
Numerical simulation of the forces and deformation of HDPE circular gravity cages
-
摘要:
该研究旨在综合探讨不同网箱周长、浮管管径、网衣高度及网目大小对整体网箱受力变形的影响,为网箱的科学合理选型提供数据参考。设定的网箱周长40~80 m,浮管管径250~630 mm,网衣高度6~20 m,网目大小45~115 mm。通过数值模拟方法对4种规格高密度聚乙烯圆形网箱在不同组合条件下网箱锚绳受力、波流力以及容积损失率进行了数值计算。结果表明,大规格网箱的锚绳受力、波流力更大,容积损失率更小,锚绳数量的增加可以大大降低锚绳受力。相比浮管管径,网衣高度和网目大小对网箱受力变形的影响更显著。整体网箱的受力变形随着网衣高度的增加而增大,随网目的增大而减小。
Abstract:The aim of this study is to comprehensively discuss the effects of cage perimeter, pipe diameter, net height as well as net mesh size on the forces and deformation of deep-water net cages in waves and current, thus provide references to select net-cages scientifically. We simulated HDPE cages with perimeter 40~80 m, pipe diameter 250~630 mm, net height 6~20 m and net mesh size 45~115 mm. With a numerical model previously validated by physical model tests, the mooring line forces, the wave-current forces and the volume reduction rate of the four kinds of HDPE cages are calculated. The simulated results indicate that the mooring line force and wave-current force are greater while the volume reduction rate is smaller for bigger cage. Moreover, the mooring line force of cage could be greatly reduced by adding mooring lines. Compared to pipe diameter, net height and net mesh size have more significant impact on the cage′s force and deformation, which increase with net height increasing, while decrease with net mesh size increasing.
-
Keywords:
- deep-water net cage /
- force and deformation /
- numerical analysis
-
黄边糙鸟蛤(Trachycardium flavum)隶属软体动物门,瓣鳃纲,帘蛤目,鸟蛤科,俗称“鸡腿螺”、“美人腿”。生活在低潮线附近,珊瑚礁间的砂质海滩或浅海,是中国广东、广西和海南潮间带习见的暖水性、埋潜性种类[1]。其肉味鲜美,营养丰富,具有很高的经济和食用价值,为国内外畅销紧俏海产品。目前,对于黄边糙鸟蛤的研究仅见于其分类特征、生态习性以及生态资源调查[1-3]、呼吸排泄[4]、生殖腺的组织学[5]的报道,未见利用通径分析和多元回归分析对其主要形态性状对质量性状如体质量、软体部质量的影响效果的报道。
近年来,多元相关方法在水产经济动物的生产量估计方面得到了广泛应用[6-10]。在贝类选择育种中,质量性状如体质量、软体部质量作为良种选育最直接的目标性状和直接反映贝类的生产性能的性状,一直以来是贝类研究的热点。弄清贝类形态性状和质量性状间的关系是贝类选择育种目标性状制定的的重要依据。王雨等[11]研究了海南野生长肋日月贝(Amusium pleuronecte)形态性状与质量性状的相关性,认为壳高可作为长肋日月贝育苗或育种的测量指标。鄢朝等[12]研究了华贵栉孔扇贝(Chlamys nobilis)数量性状的相关性及通径分析,为其辅助选择育种提供相关理论依据。肖述等[13]采用通径分析方法对湛江沿海香港巨牡蛎(Crassostrea hongkongens)雌雄群体经济性状进行分析。AHMED等[14]利用多元相关对鱼、鲸和贝类幼龄期体长、体质量相关的生长参数进行了分析。刘志刚等[15]对华贵栉孔扇贝主要经济性状与闭壳肌质量关系进行了分析,认为壳宽可作为主要的选育指标。栗志民等[16]采用多元线性回归和通径分析方法,研究了企鹅珍珠贝(Pteria penguin)主要经济性状对体质量的影响效果。刘小林等[17]研究了栉孔扇贝(C.farreri)贝壳尺寸性状与其活体质量的相关性,表明壳高是影响活体质量的决定性因素。刘文广等[18]对不同贝龄华贵栉孔扇贝数量性状进行了通径分析,指出在6月龄后以壳高为主要选择指标,可达到对体质量的选择,为华贵栉孔扇贝选择育种中选育指标的确立提供了理论指导。质量性状如活体质量、软体部质量是衡量黄边糙鸟蛤产量以及经济效益的主要指标,直接影响黄边糙鸟蛤增养殖的经济效益。笔者试验采用通径分析和多元回归的方法研究了黄边糙鸟蛤各形态性状与质量性状的关系及对质量性状的真实影响效应,从而找出与质量性状的关联度最大的目标性状,为选择育种和种质资源研究提供理论依据。
1. 材料与方法
1.1 材料
2012年6月22日,从海南陵水县新村港黄边糙鸟蛤野生群体中随机取样80个个体,贝龄1~2年,壳长为(39.510±3.956) mm,体质量为(24.496±6.356) mm,海区水温为(30.5±1.0)℃。
1.2 方法
受测的5个经济性状包括壳长、壳高、壳宽3个形态性状和2个质量性状(活体质量、软体部质量),测定前把贝体表面附着生物和淤泥洗刷干净。测定标准是: 壳长为贝壳前后与铰合线平行的最大距离;壳高为从壳顶至腹缘与铰合线垂直的最大距离;壳宽为捏紧两边贝壳使壳宽不再变小时,测量壳左右两侧间与铰合线垂直的最大距离;活体质量为倒去壳内海水,用纱布吸去表面及壳缘水分后的质量;软体部质量为取出软体部,用纱布吸去表面水分后的质量。形态性状采用游标卡尺(精度为0.01 mm)测量,活体质量和软体部质量采用电子天平(精度为0.01 g)称量。
相关数据分析处理参照张琪等[19]、陈飞飞等[20]方法,利用Excel 2003和SPSS 13.0软件对各测量数据进行处理分析,获得各性状表型参数统计量。分析性状间的表型相关、性状对质量性状通径分析以及决定系数,剖析各性状对质量性状的直接作用效应和间接作用效应。通过剔除偏回归系数不显著的性状,利用多元回归分析建立偏回归系数显著的性状与质量性状间的多元回归方程,并对方程进行显著性检验。相关计算公式为:
$$ \begin{array}{l} {r_{xy}} = \frac{{\sum\limits_{i = 1}^n {\left( {{x_i} - x} \right)} \left( {{y_i} - \bar y} \right)}}{{\sum\limits_{i = 1}^n {{{\left( {{x_i} - x} \right)}^2}} {{\left( {{y_i} - \bar y} \right)}^2}}}\\ {P_{yxi}} = b{x_i}{\sigma _{{x_i}}}/{\sigma _y};{P_{{x_i}{y_j}}} = {P_{iy}}{r_{ij}}{P_{jy}}(i \ne j)\\ {d_i} = {p_{iy}}^2;{d_{ij}} = 2{r_{ij}}{P_{iy}}\\ d = \sum\limits_{i = 1}^n {{d_i}} + \sum\limits_{\begin{array}{*{20}{c}} {i = 1}\\ {i \ne j} \end{array}}^n {\sum\limits_{j = 1}^n {{d_{ij}}} } ;R_i^2 = {d_i} + \frac{1}{2}\sum\limits_{\begin{array}{*{20}{c}} {i = 1}\\ {i \ne j} \end{array}}^n {{d_{ij}}} \end{array} $$ 式中rxy为xi对yi的相关系数,xi是自变量的标志值,yi是因变量的标志值;Pyxi为自变量xi对依变量y的直接通径系数,bxi为自变量的回归系数;σxi为自变量的标准差;σy为因变量的标准差;Pij为第i个性状通过第j个性状产生的间接通径系数。di为i性状对依变量y的直接作用系数;dij为第i个性状通过第j个性状对依变量y的间接作用系数。d为所有性状的总决定系数,Ri2为各性状对y性状的总决定系数。
质量性状(Y)的回归方程采用的线性模型:
$$ Y=b_0+b_i x_1+b_0 x_i+b_i x_2+\cdots+b_0 x_i $$ 式中Y为依变量;b0是常数项;bi是自变量xi对依变量Y的偏回归系数。
2. 结果与分析
2.1 各性状数据描述统计
文章测量了野生黄边糙鸟蛤80只,其主要性状的表型参数见表 1。各性状的变异系数范围为10.01%~31.16%,其变异系数从大到小的顺序为软体部质量>活体质量>壳高>壳宽>壳长。壳长、壳高和壳宽3个形态性状的变异系数大小相近,而2个质量性状软体部质量和活体质量的变异系数相对于形态性状而言较大,其中软体部质量的变异系数最大(31.16%)。各性状的标准误差均较小,且偏度值与0相接近,表明测量样本各性状数据均服从正态分布,可用于下一步的相关分析、多元回归分析及通径分析。
表 1 野生黄边糙鸟蛤测量性状的表型统计(n=80)Table 1. Descriptive statistics of 5 economic traits of wild T.flavum性状trait 壳长/mm shell length 壳高/mm shell height 壳宽/mm shell width 软体部质量/g edible-part weight 活体质量/g body weight 平均值mean 39.510 42.748 30.101 6.927 24.496 标准差standard deviation 3.956 4.702 3.118 2.158 6.356 标准误差standard error 0.442 0.525 0.349 0.241 0.711 峰度Kurtosis 0.046 -0.166 -0.083 -0.664 -0.508 偏度Skewness -0.452 -0.715 -0.381 0.082 -0.140 变异系数/% CV 10.01 11.00 10.36 31.16 25.94 2.2 性状间的表型相关
黄边糙鸟蛤性状之间的表型相关分析结果见表 2。各性状间的相关系数均达到极显著水平(P < 0.01)。各性状与活体质量之间的相关系数大小依次为软体部质量(0.966 7)>壳高(0.845 6)>壳长(0.839 1)>壳宽(0.777 8),表明软体部质量对活体质量的决定效应最大,壳高次之。而形态性状与软体部质量的相关系数大小依次为壳高(0.783 7)>壳长(0.779 4)>壳宽(0.747 8)。但在多变量情况下,变量间的表型相关系数仅反映各性状之间的直接和间接相关的总和,不能准确反映各性状对质量性状间的真实相关程度,有必要对这种相关关系作进一步的分析。
表 2 黄边糙鸟蛤各性状间的表型相关Table 2. Phenotype correlation coefficients among various traits of T.flavum性状trait 壳长shell length 壳高shell height 壳宽shell width 软体部质量edible-part weight 壳高shell height 0.920 0** 壳宽shell width 0.607 2** 0.663 5** 软体部质量edible-part weight 0.779 4** 0.783 7** 0.747 8** 活体质量body weight 0.839 1** 0.845 6** 0.777 8** 0.966 7** 注: * *.P < 0.01; *.P < 0.05; 后表同此
Note: * *.P < 0.01; *.P < 0.05;The same case in the following tables.2.3 各性状对质量性状影响的通径分析
2.3.1 对活体质量的通径分析
根据通径分析原理,利用SPSS 13.0和Excel 2003将黄边糙鸟蛤各经济性状对活体质量的影响作用剖分为直接作用和间接作用,结果见表 3。壳长、壳宽、软体部质量的通径系数存在统计学意义,软体部质量和壳宽对活体质量的直接作用呈现极显著水平(P < 0.01),壳长对活体质量的直接作用呈现显著水平(P < 0.05),而壳高对活体质量的直接作用不显著(P>0.05)。这些测量性状中各性状对活体质量的直接作用由大到小依次为软体部质量(0.704 8)、壳长(0.142 6)、壳宽(0.105 0)和壳高(0.093 1)。软体部质量通过壳长、壳宽以及壳高3个性状对活体质量均产生不同程度的正向间接作用,但间接作用明显小于其直接作用,因此,软体部质量是影响黄边糙鸟蛤活体质量的决定因素。而壳长、壳宽及壳高3个性状的直接作用相对较小,但3个性状通过软体部质量对活体质量的间接作用都较大,且均大于其直接作用,表明壳长、壳高、壳宽对活体质量的影响主要通过软体部质量实现的。因此,以活体质量为选育黄边糙鸟蛤高产品种时首先以软体质量为选育目标,同时加强壳长、壳宽以及壳高3个性状的间接选择可能会取得较好的效果。
表 3 各性状对活体质量的通径分析Table 3. Path analysis of effects of various traits on body weight性状trait 相关系数related coefficient 直接作用direct effect 间接作用indirect effect 壳长shell length 壳高shell height 壳宽shell width 软体部质量edible-part weight ∑ 壳长shell length 0.839 1** 0.142 6* - 0.085 6 0.063 8 0.549 4 0.698 8 壳高shell height 0.845 6** 0.093 1 0.131 2 - 0.069 7 0.552 3 0.753 2 壳宽shell width 0.777 8** 0.105 0** 0.086 6 0.061 8 - 0.527 1 0.675 4 软体部质量edible-part weight 0.966 7** 0.704 8** 0.111 1 0.072 9 0.078 5 - 0.262 6 2.3.2 对软体部质量的通径分析
壳长、壳高、壳宽3个形态性状对软体部质量产生的直接作用并不相同,其中壳宽对软体部质量直接影响最大,其直接作用为0.409 3,呈极显著相关(P < 0.01);其次为壳长,其直接作用为0.389 2,呈显著相关(P < 0.05);而壳高对软体部质量的直接影响最小(0.154 0),相关性不显著(P>0.05)(表 4)。因此,在以黄边糙鸟蛤软体质量为选育目标时,从提高产量的角度出发,应首先以壳宽为选育指标。
表 4 各形态性状对软体部质量的通径分析Table 4. Path analysis of effects of morphological traits on edible-part weight性状trait 相关系数related coefficient 直接作用direct effect 间接作用indirect effect 壳长shell length 壳高shell height 壳宽shell width ∑ 壳长shell length 0.779 4** 0.389 2* - 0.141 7 0.248 5 0.390 2 壳高shell height 0.783 7** 0.154 0 0.358 1 - 0.271 6 0.629 7 壳宽shell width 0.747 8** 0.409 3** 0.236 3 0.102 2 - 0.338 5 3个形态性状通过其他性状都能够对软体部质量产生正向的间接影响,作用大小依次为壳高(0.629 7)>壳长(0.390 2)>壳宽(0.338 5)(表 4)。除壳宽的直接效应大于其间接效应外,其他2个性状的间接效应均大于其直接效应,特别是壳高,其对软体质量产生的间接效应约是其直接效应的4倍。这一结果表明,以软体部质量选育黄边糙鸟蛤高产品种时应注重壳宽的直接选择,同时加强对壳高协同作用的选择。
2.4 各性状对质量性状的决定程度分析
2.4.1 对活体质量的决定程度分析
黄边糙鸟蛤各性状对活体质量的决定程度分析见表 5。对角线上为单一性状的直接决定系数,对角线上方为一
表 5 各性状对活体质量的决定系数Table 5. Determination coefficients of various traits on body weight性状trait 壳长shell length 壳高shell height 壳宽shell width 软体部质量edible-part weight ∑ 壳长shell length 0.020 3 0.024 4 0.018 2 0.156 0 0.219 0 壳高shell height 0.008 7 0.013 0 0.102 7 0.124 3 壳宽shell width 0.011 0 0.110 1 0.121 2 软体部质量edible-part weight 0.496 8 0.496 8 所有性状总决定系数determination coefficient of all traits 0.9612 剩余项的决定系数remaining determination coefficient 0.0388 注:对角线上为单一性状的直接决定系数,对角线上方为一性状通过另一性状的间接决定系数;后表同此
Note: The direct determination coefficients for a single trait are on the diagonal; the indirect determination coefficients produced by a trait through another are above the diagonal;The same case in the following table.性状通过另一性状的间接决定系数。所有性状决定系数总和为0.961 2,表明影响黄边糙鸟蛤活体质量的主要性状已被纳入该研究中。就直接决定系数而言,软体部质量对活体质量的直接决定程度最大(49.68%),其次分别为壳长(2.03%)、壳宽(1.10%)。在共同决定程度中,壳长、壳高、壳宽与软体部质量间的协同作用较大,分别为15.60%、10.27%、11.01%。因此,以活体质量为目标性状选育黄边糙鸟蛤高产品种时应以软体部质量为首选指标,同时加强壳长、壳宽以及壳高协同作用的选择,特别是壳长。
2.4.2 对软体部质量的决定程度分析
黄边糙鸟蛤各形态性状对软体部质量的决定程度分析见表 6。壳长、壳高以及壳宽对软体部质量的决定系数总和为0.730 1,接近1,表明影响黄边糙鸟蛤软体部质量的主要性状已找到,但其他性状如活体质量、性腺质量等对软体部质量产生一定程度的影响。壳长、壳高和壳宽对软体部质量的直接决定程度分别为15.15%、2.37%和16.75%,其中壳宽的决定程度最大,表明壳宽是影响黄边糙鸟蛤软体部质量的主要影响因素。在间接决定系数中,壳长通过壳宽对软体部质量的间接决定程度最大(19.35%)。这些结果表明,以软体部质量选育高产黄边糙鸟蛤品种时,从形态性状而言应首选壳宽,同时加强壳长的间接协同作用的选择。
表 6 各主要形态性状对软体部质量的决定系数Table 6. Determination coefficients of morphological traits on edible-part weight性状trait 壳长shell length 壳高shell height 壳宽shell width ∑ 壳长shell length 0.151 5 0.110 3 0.193 5 0.455 3 壳高shell height 0.023 7 0.083 6 0.107 4 壳宽shell width 0.167 5 0.167 5 所有性状总决定系数determination coefficient of all traits 0.730 1 剩余项的决定系数remaining determination coefficient 0.269 9 2.5 多元回归方程的建立及显著性检验
2.5.1 主要性状对活体质量的多元回归分析
根据多元相关和通径分析,以黄边糙鸟蛤活体质量为依变量,其他性状为自变量进行多元回归分析。偏回归系数检验表明,剔除偏回归系数不显著的壳高,壳长、壳宽以及软体部质量偏回归系数均达到极显著水平(P < 0.01)(表 7)。因此,可建立由壳长、壳宽以及软体部质量这3个性状估计黄边糙鸟蛤活体质量的多元线性回归方程:
$$ \begin{array}{r} y_1=-10.467 9+0.339 2 X_1+0.229 9 X_2+ 2.114 2 X_3 \end{array} $$ 表 7 黄边糙鸟蛤主要性状对活体质量的偏回归系数检验Table 7. Partial regression coefficient test for various traits to body weight of T.flavum偏回归系数partial regression coefficient 标准误差standard error t 显著性significance 截距intercept -10.467 9 2.379 8 -4.398 6 0** 壳长shell length 0.339 2 0.059 8 5.674 0 0** 壳宽shell width 0.229 9 0.071 6 3.212 4 0.001 9** 软体部质量edible-part weight 2.114 2 0.131 1 16.123 4 0* 其中R2 = 0.958 8,y1为活体质量(g),x1为壳长(mm),x2为壳宽(mm),x3为软体部质量(g)。方差分析结果显示黄边糙鸟蛤活体质量与3个性状间的线性回归关系达到极显著水平(P < 0.01)(表 8)。经回归预测,估计值与实际观察值差异不显著(P>0.05),表明该方程客观地反映了黄边糙鸟蛤主要性状与活体质量的真实关系,可以简便地应用于实际生产中。
表 8 主要性状对活体质量的多元回归的方差分析Table 8. ANOVA analysis on multi-variance regression between the main traits and body weight指标index 自由度DF 总平方和SS 均方MS F 显著性significance 回归分析regression model 3 3 060.222 0 1 020.074 0 590.405 3 0** 残差residual 76 131.309 1 1.727 7 总计total 79 3 191.531 0 2.5.2 各形态性状对软体部质量的多元回归分析
对可能影响黄边糙鸟蛤软体部质量的形态性状的偏回归系数进行显著性检验,发现壳高对软体部质量的标准偏回归系数不显著(P>0.05),剔除壳高,对其他2个性状偏回归系数进行检验,均达到极显著水平(P < 0.01)(表 9)。剔除直接影响不显著的壳高,建立以软体部质量为依变量,壳长、壳宽为自变量的多元线性回归方程:
$$ {y_2} = - 13.240 8 + 0.281 2{x_1} + 0.301 0{x_2}\left( {{R^2} = } \right.0.726 9) $$ 表 9 形态性状对软体部质量的偏回归系数检验Table 9. Partial regression coefficients test for morphological traits to edible-part weight偏回归系数partial regression coefficient 标准误差standard error t 显著性significance 截距intercept -13.240 8 1.414 5 -9.360 7 0** 壳长shell length 0.281 2 0.040 9 6.876 0 0** 壳宽shell width 0.301 0 0.051 9 5.802 2 0** 式中y2为软体部质量(g),x1为壳长(mm),x2为壳宽(mm)。
由表 10的多元回归方差分析结果表明,壳长、壳宽2个形态性状与软体部质量之间的回归达到极显著水平(P < 0.01)。
表 10 形态性状对软体部质量的多元回归方差分析Table 10. ANOVA analysis of multi-variance regression of morphological traits to edible-part weight指标index 自由度DF 总平方和SS 均方MS F 显著性significance 回归分析regression model 2 267.486 9 133.743 5 102.484 5 0 残差residual 77 100.485 9 1.305 0 总计total 79 367.972 8 3. 讨论
3.1 自变量的确定以及与通径分析特点
表型相关系数是变量间相互关系的综合体现,由于未剔除其他变量的影响,导致自变量对依变量的直接作用和通过其他自变量对依变量的间接作用的正负作用可以相互抵消,因而不能全面地对变量间的相互关系进行阐释。而通径系数是变量标准化后的偏回归系数,能够有效地区分原因对结果的直接和间接影响,其中直接影响部分即是通径系数,它不受其他有关变量的影响,因而能真实地反映原因对结果的相对重要性[21]。该研究中黄边糙鸟蛤各性状间的相关关系均达到极显著水平(P < 0.01),但并不能说明所有的性状都是影响依变量的主要因素,因而有必要进行通径分析,找出影响其质量性状的主要决定因素。在通径分析中通径系数容易随自变量个数和性质不同而改变,自变量的更换或者个数的增减都会使得通径系数发生变化,考虑自变量性状个数越多,分析结果越可靠,但统计分析也就越复杂,不能突出重点[17, 22]。通常情况下只有表型相关系数达到显著水平的自变量才能进行通径分析,而未达到显著水平的自变量应给予剔除。该研究对黄边糙鸟蛤5个性状进行表型相关分析,结果表明各性状两两间的相关系数均呈现正相关,其显著水平均达到极显著(P < 0.01),表明该数据可用于下一步的统计分析,确保了统计分析的实际意义。
3.2 影响黄边糙鸟蛤质量性状的主要因素确定
笔者研究中软体部质量对活体质量的直接决定作用最大(49.68%),且其直接作用明显大于其间接作用。在其他影响活体质量的性状中,对活体质量的直接作用大小依次为壳长(0.142 6)、壳宽(0.105 0)和壳高(0.093 1),但这3个性状的间接作用均大于其直接作用(表 5),其中壳长通过软体部质量对活体质量的间接决定作用最大,其决定作用15.60%,表明它们对活体质量的影响主要通过软体部质量实现的。这一结果与栗志民等[16]的研究结果相似。利用3个形态性状对软体部质量的决定程度分析发现,起决定作用最大的是壳宽(16.75%),且其直接作用大于间接作用;而壳长、壳高这2个性状的直接作用小于间接作用,其中壳长通过壳宽对软体部质量的间接决定程度最大(19.35%)。上述分析结果与孙泽伟等[6]认为壳高是影响近江牡蛎(Crassostrea rivularis)软体部质量的主要决定因素有所不同。这种差异可能是引入的变量不同所致,笔者研究中仅对壳长、壳宽和壳高对软体部质量的相关性进行了分析,而他们的自变量除壳长、壳高以及壳宽外,还包括全质量。
在表型相关分析的基础上进行通径系数分析和决定系数分析,只有当复相关指数或各自变量对依变量的单独决定系数及两两共同决定系数的总和∑d (数值上R2=∑d) 大于或等于0.85时,表明影响依变量的主要自变量已经找到[23-24]。笔者研究中软体部质量、壳长以及壳宽与活体质量的决定系数总和R2=0.958 8>0.85,表明软体部质量、壳长和壳宽是影响黄边糙鸟蛤活体质量的主要性状。而壳长、壳高以及壳宽对软体部质量的多元回归分析检验中,壳长和壳宽作用显著,其决定系数总和R2=0.726 9,略小于0.85,表明影响软体部质量的性状已基本找到,壳长和壳宽是决定黄边糙鸟蛤软体部质量的主要构成因子。这一结果与王庆恒等[25]报道的壳宽、壳高是影响波纹巴非蛤(Paphia undulate)软体部质量的主要性状的结果有所不同,这可能是由于不同贝类的形态性状对其软体部质量的决定程度不同所致。
3.3 黄边糙鸟蛤通径分析的意义
体质量和软体部质量是衡量黄边糙鸟蛤经济价值的重要指标,在其苗种繁育和遗传育种中起到重要作用,可作为亲本选择的主要依据。黄边糙鸟蛤贝体铰合齿结合紧密,无法直观地测量其软体部质量,只能通过剖杀贝体,取出软体部进行称量,测量操作难度大且繁琐,且贝体死亡则难以用于苗种繁育和遗传育种。而活体质量往往由于其贝壳结合紧密,壳内积累大量水分无法准确测量,从而使得数据失去意义。但相对于质量性状而言,形态指标则易于准确测量。因此,弄清黄边糙鸟蛤形态性状与质量性状之间的关系,通过对形态性状的选择达到育种目的,具有重要的实践意义。然而,贝类的体质量和软体部质量易受其肥满度的影响,与个体大小、年龄、性成熟程度和季节等多个因素相关。该研究中试验材料为处于生长期的1~2龄黄边糙鸟蛤,采样时间为6月。一般而言,在该季节水温较高,贝类性腺发育较好,但随着大潮水的刺激(初一或十五左右)贝体会自动排精排卵,贝体的肥满度将大大降低。该研究的试验材料采自刚经历大潮水之后不久,贝体肥满度较小。因此该研究结果仅适用于黄边糙鸟蛤性腺发育生长期,至于繁殖期黄边糙鸟蛤形态性状与质量性状之间的相关关系有待进一步研究。
在黄边糙鸟蛤性腺发育生长期,以活体质量为目标性状选育黄边糙鸟蛤高产品种时应优先考虑软体部质量,其次为壳长和壳宽。但由于软体部质量在实际操作过程中无法活体测量,因而不适合作为直接的育种目标性状,因此可考虑易测形态性状壳长和壳宽。而在以软体部质量为目标性状的选育过程中,壳长和壳宽对软体部质量的通径系数和总决定系数较大,是影响黄边糙鸟蛤软体部质量的重要形态性状,可作为选育的主要目标性状。由此表明通过壳长和壳宽来描述黄边糙鸟蛤活体质量或软体部质量是可行的。在黄边糙鸟蛤苗种繁育以及遗传育种过程中应首先加强对壳宽的选择,同时利用对壳长的协同作用,为黄边糙鸟蛤的遗传选育确定选育所需的目标性状。
-
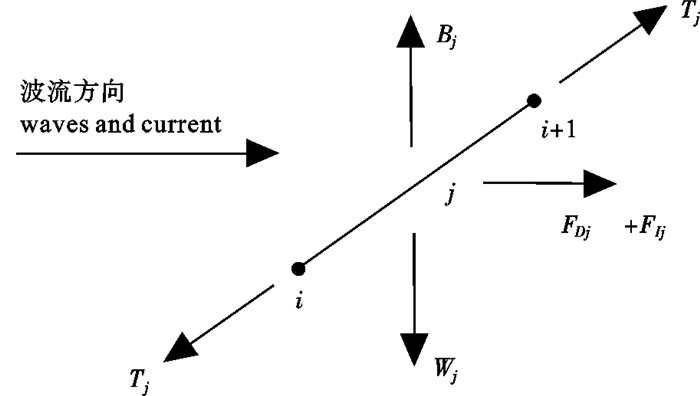
![]()
图 2 锚绳质点与构件计算示意图
Figure 2. Schematic diagram of the lumped mass point and the mooring line element for numerical calculations
表 1 不同规格网箱锚绳力和波流力计算结果(H=5 m,T=8 s,U=0.75 m · s-1)
Table 1 Calculated results of the mooring line forces and wave-current forces of the net-cages
网箱周长/m
perimeter of net-cage浮管管径/mm
pipe diameter网衣高度/m
net height网目大小/mm
net mesh size锚绳力/kN
mooring line force波流力/kN
wave-current force40 250 6 45 17.4 33.5 75 15.6 28.3 280 6 45 18.5 34.5 75 16.9 30.1 50 250 6 45 19.9 38.0 75 17.4 31.6 280 6 45 21.3 39.7 75 18.5 32.8 315 8 45 24.0 45.3 75 20.9 37.3 60 280 8 45 23.5 52.7 75 20.6 44.2 315 8 45 25.5 56.8 75 22.1 46.9 420 10 45 29.2 66.2 75 24.4 51.8 80 315 10 75 27.0 58.0 420 12 75 29.3 61.6 115 25.9 51.9 630 20 75 39.3 85.0 115 33.7 70.7  下载: 导出CSV
下载: 导出CSV
表 2 不同规格网箱变形计算结果(H=5 m,T=8 s,U=0.75 m · s-1)
Table 2 Calculated results of the net deformation of the net-cages
网箱周长/m
perimeter of net-cage浮管管径/mm
pipe diameter网衣高度/m
net height网目大小/mm
net mesh size网箱容积/m3
net-cage volume容积损失率/%
volume reduction rate40 250 6 45 331.6 56.6 75 378.2 50.5 280 6 45 315.5 58.7 75 364.4 52.3 50 250 6 45 539.7 54.8 75 600.6 49.7 280 6 45 526.6 55.9 75 591.0 50.5 315 8 45 636.8 60.0 75 708.4 55.5 60 280 8 45 1 070.4 53.3 75 1 196.4 47.8 315 8 45 1 015.4 55.7 75 1 159.8 49.4 420 10 45 1 140.3 60.2 75 1 286.4 55.1 80 315 10 75 2 597.4 49.0 420 12 75 2 994.9 51.0 115 3 379.9 44.7 630 20 75 4 196.6 58.8 115 4 481.8 56.0
下载: 导出CSV
-
[1] 郭根喜, 陶启友, 黄小华, 等. 深水网箱养殖装备技术前沿进展[J]. 中国农业科技导报, 2011, 13(5): 44-49. doi: 10.3969/j.issn.1008-0864.2011.05.07 [2] 黄小华, 郭根喜, 胡昱, 等. 波浪作用下圆形网箱浮架系统的运动特性分析[J]. 水产学报, 2009, 33(5): 878-884. doi: 10.3724/SP.J.00001 [3] 黄小华, 郭根喜, 胡昱, 等. 圆形网衣在水流作用下的运动变形特性[J]. 中国水产科学, 2010, 17(2): 312-319. https://www.fishscichina.com/zgsckx/article/abstract/1017?st=top_cited [4] 黄小华, 郭根喜, 胡昱, 等. 波流作用下深水网箱受力及运动变形的数值模拟[J]. 中国水产科学, 2011, 18(2): 443-450. doi: 10.3724/SP.J.1118.2011.0443 [5] 黄小华, 郭根喜, 胡昱, 等. HDPE圆柱形网箱与圆台形网箱受力变形特性的比较[J]. 水产学报, 2011, 35(1): 124-130. doi: 10.3724/SP.J.1231.2011.17135 [6] 郭根喜, 黄小华, 胡昱, 等. 高密度聚乙烯圆形网箱锚绳受力实测研究[J]. 中国水产科学, 2010, 17(4): 847-852. https://qikan.cqvip.com/Qikan/Article/Detail?id=34630989 [7] 唐宏结. 网箱容积变形改善研究[D]. 台北: 国立中山大学, 2002. [8] FREDRIKSSON D W, SWIFT M R, IRISH J D, et al. Fish cage and mooring system dynamics using physical and numerical models with field measurements[J]. Aquac Engin, 2003, 27(2): 117-146. doi: 10.1016/S0144-8609(02)00043-2
[9] LI Y C, GUI F K, TENG B. Hydrodynamic behavior of a straight floating pipe under wave conditions[J]. Ocean Engin, 2007, 34(3/4): 552-559. doi: 10.1016/J.OCEANENG.2006.01.012
[10] 郑艳娜, 董国海, 桂福坤, 等. 圆形重力式网箱锚碇系统的受力研究[J]. 应用力学学报, 2007, 24(2): 180-185. doi: 10.3969/j.issn.1000-4939.2007.02.003 [11] 郑国富, 黄桂芳, 魏观渊, 等. 波流作用下圆柱形近海抗风浪网箱缆绳的张力特性[J]. 水产学报, 2007, 31(1): 84-89. doi: 10.3321/j.issn:1000-0615.2007.01.012 [12] 黄六一, 梁振林, 万荣, 等. 波流作用下网格锚泊的单个重力式网箱缆绳张力[J]. 中国水产科学, 2011, 18(3): 636-645. doi: 10.3724/SP.J.1118.2011.00636 [13] DECEW J, FREDRIKSSON D W, BUGROV L, et al. A case study of a modified gravity type cage and mooring system using numerical and physical models[J]. IEEE J Oceanic Engin, 2005, 30(1): 47-58. doi: 10.1109/JOE.2004.841400
[14] HUANG C C, TANG H J, LIU J Y. Dynamical analysis of net cage structures for marine aquaculture: numerical simulation and model testing[J]. Aquac Engin, 2006, 35(3): 258-270. doi: 10.1016/j.aquaeng.2006.03.003
[15] HUANG C C, TANG H J, LIU J Y. Effects of waves and currents on gravity-type cages in the open sea[J]. Aquac Engin, 2008, 38(2): 105-116. doi: 10.1016/j.aquaeng.2008.01.003
[16] TSUKROV I, EROSHKIN O, PAUL W, et al. Numerical modeling of nonlinear elastic components of mooring systems[J]. IEEE J Oceanic Engin, 2005, 30(1): 37-46. doi: 10.1109/JOE.2004.841396
[17] ZHAO Y P, LI Y C, DONG G H, et al. A numerical study on dynamic properties of the gravity cage in combined wave-current flow[J]. Ocean Engin, 2007, 34(17/18): 2350-2363. http://www.onacademic.com/detail/journal_1000034030786010_5fdf.html
[18] DECEW J, TSUKROV I, RISSO A, et al. Modeling of dynamic behavior of a single-point moored submersible fish cage under currents[J]. Aquac Engin, 2010, 43(2): 38-45. doi: 10.1016/j.aquaeng.2010.05.002
[19] LEE C W, KIM Y B, LEE G H, et al. Dynamic simulation of a fish cage system subjected to currents and waves[J]. Ocean Engin, 2008, 35(14/15): 1521-1532. doi: 10.1016/J.OCEANENG.2008.06.009
[20] COLBOURNE D B, ALLEN J H. Observations on motions and loads in aquaculture cages from full scale and model scale measurements[J]. Aquac Engin, 2001, 24(2): 129-148. doi: 10.1016/S0144-8609(00)00069-8
[21] BESSONNEAU J S, MARICHAL D. Study of the dynamics of submerged supple nets (applications to trawls)[J]. Ocean Engin, 1998, 25(7): 563-583. doi: 10.1016/S0029-8018(97)00035-8
[22] 苏炜, 詹杰民. 等效网面法在模拟网的水动力特性中的应用[J]. 水动力学研究与进展: A辑, 2007, 22(3): 267-272. https://www.doc88.com/p-4764727594583.html -
期刊类型引用(2)
1. 王淼,周轩,洪波. 长江口桁拖网最小网目尺寸的选择性研究. 水产学杂志. 2024(04): 77-83 .  百度学术
百度学术
2. 董传举,张晓,詹世盈,李学军. 丹江口水库(河南辖区)主捕渔具调查研究. 河南师范大学学报(自然科学版). 2023(04): 25-35 . 百度学术
其他类型引用(6)
计量
- 文章访问数:
- HTML全文浏览量:
- PDF下载量:
- 被引次数: 8
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
