
Analysis of length-weight relationship for major fishing species from Bohai Bay
-
摘要:
对2008年~2009年渤海湾渔业资源调查的22种主要渔获体长(叉长、头胸甲长、头胸甲宽、胴体长)和体质量数据进行统计,并对各生物种类体长与体质量关系进行幂函数回归。结果显示,22种主要渔获包括分属4目、7科的9种鱼类,8种虾蟹类隶属十足目下的6个科,4种软体类属于2个目、2个科,以及1种虾蛄;22个种类体长与体质量关系回归的指数b介于1.271~3.316,主要集中在2.979附近,其中10种是等速生长,另10种是负异速生长,还有2种是正异速生长;除4种虾外,其余种类的体长与体质量回归式的相关系数R2均超过0.81,幂函数对体长与体质量数据的拟合效果较好。
Abstract:We described the relationship between length (fork length, carapace length, carapace width and carcass length) and weight for 22 species of fishing resources caught during 2008~2009 in Bohai Bay and conducted a regression analysis of power function. A total of 4 478 individuals belong to 8 orders and 16 families. The value of the exponent b in the length-weight relationship ranged from 1.271 to 3.316 and was mainly about 2.979. The t-test indicates that 10 species were of isometric growth pattern, 10 species were in negative algometry and 2 species were in positive algometry. The R2 of length-weight of all species except 4 species were larger than 0.81. The present length-weight key for 22 species of fishing resources is a valuable tool for fishery managers.
-
Keywords:
- Bohai Bay /
- fishery resources /
- length-weight relationship /
- growth type
-
渔业资源生物学研究是从事渔业科研和管理的重要基础内容,在所有生物学资料中,体长和体质量数据最易获得。而建立体长与体质量关系是资源评估中最基础的资料,根据体长预测体质量[1],已在形态比较、个体发育变化的分析、不同区系群体生活史的比较[2-3]、渔获量估算、现存生物量的评估等方面得到广泛应用。
渔业资源种类的长度和质量(通常为体长和体质量)的关系常以幂函数方程式W=aLb表达[4],其中指数b提供了资源的生长信息,当b=3时,表示该种类为等速生长,即个体从小到大的生长过程中,长、宽、高方向的生长速度相等;当b不等于3时,表示异速生长,即生长过程中3个线度方向的生长速度不等[5]。国内外学者对渔业资源种类的体长和体质量关系的研究较多,但与国外相比[5-6],中国的研究主要集中在对单一种类的体长-体质量关系分析,系统地研究一个海区调查所有资源种类的长度与体质量关系的报道较少[7]。文章通过对渤海湾渔业资源调查渔获物形态学数据分析,系统地阐述该海区主要渔获物长度与体质量关系,为丰富近海渔业资源基础生物学资料,进行群体对比分析、资源评估、渔获产量预测等奠定基础。
1. 材料与方法
研究所用数据为2008年6月和9月以及2009年5月渤海湾资源调查资料,取样及生物学测定均根据《海洋调查规范》[8]进行,每网次渔获各种类样本数量少于50尾的全部取样,多于50尾的则随机抽取50尾;样品的生物学测定分网次进行,测定项目包括体长、体质量、胃饱满度、性别和性腺成熟度等,长度测量精确到1 mm,体质量精确到0.01 g。
数据的统计分析主要采用R软件(i386 3.0.2)进行,根据数据分析需求,统计每一生物的样本量,体长(叉长、头胸甲长、头胸甲宽或胴体长)和体质量的均值、范围、标准误,基于体长和体质量的幂函数关系(W=aLb,其中a,b为待估算参数和指数),利用最小二乘法估算参数a和指数b,并对指数b的分布进行统计分析,当b=3时为等速生长,b < 3为负异速生长,b>3为正异速生长。
2. 结果与分析
2.1 样本的统计学特征
研究对象共22种,包括9种鱼类,分属4目、7科,8种虾、蟹类,隶属十足目下的6个科,4种软体类,属于2个目、2个科,以及1种虾蛄。样本数量差异较大,范围从19尾(长蛸)到1 168尾(口虾蛄),共计4 478尾。每种生物的样本量,测量的项目,体长、体质量的范围和均值(±标准误),体长与体质量的幂函数回归结果,以及指数b的95%置信区间等详见表 1。经统计分析,鲬鱼长度和体质量的标准误均最大,其次为长蛸,其中鲬鱼长度和体质量的标准误均大于10,长蛸仅体质量的标准误超过10,长度的标准误为2.98,且鲬鱼的体质量变异系数(CV)达到了2.58。除4种虾外,其余种类的体长(叉长、头胸甲长、头胸甲宽或胴体长)与体质量回归式的相关系数R2值均超过0.81,对回归方程进行F检验,除鲜明鼓虾的P大于0.001外(P=0.001 5),其余种类的P均小于0.001,说明幂函数对体长(叉长、头胸甲长、头胸甲宽或胴体长)与体质量的拟合效果较好。
表 1 渤海湾22种主要种类长度与体质量回归统计Table 1. Statistics of length-weight relationship for 22 main fishing species from Bohai Bay种类
species样本量
quantity of sample长度类型
type of length体长/mm length 体质量/g weight 幂函数参数 coefficient of nonlinear regression 范围
range均值±标准误
X±SE范围
range均值±标准误
X±SEa b±95% CI R2 银鲳 Pampus argenteus 98 FL 43~102 81.06±1.11 1.86~30.21 16.20±0.59 1.477e-05 3.151±0.004 0.969 矛尾 
640 BL 26~167 89.49±1.10 0.20~40.21 9.28±0.29 1.710e-05 2.884±0.004 0.967 斑尾复 76 BL 91~214 134.86±2.64 7.84~99.43 25.82±1.65 9.580e-05 2.534±0.010 0.811 黄鲫 Setipinna taty 161 BL 33~155 80.94±1.91 0.38~33.90 5.43±0.48 8.207e-06 2.985±0.007 0.963 叫姑鱼 Johnius grypotus 53 BL 67~121 94.83±1.59 4.57~36.50 15.65±0.91 5.018e-06 3.271±0.010 0.870 斑 
168 BL 64~143 89.95±0.91 2.46~38.34 9.19±0.37 2.832e-06 3.316±0.004 0.935 鲬鱼 Platycephalus indicus 44 BL 25~383 104.34±11.78 0.55~435.08 30.92±12.03 6.635e-05 2.583±0.034 0.930 小黄鱼 Larimichthys polyactis 296 BL 55~191 84.51±1.42 2.23~92.50 11.54±0.67 2.560e-05 2.884±0.003 0.977 焦氏舌鳎 Cynoglossus (Areliscus) joyneri 149 BL 54~162 108.12±1.93 0.71~28.50 8.54±0.43 3.054e-06 3.137±0.005 0.966 中国明对虾 Fenneropenaeus chinensis 35 CL 33~54 45.54±0.93 13.53~37.31 26.18±0.89 2.023e-01 1.271±0.012 0.593 鲜明鼓虾 Alpheus distinguendus 30 CL 15~24 20.47±0.38 2.17~8.89 5.88±0.32 1.101e-02 2.066±0.035 0.375 日本鼓虾 A.japonicus 73 CL 7~16 11.95±0.24 0.40~3.53 1.53±0.07 2.698e-02 1.603±0.033 0.408 葛氏长臂虾 Palaemon gravieri 182 CL 5~23 10.17±0.26 0.17~6.32 1.03±0.07 1.457e-02 1.767±0.021 0.707 隆线强蟹 Eucrate crenata 153 CW 11~42 28.25±0.42 0.99~34.02 12.76±0.55 1.035e-03 2.787±0.012 0.832 日本关公蟹 Dorippe japonica 32 CW 17~27 22.77±0.44 3.61~11.22 7.05±0.40 1.290e-03 2.743±0.013 0.886 日本 
124 CW 17~78 45.94±1.29 0.78~114.50 27.41±2.15 1.945e-04 3.026±0.008 0.971 三疣梭子蟹 Portunus trituberculatus 222 CW 27~171 79.07±1.54 1.31~342.95 32.35±2.88 4.873e-05 2.993±0.009 0.905 长蛸 Octopus cf. minor 19 ML 39~81 66±2.98 20.32~163.01 106.61±10.54 1.314e-03 2.678±0.025 0.878 短蛸 O.fangsiao 31 ML 25~56 42.61±1.24 7.75~87.04 42.98±3.53 4.318e-04 3.044±0.018 0.901 日本枪乌贼 Loliolus japonica 99 ML 43~102 81.27±1.12 1.86~48.30 16.52±0.67 1.220e-05 3.196±0.004 0.963 火枪乌贼 L.beka 625 ML 11~67 42.17±0.39 0.54~19.98 5.54±0.12 1.901e-03 2.109±0.005 0.868 口虾蛄 Oratosquilla oratoria 1168 CL 8~35 22.46±0.14 1.00~61.44 15.81±0.26 3.133e-03 2.699±0.006 0.824 注:FL.叉长;BL.体长;CL.头胸甲长;CW.头胸甲宽;ML.胴体长
Note:FL.fork length;BL.body length;CL.carapace length;CW.carapace width;ML.mantle length2.2 指数b的分布特点
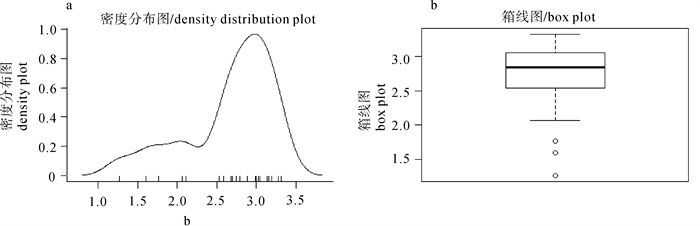
22种研究对象的体长(叉长、头胸甲长、头胸甲宽或胴体长)与体质量幂函数回归关系中,指数b范围介于1.271~3.316,均值为2.670(SE=±0.565),5个种类的指数b小于2.5,无指数b大于3.5的。分析22种研究对象的指数b密度分布(图 1-a),指数b主要集中在2.979附近,呈单峰右偏分布,左侧有明显拖尾现象,为4种虾类和火枪乌贼的指数b均未超过2.2造成。箱线图(图 1-b)显示指数b的中位数为2.836,上、下四分位数分别为3.04和2.55,即50%种类的指数b介于该区间,占全部被测生物指数b分布区间的23.96%;且中位数距上四分位数较近,同样可以说明指数b密度的右偏分布。生长类型根据对指数b进行t检验来确定[5],结果显示有10个种类(银鲳、矛尾
![]() 图 1 体长与体质量拟合关系中指数b的分布密度(a)及箱线图(b)Figure 1. Density distribution (a) and box-plot (b) of b-value in length-weight regression curve
图 1 体长与体质量拟合关系中指数b的分布密度(a)及箱线图(b)Figure 1. Density distribution (a) and box-plot (b) of b-value in length-weight regression curve3. 讨论
渔业资源种类的体长和体质量数据是资源生物学资料中最基本的信息,两者的关系不仅可以为研究资源的生长、种群分析等内容提供基础,同时还可以间接反映生活空间条件变动[9]。建立体长和体质量的回归关系,从统计学角度讲,样本数量特征和数据分布特点等对结果有明显影响,通常样本的数量会直接影响到回归效果[10],研究的22种生物中,有6种生物的样本量少于50,其中长蛸的样本量仅19尾,尽管其回归方程显著相关(F检验,P < 0.001),拟合效果很好(R2=0.878),但样本数量较少的情况下:1)存在不一定能够代表总体的可能;2)基于已建立的体长体质量关系式,对样本覆盖范围内的体质量进行预测可得到相对准确的结果,对超出样本范围的体质量进行预测,准确度是难以得到保证的[5]。因此,调查取样过程中对于样本数量相对少的种类,可以通过增加样本量,扩大数据的跨度范围,使数据更具代表性,从而提高拟合效果和方程的预测准确性。
样本数量影响回归效果的同时,数据的分布特点也会影响到回归效果,当体质量数据的CV偏大时,利用回归方程和平均体长进行平均体质量的估算,估计值与对应体长水平下通过观察值计算得到的体质量均值偏差较大,且通常偏小[11]。笔者研究中也存在此现象,22种生物体质量的CV分析中鲬鱼体质量CV值最大(2.58),尽管鲬鱼体长与体质量的幂函数回归相关系数R2达到0.930,处于22个回归方程第8位,F检验的P远小于0.001(P=2.2e-16),从统计学角度讲曲线对数据的拟合效果非常好,但利用回归方程和平均体长进行平均体质量估算时,估计值较对应体长水平下通过计算观察值得到的体质量均值偏小20.068,为全部研究对象中的最大者。造成体质量数据CV偏大的原因:1)与长度水平下体质量的离散程度有关;2)与样本特征值的范围有关。该研究中鲬鱼体长跨度达358 mm,体质量跨度超过400 g,两者均为22种研究对象对应性状的最大值。从利用回归关系进行预测的适用范围和准确度的角度来讲,样本特征值跨度大,回归结果具有更广的应用空间[5],即鲬鱼的拟合关系较其他生物的会有更好的应用基础。
与鱼类的长度和体质量关系拟合相比,虾、蟹类以及口虾蛄的蜕皮生长对长度和体质量关系的拟合效果亦会产生影响。就单个个体而言,蜕皮使其呈现出阶梯式生长,但因蜕皮只是给虾、蟹类以及口虾蛄提供了生长的机会,是否生长主要取决于饵料条件,且蜕皮与生长的生理调节方式是各自独立的[12],同时群体内的个体蜕皮非刀刃型,时间上存在非同步性,因此对群体而言,体长、体质量数据仍具有连续性[13],可应用W=aLb进行分析,但样本中蜕皮个体数量所占比例会成为影响回归效果的关键,笔者研究中中国明对虾、鲜明鼓虾和日本鼓虾回归方程拟合效果较差可能与此有关,认为可以通过选择恰当的取样时间(如越冬期、生殖前期等)或增加样本数量来减小这种影响。
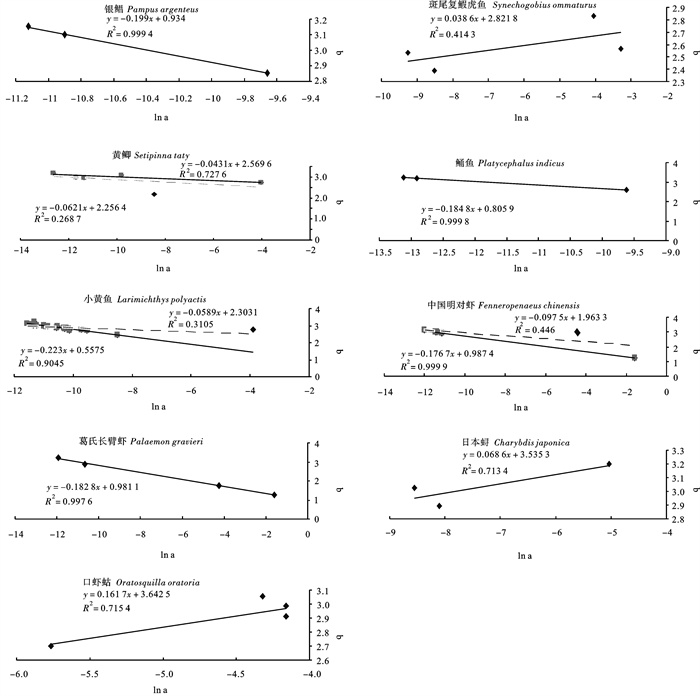
有报道指出对同一种鱼类而言,尽管采样区域不同,但鱼的体长-体质量幂指数关系参数a的对数与指数b呈显著负线性关系[14-15]。对已报道的关于银鲳[16-17]、斑尾复
![]() 图 2 长度与体质量关系参数a的自然对数与指数b的线性关系Figure 2. Linear relationship between lna and b for length-weight relationship表 2 已发表文章中与笔者研究相同种类的采样、性状及回归信息Table 2. Same data of sampling, traits and regression from the published paper
图 2 长度与体质量关系参数a的自然对数与指数b的线性关系Figure 2. Linear relationship between lna and b for length-weight relationship表 2 已发表文章中与笔者研究相同种类的采样、性状及回归信息Table 2. Same data of sampling, traits and regression from the published paper种类
species时间
time海域
sea样本量
quantity of sample数据特征 trait 回归结果 result of regression 文献和备注
references & note长度/mm
length体质量/g
weightA b R 银鲳 Pampus argenteus 2007 渤海 54.5~246.1 4.4~465 6.389e-05 2.856 6 [16] 2007 渤海 179 1.85e-05 3.098 3 0.993 3 [17] 斑尾复 1986~1988 东海 950 0.0173 9 2.827 9 [18] 1997~1999, 2001~2002 渤海 1 291 65~452 4.3~365 2e-04 2.39 0.977 1 [19] 1973~1974 黄海 365 0.037 3 2.565 8 [20] 黄鲫 Setipinna taty 2000~2002 东海 315 59~191 2~68 3.3e-06 3.192 11 0.975 1 [21] 2005~2006 黄海 1 477 70~110 0~15 1.144e-05 2.976 6 0.983 1 [22] 1991~1992 渤海 7 764 5.4e-05 3.100 8 0.898 [23]成鱼 1984~1986 渤海 2.1e-04 2.16 0.987 [23]幼鱼 2000 黄海 115 79~165 5~54 1.77e-02 2.73 [24] 鲬鱼 Platycephalus indicus 1966 黄海 742 167~500 29~971 2.48e-06 3.195 9 >0.9 [25] 2010 渤海 202 62~327 3.7~318.1 2e-06 3.225 3 0.994 6 [26] 小黄鱼 Larimichthys polyactis 1960 渤海 833 181~230 91~200 1.186e-05 3.046 0.977 0 [27] 1982 渤海 461 111~150 1.315e-05 3.042 0.975 8 1993 渤海 604 111~130 1.667e-05 2.986 0.985 8 2003 渤海 405 111~150 21~60 3.351e-05 2.836 0.957 3 2002~2003 东海, 黄海 804 9.4288e-06 3.116 1 0.98 [28] 2001 东海 3 771 58~220 7~194 2.6611e-05 2.951 3 0.956 5 [29] 1963 东海 520 110~330 26~605 3.5651e-05 2.870 5 0.981 1 [30] 1983 东海 524 109~277 25~330 7.2046e-05 2.729 9 0.970 4 2001 东海 894 58~191 13~146 2.6594e-05 2.912 6 0.956 9 1988 东海 161 1.765e-05 2.957 4 0.996 4 [31] 2004 渤海 101~250 5~170 4.007e-05 2.693 0 [32] 2000 黄海 358 8.4~16.9 8.11~64.4 0.020 38 2.791 0.957 [33] 1960 黄海 2 000 13~34.5 25~775 1.22e-05 3.246 0.956 7 [34] 1985 黄海 1 500 8.7~24.0 9.2~240 1.10e-05 3.079 0.961 0 1998 黄海 2 000 7.8~17.0 9.6~102 1.66e-05 3.008 0.977 0 2008 黄海 2 000 7.4~20.5 9~131 3.91e-05 2.831 0.972 0 2004 东海 2e-04 2.450 0 0.998 5 [35] 2005 东海 127 5.91e-05 2.750 2 0.953 1 2006 东海 126 3.27e-05 2.875 7 0.990 7 2007 东海 126 1.92e-05 2.992 4 0.931 1 中国明对虾 Fenneropenaeus chinensis 1957 渤海 9 1.51~18.05 0.037~67.38 0.011 73 3.009 5 [13]♀ 1957 渤海 9 1.51~14.4 0.04~37.00 0.012 1 2.979 5 [13]♂ 1971, 1984, 1985 黄海 6.0195e-06 3.125 8 0.997 [36] 1990 东海 449 1.429e-05 2.954 0.985 9 [37]♀ 1990 东海 507 1.446e-05 2.951 0.982 3 [37]♂ 2010 黄海 312 1.14e-05 2.997 0.994 0 [38]♀ 2010 黄海 247 1.05e-05 3.012 0.989 9 [38]♂ 葛氏长臂虾 Palaemon gravieri 1986~1992 东海 1 927 24~76 0.1~8.8 6.6415e-06 3.209 4 0.999 6 [39]♀ 1986~1992 东海 987 26~58 0.1~2.8 2.3727e-05 2.868 6 0.999 3 [39]♂ 日本 1998~1999 东海 3.0274e-04 2.891 0 0.994 [40]♀ 1998~1999 东海 6.5280e-03 3.201 1 0.982 [40]♂ 口虾蛄 Oratosquilla oratoria 2004~2005 黄海 2 378 58.7~169.1 2.8~68.0 0.015 6 2.986 4 0.978 2 [41] 0.01558 2.91 0.981 [42] 0.0156 2.91 0.981 [43] 2007~2008 黄海 1 608 35~154 0.59~53.00 0.0133 3.058 2 0.989 8 [44] 就渔业资源而言,体长和体质量主要受生物自身的生长规律及生态学因素的影响[49],从而作用于回归的结果。通常情况下,随着生物体生活阶段、饵料基础、水文条件、胃肠饱满度、性腺发育、繁殖活动等影响[50],体长与体质量回归关系的参数a和指数b都会发生变化,其中指数b主要受生物体身体3个方向(长、宽、高)的生长速度影响,即体形和比重在生长过程中是否发生变化,对于绝大多数种类而言,指数b的年度变化不显著[11];参数a受生活阶段、环境条件、采样时间等因素影响,较易发生改变[10],该次调查时间跨度较大,造成样本各自所处的生理状态及环境特征存在差异;同时,回归方程采用最小二乘法进行参数估算,此方法仅以残差平方和最小为唯一标准。因此拟合过程中,参数a和指数b受数据的分布特点影响会产生联动效应,故而有些种类的指数b估算效果不好。
-
![]()
图 1 体长与体质量拟合关系中指数b的分布密度(a)及箱线图(b)
Figure 1. Density distribution (a) and box-plot (b) of b-value in length-weight regression curve
![]()
图 2 长度与体质量关系参数a的自然对数与指数b的线性关系
Figure 2. Linear relationship between lna and b for length-weight relationship
表 1 渤海湾22种主要种类长度与体质量回归统计
Table 1 Statistics of length-weight relationship for 22 main fishing species from Bohai Bay
种类
species样本量
quantity of sample长度类型
type of length体长/mm length 体质量/g weight 幂函数参数 coefficient of nonlinear regression 范围
range均值±标准误
X±SE范围
range均值±标准误
X±SEa b±95% CI R2 银鲳 Pampus argenteus 98 FL 43~102 81.06±1.11 1.86~30.21 16.20±0.59 1.477e-05 3.151±0.004 0.969 矛尾 640 BL 26~167 89.49±1.10 0.20~40.21 9.28±0.29 1.710e-05 2.884±0.004 0.967 斑尾复 76 BL 91~214 134.86±2.64 7.84~99.43 25.82±1.65 9.580e-05 2.534±0.010 0.811 黄鲫 Setipinna taty 161 BL 33~155 80.94±1.91 0.38~33.90 5.43±0.48 8.207e-06 2.985±0.007 0.963 叫姑鱼 Johnius grypotus 53 BL 67~121 94.83±1.59 4.57~36.50 15.65±0.91 5.018e-06 3.271±0.010 0.870 斑 168 BL 64~143 89.95±0.91 2.46~38.34 9.19±0.37 2.832e-06 3.316±0.004 0.935 鲬鱼 Platycephalus indicus 44 BL 25~383 104.34±11.78 0.55~435.08 30.92±12.03 6.635e-05 2.583±0.034 0.930 小黄鱼 Larimichthys polyactis 296 BL 55~191 84.51±1.42 2.23~92.50 11.54±0.67 2.560e-05 2.884±0.003 0.977 焦氏舌鳎 Cynoglossus (Areliscus) joyneri 149 BL 54~162 108.12±1.93 0.71~28.50 8.54±0.43 3.054e-06 3.137±0.005 0.966 中国明对虾 Fenneropenaeus chinensis 35 CL 33~54 45.54±0.93 13.53~37.31 26.18±0.89 2.023e-01 1.271±0.012 0.593 鲜明鼓虾 Alpheus distinguendus 30 CL 15~24 20.47±0.38 2.17~8.89 5.88±0.32 1.101e-02 2.066±0.035 0.375 日本鼓虾 A.japonicus 73 CL 7~16 11.95±0.24 0.40~3.53 1.53±0.07 2.698e-02 1.603±0.033 0.408 葛氏长臂虾 Palaemon gravieri 182 CL 5~23 10.17±0.26 0.17~6.32 1.03±0.07 1.457e-02 1.767±0.021 0.707 隆线强蟹 Eucrate crenata 153 CW 11~42 28.25±0.42 0.99~34.02 12.76±0.55 1.035e-03 2.787±0.012 0.832 日本关公蟹 Dorippe japonica 32 CW 17~27 22.77±0.44 3.61~11.22 7.05±0.40 1.290e-03 2.743±0.013 0.886 日本 124 CW 17~78 45.94±1.29 0.78~114.50 27.41±2.15 1.945e-04 3.026±0.008 0.971 三疣梭子蟹 Portunus trituberculatus 222 CW 27~171 79.07±1.54 1.31~342.95 32.35±2.88 4.873e-05 2.993±0.009 0.905 长蛸 Octopus cf. minor 19 ML 39~81 66±2.98 20.32~163.01 106.61±10.54 1.314e-03 2.678±0.025 0.878 短蛸 O.fangsiao 31 ML 25~56 42.61±1.24 7.75~87.04 42.98±3.53 4.318e-04 3.044±0.018 0.901 日本枪乌贼 Loliolus japonica 99 ML 43~102 81.27±1.12 1.86~48.30 16.52±0.67 1.220e-05 3.196±0.004 0.963 火枪乌贼 L.beka 625 ML 11~67 42.17±0.39 0.54~19.98 5.54±0.12 1.901e-03 2.109±0.005 0.868 口虾蛄 Oratosquilla oratoria 1168 CL 8~35 22.46±0.14 1.00~61.44 15.81±0.26 3.133e-03 2.699±0.006 0.824 注:FL.叉长;BL.体长;CL.头胸甲长;CW.头胸甲宽;ML.胴体长
Note:FL.fork length;BL.body length;CL.carapace length;CW.carapace width;ML.mantle length 下载: 导出CSV
下载: 导出CSV
表 2 已发表文章中与笔者研究相同种类的采样、性状及回归信息
Table 2 Same data of sampling, traits and regression from the published paper
种类
species时间
time海域
sea样本量
quantity of sample数据特征 trait 回归结果 result of regression 文献和备注
references & note长度/mm
length体质量/g
weightA b R 银鲳 Pampus argenteus 2007 渤海 54.5~246.1 4.4~465 6.389e-05 2.856 6 [16] 2007 渤海 179 1.85e-05 3.098 3 0.993 3 [17] 斑尾复 1986~1988 东海 950 0.0173 9 2.827 9 [18] 1997~1999, 2001~2002 渤海 1 291 65~452 4.3~365 2e-04 2.39 0.977 1 [19] 1973~1974 黄海 365 0.037 3 2.565 8 [20] 黄鲫 Setipinna taty 2000~2002 东海 315 59~191 2~68 3.3e-06 3.192 11 0.975 1 [21] 2005~2006 黄海 1 477 70~110 0~15 1.144e-05 2.976 6 0.983 1 [22] 1991~1992 渤海 7 764 5.4e-05 3.100 8 0.898 [23]成鱼 1984~1986 渤海 2.1e-04 2.16 0.987 [23]幼鱼 2000 黄海 115 79~165 5~54 1.77e-02 2.73 [24] 鲬鱼 Platycephalus indicus 1966 黄海 742 167~500 29~971 2.48e-06 3.195 9 >0.9 [25] 2010 渤海 202 62~327 3.7~318.1 2e-06 3.225 3 0.994 6 [26] 小黄鱼 Larimichthys polyactis 1960 渤海 833 181~230 91~200 1.186e-05 3.046 0.977 0 [27] 1982 渤海 461 111~150 1.315e-05 3.042 0.975 8 1993 渤海 604 111~130 1.667e-05 2.986 0.985 8 2003 渤海 405 111~150 21~60 3.351e-05 2.836 0.957 3 2002~2003 东海, 黄海 804 9.4288e-06 3.116 1 0.98 [28] 2001 东海 3 771 58~220 7~194 2.6611e-05 2.951 3 0.956 5 [29] 1963 东海 520 110~330 26~605 3.5651e-05 2.870 5 0.981 1 [30] 1983 东海 524 109~277 25~330 7.2046e-05 2.729 9 0.970 4 2001 东海 894 58~191 13~146 2.6594e-05 2.912 6 0.956 9 1988 东海 161 1.765e-05 2.957 4 0.996 4 [31] 2004 渤海 101~250 5~170 4.007e-05 2.693 0 [32] 2000 黄海 358 8.4~16.9 8.11~64.4 0.020 38 2.791 0.957 [33] 1960 黄海 2 000 13~34.5 25~775 1.22e-05 3.246 0.956 7 [34] 1985 黄海 1 500 8.7~24.0 9.2~240 1.10e-05 3.079 0.961 0 1998 黄海 2 000 7.8~17.0 9.6~102 1.66e-05 3.008 0.977 0 2008 黄海 2 000 7.4~20.5 9~131 3.91e-05 2.831 0.972 0 2004 东海 2e-04 2.450 0 0.998 5 [35] 2005 东海 127 5.91e-05 2.750 2 0.953 1 2006 东海 126 3.27e-05 2.875 7 0.990 7 2007 东海 126 1.92e-05 2.992 4 0.931 1 中国明对虾 Fenneropenaeus chinensis 1957 渤海 9 1.51~18.05 0.037~67.38 0.011 73 3.009 5 [13]♀ 1957 渤海 9 1.51~14.4 0.04~37.00 0.012 1 2.979 5 [13]♂ 1971, 1984, 1985 黄海 6.0195e-06 3.125 8 0.997 [36] 1990 东海 449 1.429e-05 2.954 0.985 9 [37]♀ 1990 东海 507 1.446e-05 2.951 0.982 3 [37]♂ 2010 黄海 312 1.14e-05 2.997 0.994 0 [38]♀ 2010 黄海 247 1.05e-05 3.012 0.989 9 [38]♂ 葛氏长臂虾 Palaemon gravieri 1986~1992 东海 1 927 24~76 0.1~8.8 6.6415e-06 3.209 4 0.999 6 [39]♀ 1986~1992 东海 987 26~58 0.1~2.8 2.3727e-05 2.868 6 0.999 3 [39]♂ 日本 1998~1999 东海 3.0274e-04 2.891 0 0.994 [40]♀ 1998~1999 东海 6.5280e-03 3.201 1 0.982 [40]♂ 口虾蛄 Oratosquilla oratoria 2004~2005 黄海 2 378 58.7~169.1 2.8~68.0 0.015 6 2.986 4 0.978 2 [41] 0.01558 2.91 0.981 [42] 0.0156 2.91 0.981 [43] 2007~2008 黄海 1 608 35~154 0.59~53.00 0.0133 3.058 2 0.989 8 [44]
下载: 导出CSV
-
[1] GARCIA C B, DUARTE J O, SANDOVAL N, et al. length-weight relationships of demersal fishes from the Gulf of Salamanca, Colombia[J]. Naga, ICLARM Quarterly, 1998, 21(3): 30-32. https://digitalarchive.worldfishcenter.org/items/23734f41-bae1-4756-94d5-3271b65f2e24
[2] WEATHERLEY A H, GILL H S, CASSELMAN J M. The biology of fish growth[M]. London: Academic Press, 1987: 36-42. doi: 10.2307/4790
[3] PETRAKIS G, STERGIOU K I. 1995. Weight-length relationships for 33 fish species in Greek waters[J]. Fish Res, 1995, 21(3/4): 465-469. doi: 10.1016/0165-7836(94)00294-7
[4] RICKER W E. Linear regressions in fishery research[J]. J Fish Res Board Can, 1973, 30(3): 409-434. doi: 10.1139/f73-072
[5] MOREY G, MORANTA J, MASSUTI E, et al. Weight length relationships of littoral to lower slope fishes from the western Mediterranean[J]. Fish Res, 2003, 62(1): 89-96. doi: 10.1016/S0165-7836(02)00250-3
[6] ECOUTIN J M, ALBARET J J, TRAPE S. Length weight relationships for fish populations of a relatively undisturbed tropical estuary: the Gambia[J]. Fish Res, 2005, 72(2/3): 347-351. doi: 10.1016/j.fishres.2004.10.007
[7] 王雪辉, 杜飞雁, 邱永松. 南海北部主要经济鱼类体长与体重关系[J]. 台湾海峡, 2006, 25(2): 262-266. doi: 10.3969/j.issn.1000-8160.2006.02.017 [8] 张玉生, 杨清良, 陈瑞祥, 等. GB/T12763.6-2007, 海洋调查规范[S]. 北京: 中国标准出版社, 2007: 38-44. https://www.doc88.com/p-778380121612.html [9] SPARRE P, VENEMA S C. Introduction to tropical fish stock assessment (Part I. Manual)[J]. FAO Fisheries Technical Paper, 1989(306): 18-20. doi: 10.1067/mge.2000.107714
[10] HUXLEY J S. Constant differential growth-ratios and their significance[J]. Nature, 1924, 114(2877): 895-896. doi: 10.1038/114895a0
[11] 詹秉义. 渔业资源评估[M]. 北京: 中国农业出版社, 1995: 18-24. https://xueshu.baidu.com/usercenter/paper/show?paperid=7d780225e511d43a52f368e49751c36b&site=xueshu_se&hitarticle=1 [12] FREEMAN J A. Molt increment, molt cycle duration, and tissue growth in Palamonetes pugio Holthuis larvae[J]. J Exp Mar Biol Ecol, 1990, 143(1/2): 47-61. doi: 10.1016/0022-0981(90)90110-X
[13] 张乃禹. 中国对虾生长的数理分析[J]. 海洋科学, 1985, 9(4): 1-6. https://xueshu.baidu.com/usercenter/paper/show?paperid=23553d653091f25515266c2656b95812&site=xueshu_se&hitarticle=1 [14] STERGIOU K I, MOUTOPOULOS D K. A review of length-weight relationships of fishes from Greek marine waters[J]. Naga, 2001, 24(1/2): 23-39. https://www.researchgate.net/publication/235732640_A_Review_of_Length-Weight_Relationships_of_Fishes_from_Greek_Marine_Waters
[15] 李忠炉, 金显仕, 单秀娟, 等. 小黄鱼体长-体质量关系和肥满度的年际变化[J]. 中国水产科学, 2011, 18(3): 602-610. doi: 10.3724/SP.J.1118.2011.00602 [16] 许玉甫, 周军, 张国胜, 等. 河北沿海银鲳渔业资源现状分析[J]. 河北渔业, 2009(6): 4-6, 15. doi: 10.3969/j.issn.1004-6755.2009.06.003 [17] 崔青曼, 袁春营, 董景岗, 等. 渤海湾银鲳年龄与生长的初步研究[J]. 天津科技大学学报, 2008, 23(3): 30-32. doi: 10.3969/j.issn.1672-6510.2008.03.008 [18] 孙帼英, 陈建国. 斑尾复虎鱼的生物学研究[J]. 水产学报, 1993, 17(2): 146-153. [19] 范海洋, 纪毓鹏, 张士华, 等. 黄河三角洲斑尾复虾虎鱼渔业生物学的研究[J]. 中国海洋大学学报, 2005, 35(5): 733-736. doi: 10.3969/j.issn.1672-5174.2005.05.007 [20] 陈大刚. 鱼港内斑尾复虎鱼生物学的初步调查[J]. 动物学杂志, 1979, 14(1): 3-6. [21] 刘勇, 程家骅, 李圣法. 东海区黄鲫资源的利用现状及合理利用探讨[J]. 中国水产科学, 2006, 13(3): 485-491. doi: 10.3321/j.issn:1005-8737.2006.03.025 [22] 熊瑛, 汤建华, 刘培廷, 等. 黄海南部黄鲫(Setipinna taty)资源利用分析[J]. 海洋与湖沼, 2009, 40(4): 500-505. https://lib.cqvip.com/Qikan/Article/Detail?id=31239794&from=Qikan_Search_Index [23] 张孟海, 王勇, 张军. 渤海南部黄鲫生长与死亡特性的研究[J]. 浙江海洋学院学报: 自然科学版, 2004, 23(1): 31-36. doi: 10.3969/j.issn.1008-830X.2004.01.007 [24] 孙蜀东, 任一平. 黄海南部黄鲫Setipinna taty (Cuvier et Valenciennes)渔业生物学研究[J]. 海洋湖沼通报, 2003(1): 62-65. doi: 10.3969/j.issn.1003-6482.2003.01.010 [25] 陈万青, 赵维谦. 黄海鲬鱼年龄和生长的初步研究[J]. 水产学报, 1998, 22(3): 289-304. https://xueshu.baidu.com/usercenter/paper/show?paperid=7838d0efbb397df6791a8f3dddff228b&site=xueshu_se&hitarticle=1 [26] 秦岩, 高天翔. 东营近海鲬鱼渔业生物学及资源丰度的季节变化[J]. 中国海洋大学学报, 2012, 42(7/8): 106-111. [27] 郭旭鹏, 金显仕, 戴芳群. 渤海小黄鱼生长特征的变化[J]. 中国水产科学, 2006, 13(2): 243-249. doi: 10.3321/j.issn:1005-8737.2006.02.013 [28] 严利平, 胡芬, 凌建忠, 等. 东海北部和黄海南部小黄鱼年龄与生长的研究[J]. 中国海洋大学学报, 2006, 36(1): 95-100. doi: 10.3969/j.issn.1672-5174.2006.01.019 [29] 林龙山, 程家骅. 东海区小黄鱼渔业生物学现状的分析[J]. 中国海洋大学学报, 2006, 34(4): 565-570. doi: 10.3969/j.issn.1672-5174.2004.04.008 [30] 林龙山, 程家骅, 任一平, 等. 东海区小黄鱼种群生物学特性的分析[J]. 中国水产科学, 2004, 11(4): 333-338. doi: 10.3321/j.issn:1005-8737.2004.04.010 [31] 柳卫海, 郭振华, 詹秉义. 东海区小黄鱼资源利用现状分析[J]. 上海水产大学学报, 1999, 8(2): 105-111. https://xueshu.baidu.com/usercenter/paper/show?paperid=6484ea029689d06cf2d284c1ab47d96d&site=xueshu_se&hitarticle=1 [32] 周军, 李怡群, 张海鹏, 等. 河北省近海海域小黄鱼分布和生物学特征[J]. 河北渔业, 2006(10): 46-48. doi: 10.3969/j.issn.1004-6755.2006.10.022 [33] 任一平, 高天翔, 刘群, 等. 黄海南部小黄鱼Pseudosciaena plyactis (Bleeker)渔获群体结构与繁殖特征的初步研究[J]. 海洋湖沼通报, 2001(1): 41-46. [34] 张国政, 李显森, 金显仕, 等. 黄海中南部小黄鱼生物学特征的变化[J]. 生态学报2010, 30(24): 6854-6861. [35] 尹增强, 章守宇, 汪振华, 等. 浙江嵊泗人工鱼礁区小黄鱼生长特征与资源合理利用的初步研究[J]. 中国生态农业学报, 2010, 18(3): 588-594. doi: 10.3724/SP.J.1011.2010.00588 [36] 陈宗尧, 刘永昌, 邱盛尧, 等. 黄海中部沿岸放流增殖对虾生长特性初步研究[J]. 海洋学报, 1990, 12(6): 758-764. https://xueshu.baidu.com/usercenter/paper/show?paperid=0398080127b8c7df45e40bfd94216bd5&site=xueshu_se&hitarticle=1 [37] 张澄茂. 闽东海区中国对虾放流虾的生长特性[J]. 水产学报, 2001, 25(2): 116-119. [38] 徐炳庆. 山东近海中国对虾增殖放流的研究[D]. 上海: 上海海洋大学, 2011: 26-27. 10.7666/d.y1946982 [39] 丁天明, 宋海棠. 东海葛氏长臂虾Palaemon gravieri生物学特征研究[J]. 浙江海洋学院学报: 自然科学版, 2002, 21(1): 1-5. doi: 10.3969/j.issn.1008-830X.2002.01.001 [40] 俞存根, 宋海棠, 姚光展. 东海日本的数量分布和生物学特性[J]. 上海水产大学学报, 2005, 14(1): 40-45. [41] 徐海龙, 张桂芬, 乔秀亭, 等. 黄海北部口虾蛄体长及体质量关系研究[J]. 水产科学, 2010, 29(8): 451-454. doi: 10.3969/j.issn.1003-1111.2010.08.003 [42] 王春琳, 徐善良, 梅文骧, 等. 口虾蛄的生物学基本特征[J]. 浙江水产学院学报, 1996, 15(1): 60-62. https://xueshu.baidu.com/usercenter/paper/show?paperid=4460a4eb68508a19b0efab826c7cfdba&site=xueshu_se&hitarticle=1 [43] 王波, 张锡烈, 孙丕喜. 口虾蛄的生物学特征及其人工苗种生产技术[J]. 黄渤海海洋, 1998, 16(2): 64-73. https://xueshu.baidu.com/usercenter/paper/show?paperid=f98b86c85b28a2e1296871e022ce7df9&site=xueshu_se&hitarticle=1 [44] 盛福利. 青岛近海口虾蛄Oratosquilla oratoria渔业生物学的初步研究[D]. 青岛: 中国海洋大学, 2009: 27. [45] OLURIN K B, SAVAGE O D. Reproductive biology, length-weight relationship and condition factor of the African snake head, Parachanna obscura, from River Oshun, South-west Nigeria[J]. Int J Fish Aquac, 2011, 3(8): 146-150. https://www.internationalscholarsjournals.com/abstract/reproductive-biology-lengthweight-relationship-and-condition-factor-of-the-african-snake-head-parachanna-obscura-from-ri-59344.html
[46] BOSILJKA M, GORENKA S. Reproduction, length-weight relationship and condition of sardine, Sardina pilchardus (Walbaum, 1792), in the eastern Middle Adriatic Sea (Croatia)[J]. Periodicum Biologorum, 2010, 112(2): 133-138. https://www.researchgate.net/publication/277873386_Reproduction_length-weight_relationship_and_condition_of_sardine_Sardina_pilchardus_Walbaum_1792_in_the_eastern_Middle_Adriatic_Sea_Croatia
[47] KUMOLU-JOHNSON C A, NDIMELE P E. Length-weight relationships of nine fish species from Ologe Lagoon, Lagos, Nigeria[J]. Afr J Biotechnol, 2011, 10(2): 241-243. doi: 10.4314/AJB.V10I2
[48] PEREIRAL J N, SIMAS A, ROSA A, et al. Weight-length relationships for 27 demersal fish species caught off the Cape Verde archipelago (eastern North Atlantic)[J]. J Appl Ichthyol, 2011, 24(1): 1-4. doi: 10.1111/j.1439-0426.2011.01915.x
[49] CHERIF M, ZARRAD R, GHARBI H, et al. Length-weight relationships for 11 fish species from the Gulf of Tunis (SW Mediterranean Sea, Tunisia)[J]. Pan-Am J Aquat Sci, 2008, 3(1): 1-5. https://www.semanticscholar.org/paper/Length-weight-relationships-for-11-fish-species-the-Ch%C3%A9rif-Zarrad/a7527c9f9406d935381031fcb1e7f7fcffcd5b44
[50] KUMOLU-JOHNSON C A, NDIMELE P E. Length-weight relationships and condition factors of twenty-one fish species in Ologe Lagoon, Lagos, Nigeria[J]. Asian J Agric Sci, 2010, 2(4): 174-179. https://www.researchgate.net/publication/49582898_Length-Weight_Relationships_and_Condition_Factors_of_Twenty-One_Fish_Species_in_Ologe_Lagoon_Lagos_Nigeria
-
期刊类型引用(5)
1. 刘婷婷,刘小丽,邢燕,高慧,王勤,魏滨. 山东沿海6市市售水产品中多氯联苯污染特征及健康风险评估. 食品安全导刊. 2025(01): 99-102+108 .  百度学术
百度学术
2. 陈征兵,王楚. 水环境中持久性有机污染物风险评价研究进展. 黑龙江环境通报. 2024(01): 10-12 . 百度学术
3. 马丽莎,谢文平,田斐,尹怡,单奇,郑光明,李恒翔. 广东沿海养殖牡蛎中多氯联苯残留水平及人体饮食暴露风险评估. 南方水产科学. 2021(02): 11-19 . 本站查看
4. 杨代凤,刘腾飞,陆皓茜,毛健,钱辉,赵佳昕. 气相色谱-质谱法测定小麦中二噁英类多氯联苯. 食品安全质量检测学报. 2020(17): 6209-6215 . 百度学术
5. 肖妙,张彩云,李玉卡,刘二花,苏青. 畜产品中持久性有机污染物危害及检测. 今日畜牧兽医. 2019(10): 1-2 . 百度学术
其他类型引用(1)
计量
- 文章访问数: 3361
- HTML全文浏览量: 165
- PDF下载量: 2627
- 被引次数: 6
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
