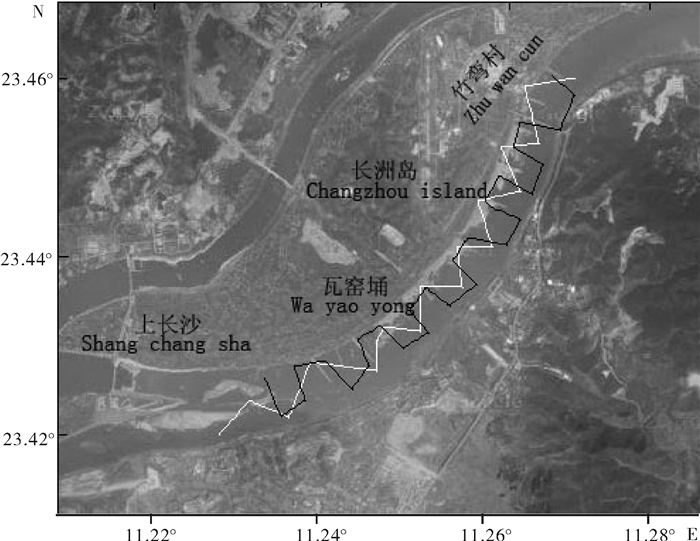
图 1 珠江西江声学调查区域
白色线为2010年航迹线,黑色线为2011年航迹线。
Figure 1. Map of acoustic survey in Pearl River
White and black lines are the track lines of 2010 and 2011, respectively.
![]()
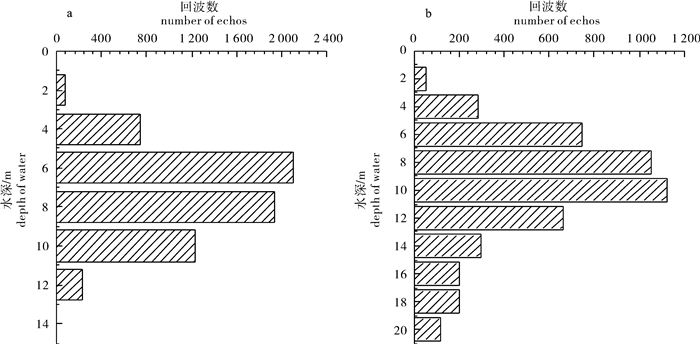
图 2 2010年(a)和2011年(b)探测区域鱼类的垂直分布
Figure 2. Vertical distribution of investigated areas in 2010 (a) and 2011 (b)
表 1 2010年和2011年主要渔获物信息
Table 1 Catches in 2010 and 2011
种类
species体质量/g
weight体长/cm
length体质量百分比/%
percentage of weight数量百分比/%
percentage of quantity2010 2011 2010 2011 2010 2011 2010 2011 广东鲂(M.terminalis) 359.56±139.68 466.33±193.71 25.72±3.69 28.78±18.85 44.41 50.10 12.54 47.90 赤眼鳟(S.curriculus) 108.03±70.37 325.00±163.21 17.56±4.06 25.93±4.85 17.18 21.71 23.68 28.63 鲮(C.moitorella) 172.23±137.72 453.85±205.02 18.23±3.51 27.09±3.49 10.05 18.37 16.87 15.08 鳊(P.pekinensis) 174.00±65.84 233.14±60.41 21.80±2.88 26.77±3.56 5.83 2.62 7.12 5.34 平均 average 334.37 405.90 19.07 27.48 - - - -  下载: 导出CSV
下载: 导出CSV
-
[1] 郑慈英. 珠江鱼类志[M]. 北京: 科技出版社, 1989: 389-398. https://www.las.ac.cn/front/book/detail?id=c1348fb22064fd8b05d40e92a8abfcf7 [2] 陆奎贤. 珠江水系渔业资源[M]. 广州: 广东科技出版社, 1990: 94-121. https://xueshu.baidu.com/usercenter/paper/show?paperid=e6239447508cf703129531efd4d95fc0 [3] 潘炯华. 广东淡水鱼类志[M]. 广州: 广东科技出版社, 1991: 11-19. https://www.gdstp.com.cn/Latest/book_1164.html [4] 王丹, 赵亚辉, 张春光, 等. 广西野生淡水鱼类的物种多样性及其资源的可持续利用[J]. 动物分类学报, 2007, 32(1): 160-173. doi: 10.3969/j.issn.1000-0739.2007.01.029 [5] 朱书礼, 李新辉, 李跃飞, 等. 西江广东肇庆段赤眼鳟的年龄鉴定及生长研究[J]. 南方水产科学, 2013, 9(2): 27-31. doi: 10.3969/j.issn.2095-0780.2013.02.005 [6] 李跃, 李新辉, 谭细畅, 等. 西江肇庆江段渔业资源现状及其变化[J]. 水利渔业, 2008, 28(2) : 80-83. https://kns.cnki.net/kcms2/article/abstract?v=WStw-PbchozE79ndqh0PqRk7wh975fjiaffZPxCfElp_1k7dVJsJz5e6wzbqA7lj81rpyyOg5YpHwg2MbJKhMQ12z6Y-qdYQzjcCb2ayv3P6TTTUM8Pb-pFFNcL6GB_5sJXLuyYzxc3qiCmRWsreoKbGdMv_MckGISiJ5WFGnKNoIgbDKNK49WlXnL0bZ7rx&uniplatform=NZKPT&language=CHS [7] 李捷, 李新辉, 谭细畅, 等. 广东肇庆西江珍稀鱼类省级自然保护区鱼类多样性[J]. 湖泊科学, 2009, 21(4) : 556-562. doi: 10.3321/j.issn:1003-5427.2009.04.015 [8] 李捷, 李新辉, 贾晓平, 等. 西江鱼类群落多样性及其演变[J]. 中国水产科学, 2010, 17(2): 298-311. http://www.fishscichina.com/zgsckx/article/abstract/1016?st=advanced_search [9] 谭细畅, 李新辉, 陶江平, 等. 西江肇庆江段鱼类早期资源时空分布特征研究[J]. 淡水渔业, 2007, 37(4): 37-40. doi: 10.3969/j.issn.1000-6907.2007.04.008 [10] 谭细畅, 李新辉, 林建志, 等. 基于水声学探测的两个广东鲂产卵群体繁殖生态的差异性[J]. 生态学报, 2009, 29(4): 1756-1762. doi: 10.3321/j.issn:1000-0933.2009.04.016 [11] 陈春亮, 曲念东, 侯秀琼, 等. 2007年伏季休渔深圳海域渔业资源调查分析[J]. 水产科学, 2008, 27(12): 648-651. doi: 10.3969/j.issn.1003-1111.2008.12.011 [12] 程家骅, 刘子藩. 东海区伏季休渔渔业生态效果的分析研究[J]. 中国水产科学, 1999, 6(4): 81-85. http://www.fishscichina.com/zgsckx/article/abstract/3171?st=advanced_search [13] 段辛斌, 刘绍平, 熊飞, 等. 长江上游干流春季禁渔前后三年渔获物结构和生物多样性分析[J]. 长江流域资源与环境, 2008, 17(6): 878-885. doi: 10.3969/j.issn.1004-8227.2008.06.010 [14] 谭细畅, 陶江平, 李新辉, 等. 回声探测仪在我国内陆水体鱼类资源调查中的初步应用[J]. 渔业现代化, 2009, 36(3): 60-64. doi: 10.3969/j.issn.1007-9580.2009.03.014 [15] 赵宪勇, 陈毓桢. 狭鳕(Theragra chalcogramma Pallas)目标强度的现场测定[J]. 中国水产科学, 1996, 2(4): 19-27. http://www.fishscichina.com/zgsckx/article/abstract/4306?st=advanced_search [16] PELTONEN H, BALK H. The acoustic target strength of herring (Clupea harengus L. ) in the northern Baltic Sea[J]. ICES J Mar Sci: Journal du Conseil, 2005, 62(4): 803-808. doi: 10.1016/j.icesjms.2005.02.001
[17] TORGERSEN T, KAARTVEDT S. In situ swimming behaviour of individual mesopelagic fishstudied by split-beam echo target tracking[J]. ICES J Mar Sci: Journal du Conseil, 2001, 58(1), 346-354. doi: 10.1006/jmsc.2000.1016
[18] FOOTE K G. Fish target strengths for use in echo integrator surveys[J]. J Acoust Soc Am, 1987, 82(3): 981. doi: 10.1121/1.395298
[19] TAN X C, MYOUNGHEE K, TAO J P. Hydroacoustic survey of fish density, spatial distribution, and behavior upstream and downstream of the Changzhou Dam on the Pearl River, China[J]. Fish Sci, 2011, 77(6): 891-901. doi: 10.1007/s12562-011-0400-5
[20] KIESER R, MULLIGAN T J. Analysis of echo counting data: a model[J]. Can J Fish Aquat Sci, 1984, 41(3): 451-458. doi: 10.1139/f84-054
[21] GERLOTTO F, FRÉON P. Some elements on vertical avoidance of fish schools to a vessel during acoustic surveys[J]. Fish Res, 1992, 14(4): 251-259. doi: 10.1016/0165-7836(92)90035-R
[22] MISUND O A, TOTLAND B, FLOEN S, et al. Computer based detection of fish schools by a multibeam sonar[C]//BJORNO L. Proc 2nd Eur Conf Underwater Acoustics, Conpenhagen, Denmark, 4-8 July, 1994. Amsterdam: Elservier, 1994: 815-820.
[23] 谭细畅, 李新辉, 陶江平, 等. 青皮塘产卵场广东鲂繁殖群体的水声学探测[J]. 生态学杂志, 2008, 27(5): 785-790. https://kns.cnki.net/kcms2/article/abstract?v=WStw-PbchoxD5ApYtOdQXmC5Q4hYcSoxHvZXPHecqsKM0WCYJ3391_2ae6Yumt7PFrydgGbdcd4Nv8nzoEAa5qftQTNb9sYerEP1x9EoqpNTRW4eUHA8Z8zV6OES3CZhEg0Y1IjB0RZLOsif8TElMa74zRkSo1fRsvnrd1MoQ8sahsonnZckze4esTW7WUAP&uniplatform=NZKPT&language=CHS -
期刊类型引用(17)
1. 汪洋,袁跃峰,苗晓君,范庆仁. 工厂化水产养殖自动投饲机下料仿真. 农村经济与科技. 2024(04): 68-72 .  百度学术
百度学术
2. 高炜鹏,谢永和,李德堂,王君. 基于CFD-DEM耦合的养殖工船自动投饲机的研制与实验. 饲料工业. 2024(10): 10-18 . 百度学术
3. 王龙宝,杨翰瑜,王庆杰,卢彩云,王超,何进. 小麦气流辅助直线投种装置设计与试验. 农业机械学报. 2024(06): 111-120 . 百度学术
4. 董建涛,陈传峰,杨蕾. 基于EDEM-Fluent耦合仿真的湿性混凝土颗粒气力清扫起动特性研究. 大众科技. 2023(04): 32-36 . 百度学术
5. 高炜鹏,谢永和,李德堂,王君,陈卿,洪永强,张佳奇. 养殖工船自动投饲机设计和螺旋下料器的仿真分析. 渔业现代化. 2023(04): 59-67 . 百度学术
6. 骆意,张方华,朱端祥,魏树辉,黄建伟,程晓夏. 深远海养殖设备投料选择器结构设计探讨. 机械工程师. 2023(12): 85-88 . 百度学术
7. 欧阳联格,汪仑,王会,王冠,黄飞,龙天明. 抽吸式地震救援机特大颗粒负压输送数值模拟. 震灾防御技术. 2023(04): 864-872 . 百度学术
8. 俞国燕,陈振雄,刘皞春,姬文超,张宏亮. 船载式投饲机饲料输送关键参数仿真分析. 渔业现代化. 2022(02): 10-17 . 百度学术
9. 丁乐声,陈潇,谢庆墨,严俊,曹南. 基于CFD-DEM的气力投饵分配器参数影响分析. 饲料研究. 2022(08): 118-122 . 百度学术
10. 张三丰,熊威,柯林,黄静林,沈学锋. 针对饲料气力输送系统设备选型参数的理论及仿真计算. 中国水运(下半月). 2022(08): 72-74 . 百度学术
11. 黄建伟,骆意,魏树辉,陈铭治,朱端祥,刘亮清. 深远海养殖自动投饲系统仿真分析与试验验证. 渔业现代化. 2022(05): 68-75 . 百度学术
12. 买买提明·艾尼,加合甫·阿汗,吾尔科木·冉合木,古丽巴哈尔·托乎提,金阿芳. 籽棉团悬浮速度的理论算法与试验验证. 农业工程学报. 2022(24): 52-62 . 百度学术
13. 张三丰,熊威,柯林,黄静林,沈学锋. 针对饲料气力输送系统设备选型参数的理论及仿真计算. 中国水运. 2022(16): 72-74 . 百度学术
14. 王晓晨,高自成,李立君,廖凯,庞国友,赵凯杰. 基于CFD-DEM油茶果负压吸附系统数值模拟及试验研究. 农机化研究. 2021(08): 192-197 . 百度学术
15. 侯娟,周为峰,王鲁民,樊伟,原作辉. 中国深远海养殖潜力的空间分析. 资源科学. 2020(07): 1325-1337 . 百度学术
16. 仝玉超. 锂离子电池正负极材料气力输送系统研究. 决策探索(中). 2020(08): 94 . 百度学术
17. 林礼群,王志勇. 工船养殖颗粒饲料气力输送系统参数优化. 船舶工程. 2020(S2): 51-55 . 百度学术
其他类型引用(11)


计量
- 文章访问数: 3547
- HTML全文浏览量: 203
- PDF下载量: 2042
- 被引次数: 28
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
