
Determination of methylene blue and its metabolites in aquatic products by UPLC/ESI-MS/MS
-
摘要:
建立了超高效液相色谱-电喷雾串联质谱法同时检测水产品中亚甲基蓝及其代谢物的分析方法。经过条件优化,试样用10 mL乙腈提取3次,并在体系中添加盐酸羟胺、对甲苯磺酸等提高提取效率,提取液经对丙磺酸固相萃取小柱净化,按V(乙腈)∶V(乙酸铵,1 mol·L-1)=1∶ 1洗脱,采用Thermo Hypersil C18色谱柱,以甲醇和0.2%甲酸为流动相进行梯度洗脱分离,电喷雾离子源(ESI),正离子模式,采用选择反应监测(SRM)扫描模式。结果表明,亚甲基蓝及代谢物(天青A、天青B、天青C)在0.50~100.00 μg·L-1范围内线性关系良好,相关系数R>0.99,在10 min内实现分离,3个加标水平(2 μg·kg-1、4 μg·kg-1、10 μg·kg-1)的平均回收率为74.23%~94.40%(n=6),相对标准偏差(RSD)为1.13%~10.28%,检测限(LOD)为0.40~0.60 μg·kg-1,定量限(LOQ)为2.00 μg·kg-1。该方法简便快捷、准确度高,易于推广应用,可作为快速测定水产品中亚甲基蓝及其代谢物的有效方法。
-
关键词:
- 超高效液相色谱-电喷雾串联质谱 /
- 亚甲基蓝及代谢物 /
- 水产品 /
- 检测
Abstract:We established a method for simultaneous determination of methylene blue and its metabolites in aquatic products using ultra-high performance liquid chromatography-electrospray ionization tandem mass spectrometry (UPLC/ESI-MS/MS). We extracted the sample with acetonitrile for three times and added hydroxylamine hydrochloride and paratoluenesulfonic acid for efficiency in the meantime, purified it by a PRS SPE column, and eluted by V(Acetonitrile) : V(Ammonium acetate, 1mol·L-1)=1 : 1. Next, based on Thermo Hypersil Gold C18 column, we used a methanol and 2% formic acid as gradient elution separation to deal with the sample, with the electrospray of mass spectrometry in positive ionization mode and the identifying of methylene blue and its metabolites in selected reaction monitoring (SRM mode). The results show that methylene blue and its metabolites have a good linear relationship within 0.50~100.00 μg·L-1(R>0.99). The separation is finished within 10 min. Average recovery (n=6) of the target compounds at 3 levels (2 μg·kg-1, 4 μg·kg-1, 10 μg·kg-1) is 78.11%~94.40%, while the relative standard deviation (RSD) is 2.12%~8.13%.Besides, the limit of detection (LOD) ranges from 0.40 to 0.60 μg·kg-1, and the limit of quantity (LOQ) is 2.00 μg·kg-1.The method is convenient, accurate, easily be applied and popularized, which is suitable for determination and confirmation of methylene blue and its metabolites in aquatic products.
-
亚甲基蓝(methylene blue,MB)又名次甲基蓝、碱性湖蓝、美蓝,是一种人工合成的噻嗪类染料,具有抗菌和消毒的作用,可用于治疗淡水鱼类的小爪虫病、斜管虫病、水霉病和凡纳滨对虾(Penaeus vannamei)幼体粘脏病,也可用作抗真菌药物 [1-3],降低鱼类运输中的死亡率。由于亚甲基蓝及其代谢物被认为对动物体有毒性并有致突变作用[4-8],在美国、日本等国家禁止用于水产养殖,在国内也未被批准使用。
亚甲基蓝及其代谢物残留的检测方法有分光光度法[9]、毛细管电泳法[10]、液相色谱法等[11-13],其中对水产品中的残留检测方法国外研究较少,国内主要是液相色谱法。超高效液相色谱串联质谱方法,可以实现同时对水产品中几种有害物质进行定性并定量检测,具有检测限低、准确度高的优点[14],是近年来食品中有害物质残留检测的发展趋势[15-17],笔者在已有研究的基础上[10-11],建立了使用超高效液相色谱-电喷雾串联质谱仪同时测定水产品中亚甲基蓝及其3种代谢产物——天青A、天青B、天青C含量的方法,具有实际意义和推广前景。
1. 材料与方法
1.1 试验材料
试验用草鱼(Ctenopharyngodon idellus)6尾,体长(60±2)cm,每尾体质量为(2.5±0.3)kg,去鳞、去皮,沿脊背取肌肉;凡纳滨对虾200尾,体长(10±1)cm,每尾体质量为(10±1.2)g,去头、去壳、去肠腺,取肌肉部分;中华绒鳌蟹(Eriocheir sinensis)50只,每只体质量为(100±7.5)g,取可食部分。样品充分绞碎、混匀,于-20 ℃冷冻保藏。试验所用水产品均购自南京市迎宾菜市场。
混合型阳离子交换柱(MCX)、C8/SCX混合模式柱(MCAX)、C18柱(C18)、对丙磺酸柱(PRS)(美国Agilent公司出品),规格均为500 mg/3 mL。
1.2 仪器
Accela超高效液相色谱、TSQ Quantum Access Max三重四级杆质谱仪(美国Thermo Fisher公司出品)、Allegra 64R高速冷冻离心机(美国Beckman公司出品)、Caliper Turbovap LV全自动浓缩工作站(美国Zymark公司出品)、12通道自动固相萃取装置(德国CNW公司出品)、十万分之一电子天平(德国Sartorius公司出品)、Milli-Q去离子水发生器(美国Millipore公司出品)。
1.3 试剂
亚甲基蓝、天青A和天青C购自美国Fluka公司,纯度均大于95%;天青B购自美国Sigma公司,纯度为89%。甲醇、乙腈、二氯甲烷、甲酸、乙酸铵均为色谱纯试剂(美国TEDIA公司出品),盐酸羟胺、对甲苯磺酸均为分析纯试剂。
1.4 样品前处理方法
称取5.00 g样品,其中中华绒螯蟹样品2.00 g,于50 mL离心管中分别加入0.5 g·mL-1的盐酸羟胺溶液0.75 mL、2 mol·L-1的对甲苯磺酸溶液1.25 mL、pH 4.5 0.1 mol·L-1的乙酸铵缓冲液5 mL,涡旋1 min;再加入10 mL乙腈涡旋1 min,8 000 r·min-1离心10 min,上清液倒入125 mL分液漏斗中;再用10 mL乙腈重复提取2次;加入25 mL二氯甲烷振荡萃取,静置1 h后取下层溶液用45 ℃氮气吹至近干。用3 mL乙腈溶解残渣,上样于用3 mL乙腈活化好的PRS柱,再用乙腈重复溶解残渣2次,待样液流尽后将柱子吹至近干,用3 mL洗脱液[V(乙酸铵,1 mol·L-1) : V(乙腈) =1 : 1]洗脱,洗脱液收集于10 mL玻璃试管中,用蒸馏水稀释到10 mL。涡旋混匀后过0.22 μm尼龙滤膜供质谱检测。
1.5 色谱条件
色谱柱为Thermo Hypersil Gold C18柱,50 mm×2.1 mm(内径),粒度1.9 μm;柱温为30 ℃;进样体积10 μL;流动相为甲醇+0.2%甲酸溶液,采用梯度洗脱的方法,梯度洗脱程序见表 1。
表 1 流动相梯度洗脱程序Table 1. Gradient elution with mobile phasest/min 0.2%甲酸(A)
0.2% methanoic acid甲醇(B)
methanol0.0 90 10 1.0 90 10 5.0 10 90 5.1 90 10 9.0 90 10 1.6 质谱条件
电喷雾离子源(ESI),正离子模式,喷雾电压3 000 V,蒸发器温度300 ℃,鞘气为10.5 L·min-1,辅助气3 L·min-1,毛细管温度270 ℃,碰撞气为氩气(0.2 Pa),选择反应监测(SRM)扫描模式。SRM模式各离子信息详见表 2。
表 2 亚甲基蓝及代谢物的定性定量特征离子及优化的质谱参数Table 2. Qualitative and quantitative characteristic ions and optimized MS/MS parameters of methylene blue and metabolites标准物
compounds母离子/(m/z)
mother ion子离子/(m/z)
daughter ion碰撞能量/V
collision energy亚甲基蓝 MB 284 268.2* 33 252.1 52 天青A AzuA 256 214.2* 33 199.1 42 天青B AzuB 270 254.1* 35 255.2 28 天青C AzuC 242 200.1* 34 227.2 27 注:*.定量离子
Note: *.MS/MS quantitative ion2. 结果与讨论
2.1 条件优化研究
2.1.1 提取溶液的选择
提取时体系中加入适量浓度的盐酸羟胺溶液可维持还原环境[15],使亚甲基蓝及其代谢物结构保持稳定;同时,对甲苯磺酸作为离子对试剂,可与离子型化合物结合成为分子型物质[15],更有利于亚甲基蓝及其代谢物的提取。向空白阴性草鱼样品中添加200 μL质量浓度为100 μg·L-1的4种混和标准溶液,涡旋混匀,放置30 min后分别使用3.0、4.5、5.5、7.0、10.0等5个不同pH的乙酸铵缓冲液提取,每组4个平行,分别测定每次提取所得亚甲基蓝及其代谢物的回收率,采用pH 4.5的乙酸铵缓冲液提取时亚甲基蓝及其代谢物回收率最高(表 3)。
表 3 不同pH值的提取液提取时亚甲基蓝及代谢物的回收率Table 3. Recovery of methylene blue and metabolites extracting in different pH level% 目标物质
target compoundpH 3.0 4.5 5.5 7.0 10.0 亚甲基蓝 MB 80.63 87.38 78.69 73.94 68.13 天青A AzuA 76.32 91.58 83.12 68.35 70.33 天青B AzuB 79.36 90.19 80.39 48.10 77.31 天青C AzuC 78.98 86.71 81.55 57.31 68.96 2.1.2 提取次数的选择
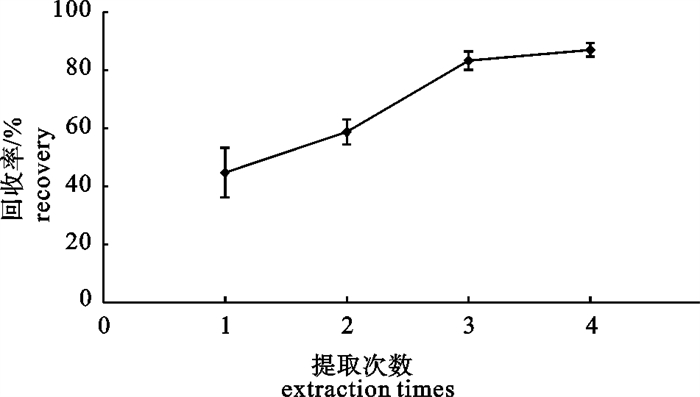
分别使用10 mL的乙腈提取1、2、3、4次,每次4个平行样品,分别测定提取所得亚甲基蓝及其代谢物的回收率,提取1次时亚甲基蓝及其代谢物回收率仅有50%左右,随着提取次数的增多、亚甲基蓝及其代谢物的回收率提高,提取3次和4次的差别不大(图 1),但综合考虑检测成本和时效性,最终选用提取3次。
![]() 图 1 不同提取次数对亚甲基蓝及代谢物回收率的影响Figure 1. Recovery of methylene blue and metabolites under different extraction times
图 1 不同提取次数对亚甲基蓝及代谢物回收率的影响Figure 1. Recovery of methylene blue and metabolites under different extraction times2.1.3 SPE净化小柱的选择
在试管中将200 μL 100 μg·L-1的标准溶液用乙腈稀释到3 mL,加到活化好的对丙磺酸小柱(PRS)、混合型阳离子交换小柱(MCX)、C8/SCX混合模式小柱(MCAX)、C18小柱(C18)4种固相萃取柱上,用3 mL乙腈重复洗涤试管2次,用体积分数为50%的乙酸铵(1 mol·L-1)乙腈洗脱,每组4个平行,考察了不同SPE小柱对亚甲基蓝及其代谢物提取效率的影响。表 4显示MCX过柱速度太快,难以控制流速,目标物损失严重,亚甲基蓝及其代谢物回收率不到40%;C18柱净化效果不理想,过柱后仍有许多杂质;MCAX和PRS柱对目标物质回收率相似,但PRS柱萃取过程中样液流速适中,更易于控制,且加标回收率更稳定,所以采用PRS柱。
表 4 不同的净化小柱对亚甲基蓝及代谢物回收率的影响Table 4. Recovery of methylene blue and metabolites under different solid phase extraction columns% 目标物质
target compounds丙磺酸小柱
PRS column混合型阳离子交换小柱
MCX columnC8/SCX混合模式小柱
MCAX columnC18小柱
C18 column亚甲基蓝 MB 93.2 36.8 90.1 68.3 天青A AzuA 90.3 35.1 80.3 59.2 天青B AzuB 89.2 30.5 87.5 70.5 天青C AzuC 87.1 33.7 79.2 66.9 2.1.4 洗脱剂的选择
在试管中将200 μL 100 μg·L-1的标准溶液用乙腈稀释到3 mL,加到活化好的对丙磺酸小柱(PRS)上,用3 mL乙腈重复洗涤试管2次,分别用甲醇、乙腈、体积分数为50%的乙酸铵(1 mol·L-1)乙腈、体积分数为50%的乙酸铵(1 mol·L-1)甲醇洗脱。结果显示,纯的有机溶剂如甲醇、乙腈等不能将目标物洗脱下来,乙酸铵和甲醇混合液洗脱时仅有部分洗脱下来;采用乙酸铵和乙腈等体积混合洗脱效率明显增强,目标物质峰型良好。此外,试验还考察了采用不同浓度的乙酸铵溶液洗脱对亚甲基蓝及其代谢物回收率的影响(表 5)。结果表明,采用浓度为1 mol·L-1的乙酸铵溶液和等体积的乙腈混和时,亚甲基蓝及其代谢物回收率最高。将洗脱后的溶液稀释到10 mL再进样,使样品中的盐浓度降低,可起到保护质谱仪的作用。
表 5 不同浓度的乙酸铵乙腈洗脱时亚甲基蓝及代谢物的回收率Table 5. Recovery of methylene blue and metabolites with different concentration of eluents% 目标物质
target compoundsc(乙酸铵)/ mol·L-1
ammonium acetate content0.1 0.5 1.0 亚甲基蓝 MB 40.38 73.94 98.13 天青A AzuA 31.58 68.35 95.33 天青B AzuB 30.19 68.10 97.31 天青C AzuC 36.71 87.31 95.96 2.1.5 色谱条件的优化及选择
此试验分别采用5 mmol·L-1乙酸铵-甲醇、5 mmol·L-1乙酸铵-乙腈、0.2%甲酸-甲醇、0.2%甲酸-乙腈4种流动相试验。结果表明,采用0.2%甲酸溶液和甲醇作为流动相时亚甲基蓝及其代谢物峰型最好。甲酸溶液与甲醇的比例不同,对亚甲基蓝及代谢物出峰时间和强度存在明显差异。当流动相中甲醇比例较高时,亚甲基蓝及其代谢物在色谱柱上的保留时间短,峰形也较差;随着甲酸溶液比例增高,亚甲基蓝及其代谢物的出峰时间推迟,且峰形变好。最后形成的梯度洗脱程序见表 1。
2.1.6 质谱条件的优化及确定
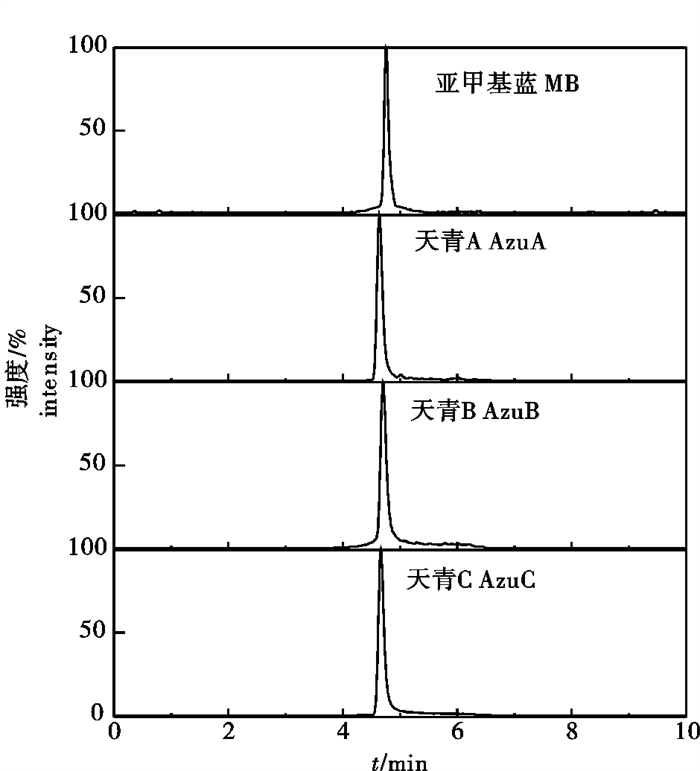
取质量浓度为10 mg·L-1的亚甲基蓝及天青标准品用质谱仪检测,采用注射泵以5 μL·min-1的速度进样,调节毛细管电压和碰撞电压,探索亚甲基蓝及代谢物标准品最佳质谱条件。根据亚甲基蓝及天青的结构特征选择正离子模式,优化喷雾电压、辅助气和鞘气等质谱参数,获得亚甲基蓝和天青稳定的分子离子峰,然后以氩气为碰撞气,获得碎片离子的质谱信息,并进一步优化碰撞能量、毛细管温度和蒸发器温度等质谱参数,使碎片离子信号强度达到最大。最后选择相对丰度最高2个离子作为定量和定性离子,第一强的碎片离子作为定量离子,次强的作为定性离子。亚甲基蓝及代谢物的定性与定量特征离子及优化的质谱参数见表 2。优化条件下各种目标物的SRM图见图 2。
![]() 图 2 优化条件下的亚甲基蓝及天青A、天青B、天青C的SRM图Figure 2. SRM mass spectrum of methylene blue and metabolites under optimized conditions
图 2 优化条件下的亚甲基蓝及天青A、天青B、天青C的SRM图Figure 2. SRM mass spectrum of methylene blue and metabolites under optimized conditions2.2 方法学研究
2.2.1 方法的线性范围、检出限、定量限
准确称取4种标准品各5.0 mg,分别以甲醇溶解并定容至100 mL,其质量浓度均为50 mg·L-1的单标储备液。采用逐级稀释的方式,配制成质量浓度为0.50 μg·L-1、2.00 μg·L-1、5.00 μg·L-1、10.00 μg·L-1、50.00 μg·L-1、100.00 μg·L-1的混合标准工作液,按照质量浓度由低到高进样,对各目标物峰面积及其质量浓度进行线性回归分析,得到亚甲基蓝及其代谢物的标样工作曲线。结果表明,4种目标物在0.50~100.00 μg·L-1范围内呈良好的线性关系,相关系数R>0.99。检出限(LOD)采用向空白样品中逐级降低加标浓度的方法来确定,以大于等于3倍信噪比(S/N≥3)对应的目标物浓度为检出限,以S/N≥10对应的浓度为定量限(表 6)。
表 6 0.50~100.00μg·L-1范围内亚甲基蓝及其代谢物的线性关系及相关系数Table 6. Linear relationship and correlation coefficient of methylene blue and metabolites within scope 0.50~100.00 μg·L-1物质
compound线性方程
linear relationship相关系数
R检出限/μg·kg-1
LOD定量限/μg·kg-1
LOQ亚甲基蓝 MB y=45 254x+140 368 0.997 9 0.40 2.00 天青A AzuA y=3 308.3x+16 774 0.995 9 0.60 2.00 天青B AzuB y=49 098x+167 560 0.998 0 0.50 2.00 天青C AzuC y=13 223x+30 124 0.995 3 0.40 2.00 注:y.亚甲基蓝及其代谢物的定量离子的质谱峰面积;x.亚甲基蓝及其代谢物的质量浓度(μg·L-1)
Note: y. mass spectra peak area of methylene blue and metabolites; x. content of methylene blue and metabolites(μg·L-1)2.2.2 样品加标的回收率和精密度
选取空白草鱼、凡纳滨对虾和中华绒螯蟹3种类型的水产品,分别添加2 μg·kg-1、4 μg·kg-1、10 μg·kg-13个水平的亚甲基蓝及其代谢物标准品,按1.4、1.5、1.6所述方法进行加标回收率测定,每个加标水平平行测定6次,计算平均回收率和相对标准偏差。4种目标物的加标回收率为74.23%~94.40%,在各个加标水平下加标回收率比较稳定,相对标准偏差(RSD)为1.13%~10.28%(表 7~表 9),可满足水产品中亚甲基蓝及代谢物残留的检测要求。
表 7 空白草鱼肉样中亚甲基蓝及代谢物的加标回收率和精密度(n=6)Table 7. Recovery and precision of methylene blue and azures of blank Ctenoparyngodon idellus samples目标物质 target compound 加标量/μg·kg-1 added amount 回收率/% recovery 相对标准偏差/% RSD 亚甲基蓝 MB 2 83.10 1.13 4 81.59 4.09 10 88.35 3.97 天青A AzuA 2 85.47 9.41 4 81.75 5.03 10 87.68 9.76 天青B AzuB 2 82.94 2.53 4 89.65 5.83 10 88.56 8.03 天青C AzuC 2 87.91 9.72 4 90.69 3.61 10 92.60 5.98 表 8 空白凡纳滨对虾样品中亚甲基蓝及代谢物的加标回收率和精密度(n=6)Table 8. Recovery and precision of methylene blue and azures of blank Penaeus vannamei samples目标物质 target compound 加标量/μg·kg-1 added amount 回收率/% recovery 相对标准偏差/% RSD 亚甲基蓝 MB 2 87.94 5.46 4 89.42 4.91 10 80.80 1.46 天青A AzuA 2 78.11 8.97 4 84.86 9.81 10 82.20 6.06 天青B AzuB 2 80.01 5.53 4 83.62 8.58 10 90.83 5.24 天青C AzuC 2 91.49 8.69 4 94.40 5.21 10 92.86 4.10 表 9 空白中华绒螯蟹样品中亚甲基蓝及代谢物的加标回收率和精密度(n=6)Table 9. Recovery and precision of methylene blue and azures of blank Eriocheir sinensis samples目标物质 target compound 加标量/μg·kg-1 added amount 回收率/% recovery 相对标准偏差/% RSD 亚甲基蓝 MB 2 81.85 5.42 4 80.35 6.22 10 81.68 4.20 天青A AzuA 2 74.54 8.75 4 81.05 8.25 10 90.42 9.74 天青B AzuB 2 74.23 6.83 4 85.24 7.84 10 87.05 10.28 天青C AzuC 2 86.69 2.59 4 86.21 6.50 10 88.50 4.24 3. 结论
建立了超高效液相色谱-串联质谱法分析水产品中亚甲基蓝及其代谢物残留量的方法。样品用乙腈提取,经PRS柱固相萃取,乙酸铵乙腈洗脱,以甲醇和0.2%甲酸为流动相进行梯度洗脱分离,质谱条件采用电喷雾离子源(ESI),正离子模式,选择反应监测(SRM)的扫描模式定量分析。该方法准确度高、实用性强、简便易行,能满足实际样品中亚甲基蓝及其代谢物的检测要求,也可作为亚甲基蓝其他方面研究的参考方法。
-
![]()
图 1 不同提取次数对亚甲基蓝及代谢物回收率的影响
Figure 1. Recovery of methylene blue and metabolites under different extraction times
![]()
图 2 优化条件下的亚甲基蓝及天青A、天青B、天青C的SRM图
Figure 2. SRM mass spectrum of methylene blue and metabolites under optimized conditions
表 1 流动相梯度洗脱程序
Table 1 Gradient elution with mobile phases
t/min 0.2%甲酸(A)
0.2% methanoic acid甲醇(B)
methanol0.0 90 10 1.0 90 10 5.0 10 90 5.1 90 10 9.0 90 10  下载: 导出CSV
下载: 导出CSV
表 2 亚甲基蓝及代谢物的定性定量特征离子及优化的质谱参数
Table 2 Qualitative and quantitative characteristic ions and optimized MS/MS parameters of methylene blue and metabolites
标准物
compounds母离子/(m/z)
mother ion子离子/(m/z)
daughter ion碰撞能量/V
collision energy亚甲基蓝 MB 284 268.2* 33 252.1 52 天青A AzuA 256 214.2* 33 199.1 42 天青B AzuB 270 254.1* 35 255.2 28 天青C AzuC 242 200.1* 34 227.2 27 注:*.定量离子
Note: *.MS/MS quantitative ion
下载: 导出CSV
表 3 不同pH值的提取液提取时亚甲基蓝及代谢物的回收率
Table 3 Recovery of methylene blue and metabolites extracting in different pH level
% 目标物质
target compoundpH 3.0 4.5 5.5 7.0 10.0 亚甲基蓝 MB 80.63 87.38 78.69 73.94 68.13 天青A AzuA 76.32 91.58 83.12 68.35 70.33 天青B AzuB 79.36 90.19 80.39 48.10 77.31 天青C AzuC 78.98 86.71 81.55 57.31 68.96
下载: 导出CSV
表 4 不同的净化小柱对亚甲基蓝及代谢物回收率的影响
Table 4 Recovery of methylene blue and metabolites under different solid phase extraction columns
% 目标物质
target compounds丙磺酸小柱
PRS column混合型阳离子交换小柱
MCX columnC8/SCX混合模式小柱
MCAX columnC18小柱
C18 column亚甲基蓝 MB 93.2 36.8 90.1 68.3 天青A AzuA 90.3 35.1 80.3 59.2 天青B AzuB 89.2 30.5 87.5 70.5 天青C AzuC 87.1 33.7 79.2 66.9
下载: 导出CSV
表 5 不同浓度的乙酸铵乙腈洗脱时亚甲基蓝及代谢物的回收率
Table 5 Recovery of methylene blue and metabolites with different concentration of eluents
% 目标物质
target compoundsc(乙酸铵)/ mol·L-1
ammonium acetate content0.1 0.5 1.0 亚甲基蓝 MB 40.38 73.94 98.13 天青A AzuA 31.58 68.35 95.33 天青B AzuB 30.19 68.10 97.31 天青C AzuC 36.71 87.31 95.96
下载: 导出CSV
表 6 0.50~100.00μg·L-1范围内亚甲基蓝及其代谢物的线性关系及相关系数
Table 6 Linear relationship and correlation coefficient of methylene blue and metabolites within scope 0.50~100.00 μg·L-1
物质
compound线性方程
linear relationship相关系数
R检出限/μg·kg-1
LOD定量限/μg·kg-1
LOQ亚甲基蓝 MB y=45 254x+140 368 0.997 9 0.40 2.00 天青A AzuA y=3 308.3x+16 774 0.995 9 0.60 2.00 天青B AzuB y=49 098x+167 560 0.998 0 0.50 2.00 天青C AzuC y=13 223x+30 124 0.995 3 0.40 2.00 注:y.亚甲基蓝及其代谢物的定量离子的质谱峰面积;x.亚甲基蓝及其代谢物的质量浓度(μg·L-1)
Note: y. mass spectra peak area of methylene blue and metabolites; x. content of methylene blue and metabolites(μg·L-1)
下载: 导出CSV
表 7 空白草鱼肉样中亚甲基蓝及代谢物的加标回收率和精密度(n=6)
Table 7 Recovery and precision of methylene blue and azures of blank Ctenoparyngodon idellus samples
目标物质 target compound 加标量/μg·kg-1 added amount 回收率/% recovery 相对标准偏差/% RSD 亚甲基蓝 MB 2 83.10 1.13 4 81.59 4.09 10 88.35 3.97 天青A AzuA 2 85.47 9.41 4 81.75 5.03 10 87.68 9.76 天青B AzuB 2 82.94 2.53 4 89.65 5.83 10 88.56 8.03 天青C AzuC 2 87.91 9.72 4 90.69 3.61 10 92.60 5.98
下载: 导出CSV
表 8 空白凡纳滨对虾样品中亚甲基蓝及代谢物的加标回收率和精密度(n=6)
Table 8 Recovery and precision of methylene blue and azures of blank Penaeus vannamei samples
目标物质 target compound 加标量/μg·kg-1 added amount 回收率/% recovery 相对标准偏差/% RSD 亚甲基蓝 MB 2 87.94 5.46 4 89.42 4.91 10 80.80 1.46 天青A AzuA 2 78.11 8.97 4 84.86 9.81 10 82.20 6.06 天青B AzuB 2 80.01 5.53 4 83.62 8.58 10 90.83 5.24 天青C AzuC 2 91.49 8.69 4 94.40 5.21 10 92.86 4.10
下载: 导出CSV
表 9 空白中华绒螯蟹样品中亚甲基蓝及代谢物的加标回收率和精密度(n=6)
Table 9 Recovery and precision of methylene blue and azures of blank Eriocheir sinensis samples
目标物质 target compound 加标量/μg·kg-1 added amount 回收率/% recovery 相对标准偏差/% RSD 亚甲基蓝 MB 2 81.85 5.42 4 80.35 6.22 10 81.68 4.20 天青A AzuA 2 74.54 8.75 4 81.05 8.25 10 90.42 9.74 天青B AzuB 2 74.23 6.83 4 85.24 7.84 10 87.05 10.28 天青C AzuC 2 86.69 2.59 4 86.21 6.50 10 88.50 4.24
下载: 导出CSV
-
[1] 李悦悦, 林云萍, 林诗燕, 等. 孔雀绿、福尔马林、亚甲基蓝对河蟹离体胚胎真菌病防治效果的比较[J]. 水产科技情报, 1998, 25 (2): 77-81. https://d.wanfangdata.com.cn/periodical/QK199800484055 [2] 梁华芳. 凡纳对虾溞状幼体黏脏病的治疗[J]. 海洋湖沼通报, 2004 (1): 28-31. doi: 10.3969/j.issn.1003-6482.2004.01.005 [3] 杨方, 刘正才, 胡小钟, 等. 噻嗪类染料在动物体内的代谢及检测研究进展[J]. 药物分析杂志, 2008, 2 (8): 1395-1399. https://www.cqvip.com/QK/95650X/20088/28125655.html [4] CHUNG K T, FULK G E, ANDREWS A W. Mutagenicity testing of some commonly used dyes[J]. Appl Environ Microbiol, 1981, 42 (4): 641-648. doi: 10.1128/aem.42.4.641-648.1981
[5] ZIV G, HEAVNER J E. Permeability of the blood-milk barrier to methylene blue in cows and goats[J]. J Vet Pharmacol Ther, 1984, 7 (1): 55-59. doi: 10.1111/j.1365-2885.1984.tb00879.x
[6] CODDINGTON C C, ANDERSON T L, ACCETTA C R, et al. Adverse effects of methylene blue on human sperm motility, components of human reproductive tract fluids, and mouse embryo cleavage[J]. Fertil Steril, 1989, 51(3): 480-485. doi: 10.1016/S0015-0282(16)60558-7
[7] BROWNSTEIN S, LISZAUER A D, JACKSON W B. Ocular complications of a topical methylene blue-vasoconstrictor-anesthetic preparation[J]. Can J Ophthalmol, 1989, 24 (7): 317-324. https://pubmed.ncbi.nlm.nih.gov/2624913/
[8] LIU J Y, PENG S F. Acute toxicity of methylene blue to zoea mysis and post larvae of Penaeus vannamei[J]. Trans Oceanol Limnol, 2005 (1): 49-53. https://www.sciencedirect.com/science/article/pii/004484868890333X
[9] BELAZ-DAVID N, DECOSTERD L A, APPENZELLER M, et al. Spectrophotometric determination of methylene blue in biological fluids after ion-pair extraction and evidence of its adsorption on plastic polymers[J]. Eur J Pharm Sci, 1997, 5 (6): 335-345. doi: 10.1016/S0928-0987(97)00061-4
[10] BORWITZKY H, HAEFELI W E, BURHENNE J. Analysis of methylene blue in human urine by capillary electrophoresis[J]. J Chromatogr B: Analyt Technol Biomed Life Sci, 2005, 826 (1/2): 244-251. doi: 10.1016/j.jchromb.2005.09.013
[11] 宫向红, 徐英江, 任传博, 等. HPLC测定水产品中孔雀石绿、亚甲基蓝、结晶紫及其代谢物的残留量[J]. 食品科学, 2012, 33 (4): 144-147. https://www.cqvip.com/qk/89028x/201203/41251927.html [12] 吴艳兵, 王建华, 李广领, 等. 高效液相色谱法检测水产品中亚甲基蓝的残留[J]. 湖南农业科学, 2008(3): 165-167. doi: 10.3969/j.issn.1006-060X.2008.03.059 [13] 杨方, 范克伟, 刘正才, 等. 液相色谱法检测水产品中亚甲蓝及其代谢物残留[J]. 分析化学, 2009, 37 (4): 517-521. doi: 10.3321/j.issn:0253-3820.2009.04.008 [14] 李邦瑞. 高效液相色谱-串联质谱联用法在食品安全分析中的应用[D]. 长沙: 湖南大学, 2009. 10.7666/d.y1723965 [15] 王丽敏, 杜雪莉, 赵艳, 等. 超高效液相色谱-串联质谱快速测定水产品中孔雀石绿、结晶紫及其代谢物残留量[J]. 南方水产, 2010, 6 (4): 32-36. doi: 10.3969/j.issn.1673-2227.2010.04.006 [16] 朱世超, 钱卓真, 吴成业. 水产品中7种大环内酯类抗生素残留量的测定HPLC-MS/MS法[J]. 南方水产科学, 2012, 8 (1): 54-60. doi: 10.3969/j.issn.2095-0780.2012.01.009 [17] 赵东豪, 黎智广, 黄珂, 等. 高效液相色谱-串联质谱法检测水产品中三聚氰胺残留的比较研究[J]. 南方水产, 2010, 6 (3): 32-35. doi: 10.3969/j.issn.1673-2227.2010.03.006 [18] 张海琪, 何中央, 郑重莺, 等. 液相色谱-串联质谱法测定水产品中孔雀石绿的残留量[J]. 南方水产, 2007, 3 (6): 14-21. doi: 10.3969/j.issn.2095-0780.2007.06.003 -
期刊类型引用(10)
1. 孙良娟,李红权,蔡润斌,庄姜云,唐庆强,唐媛媛,钟键,黄武. 高效液相色谱-串联质谱法测定水产品中9种禁用染料类药物残留. 分析测试学报. 2023(01): 89-95 .  百度学术
百度学术
2. 李涛,贾立平,李锋刚,王绿洲. 亚甲基蓝在鲤体内消除规律研究. 水产科技情报. 2022(02): 99-105 . 百度学术
3. 黄宣运,杨光昕,孔聪,黄冬梅,蔡友琼. 间接竞争酶联免疫法测定鱼体中的亚甲基蓝. 食品安全质量检测学报. 2020(22): 8235-8239 . 百度学术
4. 余玮玥,黄冬梅,史永富,孔聪,田良良,韩峰,张政权. 液相色谱-串联质谱法测定水产品中亮绿、亚甲基蓝及其代谢物残留量. 分析测试学报. 2019(01): 58-63 . 百度学术
5. 施雅. 淡水鱼中亚甲基蓝、孔雀石绿类药物残留检测. 广州化工. 2019(14): 131-133 . 百度学术
6. 高平,杨曦,莫彩娜,陈日檬,曾丹丹,刘唤明,洪鹏志,周凯,陈营寿. 通过式固相萃取净化/高效液相色谱-串联质谱法快速测定水产品中6种麻醉剂残留. 分析测试学报. 2019(09): 1059-1065 . 百度学术
7. 杨卫军,张静余,严敏鸣. 亚甲基蓝检测方法及代谢动力学研究进展. 食品安全质量检测学报. 2018(10): 2419-2425 . 百度学术
8. Jin Fan,Zhi-Hai Xie,Xiao-Xiao Teng,Yu Zhang. Determination of methylene blue by resonance light scattering method using silica nanoparticles as probe. Chinese Chemical Letters. 2017(05): 1104-1110 . 必应学术
9. 梁楠,胡鲲,刘腾飞,吴映捷,杨先乐. 亚甲基蓝及其代谢物在异育银鲫体内分布及消除规律的研究. 南方水产科学. 2017(01): 19-25 . 本站查看
10. 龙举,李子孟,喻亮,李佩佩,严忠雍,张小军. 高效液相色谱法测定养殖水体中的亚甲基蓝. 安徽农业科学. 2015(24): 5-6+16 . 百度学术
其他类型引用(5)
计量
- 文章访问数: 3482
- HTML全文浏览量: 146
- PDF下载量: 2060
- 被引次数: 15
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
