
Study on fermentation conditions for oceanic red yeast strain RH1
-
摘要:
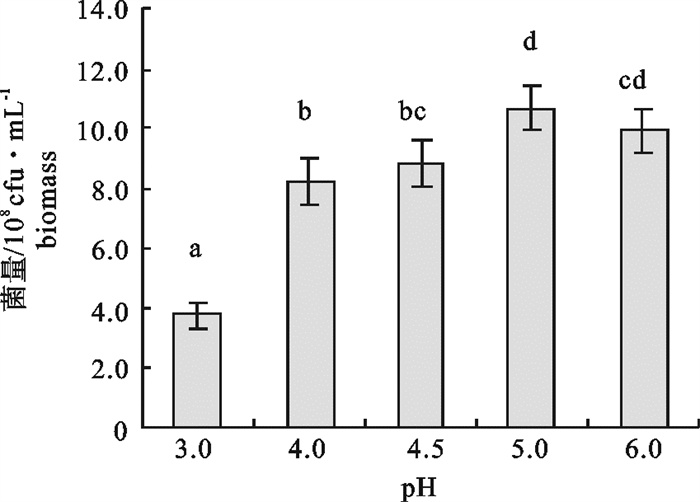
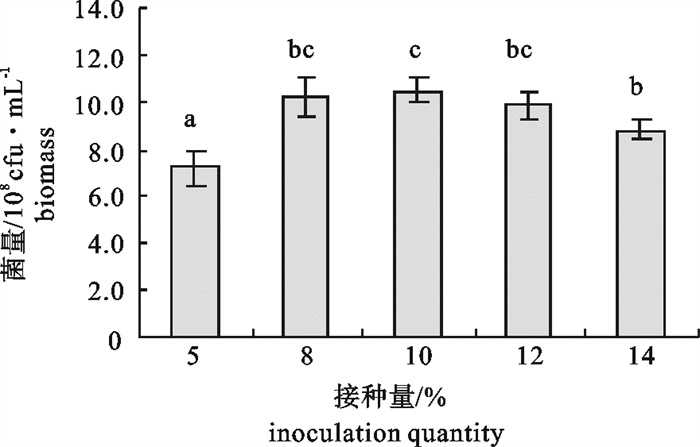
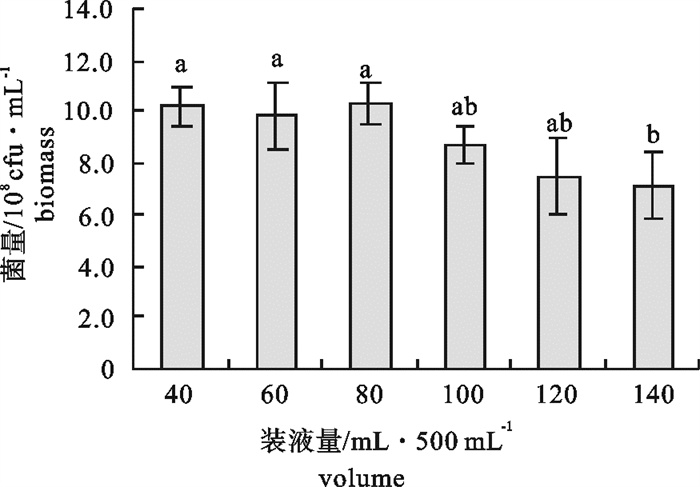
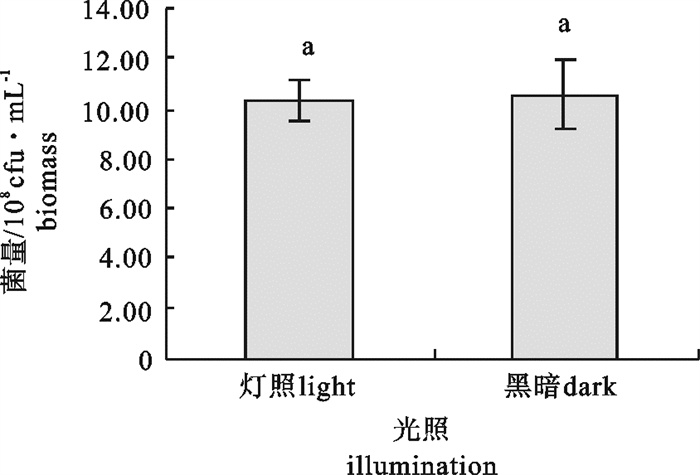
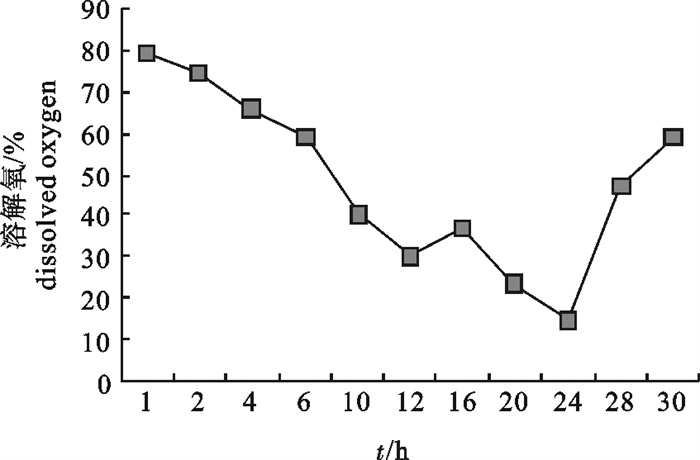
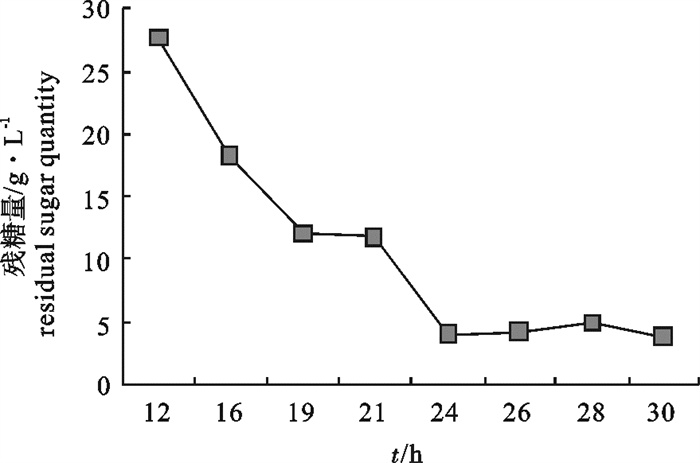
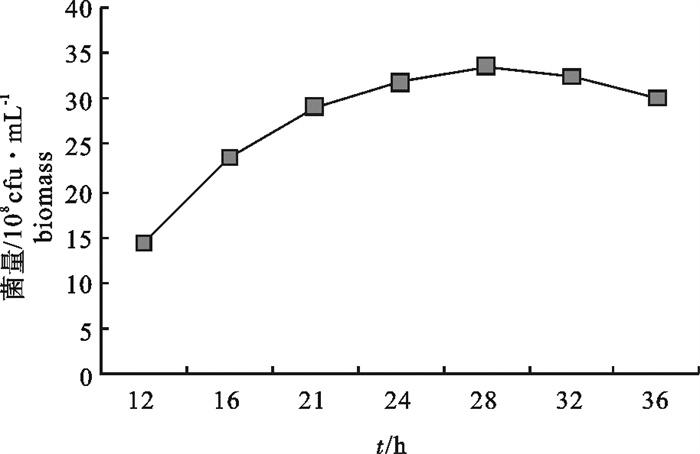
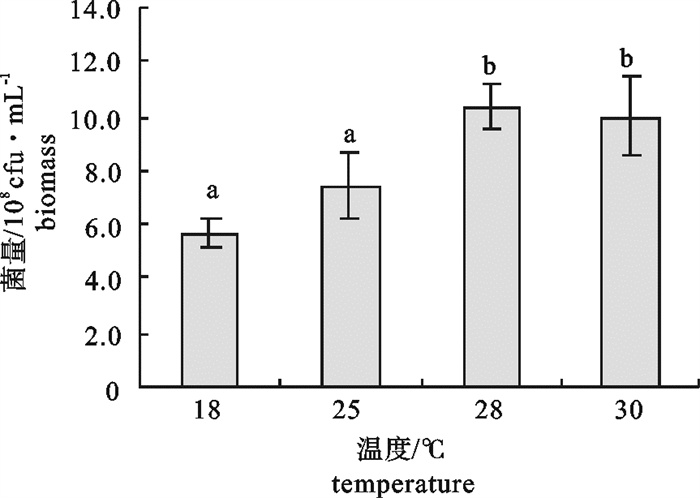
该研究比较了不同碳源、氮源、无机盐对海洋红酵母菌(Rhodotorula sp.)RH1菌株发酵产量的影响,通过葡萄糖、蛋白胨、酵母膏和硫酸镁(MgSO4)4因素3水平的正交试验,确定了RH1菌株的最优培养基为蛋白胨10 g · L-1,酵母膏15 g · L-1,葡萄糖20 g · L-1,MgSO4 0.25 g · L-1,磷酸二氢钾(KH2PO4)0.25 g · L-1,氯化钠(NaCl)10 g · L-1。结果显示,该菌株最佳摇瓶发酵条件为接种量10%,初始pH 5.0,摇瓶装液量80 mL/500 mL三角瓶,培养温度28 ℃,经48 h培养,菌量可达10.46×108 cfu · mL-1,比优化条件前提高23.9%。还进行了RH1菌株25 L发酵罐扩大培养试验,在接种量为8%、初始pH 5.0、搅拌速率350~400 r · min-1、通气量9 L ·min-1、温度28 ℃的条件下,经28 h的培养,菌量可达33.6×108 cfu · mL-1。预计通过连续补料的方式进行培养,有望进一步提高菌量。
Abstract:We compared the effects of different carbon sources, nitrogen sources and inorganic salts on fermentation yield of oceanic red yeast (Rhodotorula sp.)RH1. By 4 factors and 3 levels orthogonal test, the optimum fermentation conditions for RH1 were determined: peptone 10 g · L-1, yeast extract 15 g · L-1, glucose 20 g · L-1, MgSO4 0.25 g · L-1, KH2PO4 0.25 g · L-1, NaCl 10 g · L-1, inoculation amount 10%, initial pH 5.0, temperature 28 ℃, 80 mL medium in 500 mL triangle bottle, and fermentation time 48 h. Under above conditions, the cell biomass of RH1 can reach 10.46×108 cfu · mL-1, which increases by 23.9% as compared with the fermentation before optimization. An experiment of expanded culture is also conducted in a 25 L fermentation tank. The cell biomass of RH1 increased to 33.6×108 cfu · mL-1 in 28 h under the conditions of inoculation amount 8%, initial pH 5.0, revolution speed 350~400 r · min-1, air ventilation volume 9 L · min-1 and temperature 28 ℃. It is expected to further increase the biomass of strain RH1 by continuous filling of the materials.
-
Keywords:
- Rhodotorula sp. /
- culture conditions /
- optimization
-
全球气候变化对生态系统产生深远影响,制约着物种的生态习性和地理分布,是影响海洋生物分布格局的重要因素之一[1]。随着气候变化引起的海水酸化和温度上升等进程的持续,大多数海洋生物的新陈代谢、个体发育和生活史特征等在一定程度上受到了影响,导致群落结构发生变化,同时分布格局也随气候变化产生生境迁移[2-4]。这些改变可能导致生物面临种群数量显著下降或加速灭绝的风险,进而引起生态系统结构和功能紊乱、生物多样性丧失[5]。作为海洋生物的典型代表,长棘海星 (Acanthaster planci) 和褐拟鳞鲀 (Balistoides viridescens) 对气候变化的响应受到了广泛关注。了解它们对全球气候变化的响应并确定影响其生境分布的关键因素,对维持生物多样性的稳定以及珊瑚礁生态系统保护与恢复管理措施的制定具有至关重要的作用。
长棘海星隶属于棘皮动物门、海星纲、有棘目、长棘海星科、长棘海星属,分布于印度-太平洋热带珊瑚礁海域[6],是著名的“珊瑚杀手”。据统计,1963年以来澳大利亚大堡礁共经历了3次长棘海星大规模暴发,导致大量珊瑚白化,珊瑚礁生态系统严重退化、生物多样性显著降低[7]。1985—2012年间对珊瑚礁的广泛调查发现,全球珊瑚礁的活珊瑚覆盖率从28.0%下降至13.8%,其中由长棘海星造成的损害高达42%,对珊瑚礁生态系统造成了严重危害[8]。由于人类对其天敌 [如法螺 (Charonia tritonis) 等] 的过度捕捞、营养盐大量流入等影响,导致长棘海星易出现周期性暴发[9]。了解长棘海星在未来不同气候情景下的潜在分布变化可为保护和修复珊瑚礁生态系统提供理论依据和科学指导。遗憾的是我国对于长棘海星的研究仍处于起步阶段,相关研究鲜有报道。褐拟鳞鲀是长棘海星的重要捕食者,隶属于辐鳍鱼纲、鲀形目、鳞鲀科,分布在热带和温带的淡水和海洋环境中[10],具有非常高的经济价值。在印度洋-太平洋的海域中,褐拟鳞鲀会捕食成年长棘海星,与长棘海星的种群动态存在一定关联[11]。此外,气候变化对两个物种潜在适宜生境分布的影响也尚未得到评估。
物种分布模型 (Species distribution models, SDMs) 是研究气候变化对物种潜在生境分布格局影响及生物多样性响应的有效工具[12]。随着统计学和生态学理论的不断完善及计算机应用技术的深入发展,SDMs技术也得到了改进,并广泛应用于陆地、淡水和海洋物种的地理分布预测、生境气候适宜性评价及野生资源保护与管理等方面[13]。Feitosa等[14]利用SDMs并结合公开数据对小尾真鲨 (Carcharhinus porosus) 当前和未来的潜在分布做出预测,确定南美北部海岸为保护该物种最有潜力的区域;Zhang等[15]利用SDMs预测了未来不同情景下中国、韩国和日本沿海的日本少鳞鱚 (Sillago japonica) 的潜在生境分布情况,指出未来管理战略需考虑其适宜生境范围的缩小及北移,为制定管理策略提供了重要指导。可见,SDMs在模拟未来气候变化情景下物种的生境变化具有广泛应用。
目前关于长棘海星的研究主要集中在其生物累积与分布特征、暴发原因探讨、危害与防治等方面[16-17],也有少数利用单一物种分布模型对长棘海星的研究[18]。而褐拟鳞鲀的相关报道则集中在其生物学特征与遗传方面[10,19]。本研究结合长棘海星、褐拟鳞鲀的现有分布数据,利用组合物种分布模型对两者的适宜生境分布进行模拟,揭示影响分布的关键环境因子,并绘制两个物种当前和未来不同气候情景下的适宜生境,为维持珊瑚礁生态可持续发展及制定未来气候适应管理决策提供科学参考。
1. 材料与方法
1.1 物种分布数据
本研究主要针对长棘海星及其捕食者褐拟鳞鲀在全球范围内的分布情况。通过查询全球生物多样性信息网络 (Global biodiversity information facility, GBIF, http://www.Gbif.org) 获取2000年以来长棘海星及褐拟鳞鲀的发生记录。为减少采样带来的偏差并与环境数据分辨率相匹配,筛选掉重复及坐标不精确的样本,保证每个栅格数据内随机选取1个样本,最终获得1 094个长棘海星和1 793个褐拟鳞鲀的数据用于模型构建 (图1)。建模过程中部分算法有时需要环境条件来与发现物种的环境进行对比,需要物种的实际和缺席分布数据[20]。由于缺乏两物种不存在点数据,因此运行模型时在全球地理范围内随机生成10 000个坐标点作为伪不存在点,以提高模型性能。
1.2 环境变量
综合考虑数据可用性和环境相关性,本研究以全球海洋环境数据为分析依据。当前和未来 (2040—2050年) 的海洋遥感数据来源于BIO-Oracle v2.1 (https://bio-oracle.org/) 数据库及Global Marine Environment Datasets数据库 (http://gmed.auckland.ac.nz/index.html)。共下载57个环境变量,经过处理后空间分辨率为0.083 333 333。考虑到环境因子间的自相关性会影响模型的准确度,因此计算每对变量间的Pearson相关系数,若 >|0.7| 则认为相关性较强,保留其中1个变量[21],最终得到表1的12个环境变量。未来海洋环境数据的科学假设来自于4种排放情景,即“典型浓度目标 (Representative concentration pathways, RCP)”,分别代表不同CO2排放标准下的未来情景,包括RCP26 (最低温室气体排放)、RCP45 (中等温室气体排放)、RCP60 (较高温室气体排放) 和RCP85 (最高温室气体排放)[22]。本研究选择RCP26、RCP60和RCP85情景来模拟未来环境,用对应变量建模并对未来气候进行投影。
表 1 环境变量说明Table 1. Description of environmental variables变量 Variable 单位 Unit 最大叶绿素质量浓度 Maximum chlorophyll mg·m−3 最大流速 Maximum current velocity m·s−1 最小流速 Minimum current velocity m·s−1 平均溶解氧浓度Mean dissolved oxygen mol·m−3 最小铁离子浓度 Minimum iron μmol·m−3 最小硝酸盐浓度 Minimum nitrate mol·m−3 最小浮游动物浓度 Minimum phytoplankton μmol·m−3 最小初级生产力浓度
Minimum primary productivityg·m−3·d−1 平均盐度 Mean salinity ‰ 最小温度 Minimum temperature ℃ 深度 Depth m 离岸距离 Land distance km 1.3 物种分布模型构建
本研究基于R语言biomod2程序包对两个物种当前和未来的潜在分布区进行预测。选取广义线性模型 (Generalized Linear Model, GLM)、推进式回归树模型 (Generalized Boosting Model, GBM)、分类树分析 (Classification Tree Analysis, CTA)、人工神经网络 (Artificial Neural Network, ANN)、表面分布区分室 (Surface Range Envelope, SRE) 模型、柔性判别分析 (Flexible Discriminant Analysis, FDA) 以及随机森林 (Random Forest, RF) 7种单一模型。为构建更多单一模型结果,减小单次模型运行带来的误差,每种模型算法运行10次,模型参数设置为默认。而后对单一模型结果进行集成产生组合模型用于后续的结果分析,以提高模型精度。
1.4 模型评估方法
模型运行时,将数据随机拆分,80%作为训练集样本,20%作为测试集样本,对模型精度进行交叉验证。模型评价使用真实技巧统计 (True skill statistic, TSS) 和受试者操作特征曲线 (Receiver operating curve, ROC) 2个统计学参数[23]。ROC曲线以假正率为横坐标,真正率为纵坐标。TSS值综合敏感度和特异性指标,考虑了疏忽性和替代性的错误,其取值范围为 [0,1],>0.8时拟合效果极好,0.6~0.8拟合效果好,0.4~0.6拟合效果一般[24]。
1.5 模型集成方法
综合考虑所有模型的运算结果,保留TSS和ROC值均 >0.9的单模型构成最终组合模型的结果。采用投票平均 (Committee Averaging, CA) 和概率加权平均 (Weighted Mean of Probabilities, WM) 对所有单一模型进行集成产生组合模型 (Ensemble Model),并对模型的不确定性进行评估[25]。
2. 结果
2.1 模型性能评价
模型交叉验证结果显示,两个物种的模型性能结果存在一定的相似性 (图2),表现最好的模型均为随机森林模型 (RF),长棘海星RF的TSS和ROC值分别为0.96和0.99 (图2-a),褐拟鳞鲀RF的TSS和ROC值分别为0.97和0.99 (图2-b),表明模型的预测精度高,整体性能较优。然而SRE模型表现出了最差的性能 (图2),两物种SRE模型的TSS值均为0.62、ROC值均为0.81,结果显示了不同单一模型间的不确定性。长棘海星与褐拟鳞鲀投票平均和概率加权平均组合模型的TSS、ROC值分别为0.96、0.99与0.97、0.99,评估结果表明,组合模型能够一定程度上减少不同单一模型的不确定性,提高预测的准确性。
![]() 图 2 基于TSS和ROC的7种物种分布模型统计结果注:a. 长棘海星;b. 褐拟鳞鲀;GLM. 广义线性模型;GBM. 推进式回归树模型;CTA. 分类树分析模型;ANN. 人工神经网络模型;SRE. 表面分布区分室模型;FDA. 柔性判别分析模型;RF. 随机森林模型。Figure 2. Results of statistical analyses of seven species distribution models based on analyses of true skill statistics and receiver operating curveNote: a. A. planci; b. B. viridescens; GLM. Generalized Linear Model; GBM. Generalized Boosting Model; CTA. Classification Tree Analysis; ANN. Artificial Neural Network; SRE. Surface Range Envelope; FDA. Flexible Discriminant Analysis; RF. Random Forest.
图 2 基于TSS和ROC的7种物种分布模型统计结果注:a. 长棘海星;b. 褐拟鳞鲀;GLM. 广义线性模型;GBM. 推进式回归树模型;CTA. 分类树分析模型;ANN. 人工神经网络模型;SRE. 表面分布区分室模型;FDA. 柔性判别分析模型;RF. 随机森林模型。Figure 2. Results of statistical analyses of seven species distribution models based on analyses of true skill statistics and receiver operating curveNote: a. A. planci; b. B. viridescens; GLM. Generalized Linear Model; GBM. Generalized Boosting Model; CTA. Classification Tree Analysis; ANN. Artificial Neural Network; SRE. Surface Range Envelope; FDA. Flexible Discriminant Analysis; RF. Random Forest.2.2 环境变量重要性评估
筛选后参与建模的环境变量共12个 (表1),其Pearson相关系数均 <|0.7|,表明变量间的相关性较弱,可提高预测准确度。对于长棘海星,最小温度和离岸距离的重要性最高,两者重要性之和超过70% (表2),表明这两个变量是长棘海星空间分布的主要影响因子。对于褐拟鳞鲀,最小温度、平均溶解氧浓度和离岸距离的重要性最高,三者重要性之和占70%以上,因此,这3个变量可能是决定褐拟鳞鲀空间分布的主要影响因子 (表3)。
表 2 长棘海星影响因子重要性排序结果Table 2. Importance ranking of factors influencing distribution of A. planci影响因子
Environmental factor重要性
Importance/%排序
Sorting最小温度 Minimum temperature 43.34 1 离岸距离 Land distance 28.54 2 最小铁离子浓度 Minimum iron 8.03 3 平均溶解氧浓度 Mean dissolved oxygen 7.61 4 平均盐度 Mean salinity 2.75 5 最小初级生产力浓度
Minimum primary productivity1.90 6 深度 Depth 1.90 6 最小硝酸盐浓度 Minimum nitrate 1.48 8 最大流速 Maximum current velocity 1.27 9 最小浮游动物浓度
Minimum phytoplankton1.27 9 最大叶绿素浓度 Maximum chlorophyll 1.06 11 最小流速 Minimum current velocity 0.85 12 表 3 褐拟鳞鲀影响因子重要性排序结果Table 3. Importance ranking of factors influencing distribution of B. viridescens影响因子
Environmental factor重要性
Importance/%排序
Sorting最小温度 Minimum temperature 48.82 1 平均溶解氧浓度 Mean dissolved oxygen 15.94 2 离岸距离 Land distance 11.81 3 最小铁离子浓度 Minimum iron 8.86 4 平均盐度 Mean salinity 4.13 5 最小初级生产力
Minimum primary productivity2.36 6 最大流速 Maximum current velocity 2.17 7 最小硝酸盐浓度 Minimum nitrate 1.57 8 深度 Depth 1.57 8 最小浮游动物浓度
Minimum phytoplankton1.18 10 最大叶绿素浓度 Maximum chlorophyll 0.98 11 最小流速 Minimum current velocity 0.59 12 环境变量的响应曲线表示的是物种存在概率与对应环境变量之间的关系,可从中解析长棘海星和褐拟鳞鲀在不同环境下的生态位,从而确定其适宜生境。长棘海星的存在概率与温度整体上呈非线性关系,多数模型预测其存在概率随温度增加而逐渐增大并趋于稳定 (图3)。离岸距离响应曲线的结果显示出了模型间的差异,整体而言,离岸距离越小,长棘海星的生境适宜性越高。不同模型的铁离子浓度响应曲线也表现出一定的差异,长棘海星生境适宜性随铁离子浓度的增加而降低 (图3)。
![]() 图 3 预测长棘海星与褐拟鳞鲀发生概率随环境变量的响应曲线注:GLM. 广义线性模型;GBM. 推进式回归树模型;CTA. 分类树分析模型;ANN. 人工神经网络模型;SRE. 表面分布区分室模型;FDA. 柔性判别分析模型;RF. 随机森林模型。Figure 3. Response curves of predicted occurrence probability of A. planci and B. viridescens against environment variablesNote: GLM. Generalized Linear Model; GBM. Generalized Boosting Model; CTA. Classification Tree Analysis; ANN. Artificial Neural Network; SRE. Surface Range Envelope; FDA. Flexible Discriminant Analysis; RF. Random Forest.
图 3 预测长棘海星与褐拟鳞鲀发生概率随环境变量的响应曲线注:GLM. 广义线性模型;GBM. 推进式回归树模型;CTA. 分类树分析模型;ANN. 人工神经网络模型;SRE. 表面分布区分室模型;FDA. 柔性判别分析模型;RF. 随机森林模型。Figure 3. Response curves of predicted occurrence probability of A. planci and B. viridescens against environment variablesNote: GLM. Generalized Linear Model; GBM. Generalized Boosting Model; CTA. Classification Tree Analysis; ANN. Artificial Neural Network; SRE. Surface Range Envelope; FDA. Flexible Discriminant Analysis; RF. Random Forest.由图3可知,当温度低于6 ℃时,褐拟鳞鲀存在的概率很低,随着温度的增加,其存在的概率增大,当温度高于16 ℃时,其生境适宜性陡然增加,达0.8以上。而多数模型溶解氧浓度响应曲线的结果相似,其溶解氧浓度的适宜范围约为100~200 mol·m−3。褐拟鳞鲀的离岸距离响应曲线结果与长棘海星相似,不同模型间的差异较大。总体而言,离岸距离越小,生境适宜性越高。
2.3 长棘海星、褐拟鳞鲀适宜生境分布预测
2.3.1 长棘海星、褐拟鳞鲀当前在世界范围内的适宜生境分布预测
当前长棘海星潜在分布区主要集中在北印度洋和西太平洋海域,其极适宜生境广泛分布于澳大利亚北部海岸、印度尼西亚海岸以及中国南海一直延伸至东海海域,美洲中部海域及红海海域也有少量分布 (图4-a—4-b)。从褐拟鳞鲀的预测结果来看,其分布的主要区域与长棘海星大致重叠。其极适宜生境同样分布在澳大利亚北部海岸、印度尼西亚海岸以及中国南海海域 (图4-c—4-d)。与长棘海星不同的是,褐拟鳞鲀在非洲南部海岸、美洲中部海岸的潜在分布明显低于长棘海星,而在红海海域的分布明显高于长棘海星。两物种的预测结果总体表现为极高适宜生境与高适宜生境分布在热带、亚热带等低纬度地区,而低、中适宜生境向高纬度地区延伸扩散。
![]() 图 4 当前环境下长棘海星与褐拟鳞鲀的潜在分布适生区注:a、b. 长棘海星;c、d. 褐拟鳞鲀;WM. 概率加权平均组合模型;CA. 投票平均组合模型。Figure 4. Potential suitable habitat of A. planci and B. viridescens in current environmentNote: a, b. A. planci; c, d. B. viridescens; WM. Weighted Mean of Probabilities Ensemble Model; CA. Committee Averaging Ensemble Model.
图 4 当前环境下长棘海星与褐拟鳞鲀的潜在分布适生区注:a、b. 长棘海星;c、d. 褐拟鳞鲀;WM. 概率加权平均组合模型;CA. 投票平均组合模型。Figure 4. Potential suitable habitat of A. planci and B. viridescens in current environmentNote: a, b. A. planci; c, d. B. viridescens; WM. Weighted Mean of Probabilities Ensemble Model; CA. Committee Averaging Ensemble Model.长棘海星潜在适宜生境面积达3 027.80万km2,其中极适宜生境面积为460.90万km2 (约占全球海洋面积的1.2%),占适宜生境面积的15.22% (图4-a,表4),非适宜生境面积占绝大部分,达37 039.68万km2 (92.4%)。褐拟鳞鲀的潜在适宜生境比长棘海星更广,其面积达3 813.84万km2,其中极适宜生境面积为590.00万km2 (约占全球海洋面积的1.5%),占适宜生境面积的15.47% (图4-c,表4),非适宜生境面积达36 253.64万km2 (90.5%)。
表 4 长棘海星与褐拟鳞鲀的适宜生境占比Table 4. Proportions of suitable habitat for A. planci and B. viridescens物种
Species气候情景
Climate scenario低适宜生境占比
Proportion of
low suitability
habitat/%中适宜生境占比
Proportion of
moderately suitable
habitat/%高适宜生境占比
Proportion of
highly suitable
habitat/%极适宜生境占比
Proportion of
extremely suitable
habitat/%长棘海星
A. planci现在Current 40.23 24.33 20.22 15.22 RCP26 40.35 24.11 20.12 15.42 RCP60 40.18 24.06 20.09 15.67 RCP85 40.13 24.05 20.14 15.68 褐拟鳞鲀
B. viridescens现在Current 42.22 24.07 18.24 15.47 RCP26 41.82 24.10 18.31 15.77 RCP60 41.86 23.99 18.18 15.97 RCP85 41.86 23.90 18.24 16.00 注:非适宜生境不在计算范畴内。 Note: Unsuitable habitat is not included in the calculation. 2.3.2 未来不同气候情景下长棘海星和褐拟鳞鲀的适宜生境预测
两种不同组合模型 (WM、CA) 预测的长棘海星的适宜生境面积存在差异,但预测的总体趋势均为增大,不同气候情景下增幅不同。WM模型预测未来长棘海星在RCP26、RCP60和RCP85情景下总适宜生境面积分别增加78.87万、74.70万和102.41万km2,增幅分别为2.60%、2.47%和3.38%,其中极适宜生境增幅分别为3.97%、5.49%和6.49% (图5-a、5-c、5-e)。而CA模型预测RCP26、RCP60和RCP85情景下总适宜生境面积分别增加84.14万、75.83万和107.71万km2,增幅为3.50%、3.15%和4.48%,其中极适宜生境分别增加3.41%、3.82%和4.60% (图5-b、5-d、5-f)。不同气候情景下长棘海星未来的潜在分布面积均会扩大,且CA模型下的增幅略高于WM模型,但WM模型预测的极适宜生境面积增幅明显高于CA模型。
![]() 图 5 长棘海星未来不同气候情景下的潜在分布注:WM. 概率加权平均组合模型;CA. 投票平均组合模型;RCP26. 最低温室气体排放情景;RCP60. 较高温室气体排放情景;RCP85. 最高温室气体排放情景。Figure 5. Potential future distribution of A. planci under different climate scenariosNote: WM. Weighted Mean of Probabilities Ensemble Model; CA. Committee Averaging Ensemble Model; RCP26. Minimum greenhouse gas emission scenarios; RCP60. Higher greenhouse gas emission scenarios; RCP85. Maximum greenhouse gas emission scenarios.
图 5 长棘海星未来不同气候情景下的潜在分布注:WM. 概率加权平均组合模型;CA. 投票平均组合模型;RCP26. 最低温室气体排放情景;RCP60. 较高温室气体排放情景;RCP85. 最高温室气体排放情景。Figure 5. Potential future distribution of A. planci under different climate scenariosNote: WM. Weighted Mean of Probabilities Ensemble Model; CA. Committee Averaging Ensemble Model; RCP26. Minimum greenhouse gas emission scenarios; RCP60. Higher greenhouse gas emission scenarios; RCP85. Maximum greenhouse gas emission scenarios.褐拟鳞鲀全球潜在适宜生境在未来不同气候情景下也均表现为增加,WM模型预测RCP26、RCP60和RCP85情景下其总适宜生境面积分别增加90.75万、90.64万和112.09万km2,增幅分别为2.38%、2.37%和2.94%,其中极适宜生境分别增加4.38%、5.69%和6.49% (图6-a、6-c、6-e)。CA模型预测RCP26、RCP60和RCP85情景下总适宜生境面积分别增加111.13万、106.98万和145.99万km2,增幅为3.71%、3.58%和4.88%,其中极适宜生境分别增加5.02%、5.77%和6.78% (图6-b、6-d、6-f)。与长棘海星结果不同的是,CA模型预测褐拟鳞鲀的未来潜在分布总面积与极适宜生境面积增幅均高于WM模型,表现出模型对于不同物种的差异。除此之外,两物种的潜在分布区域均有向高纬度地区移动的趋势。
![]() 图 6 褐拟鳞鲀未来不同气候情景下的潜在分布注:WM. 概率加权平均组合模型;CA. 投票平均组合模型;RCP26. 最低温室气体排放情景;RCP60. 较高温室气体排放情景;RCP85. 最高温室气体排放情景。Figure 6. Potential future distribution of B. viridescens under different climate scenariosNote: WM. Weighted Mean of Probabilities Ensemble Model; CA. Committee Averaging Ensemble Model; RCP26. Minimum greenhouse gas emission scenarios; RCP60. Higher greenhouse gas emission scenarios; RCP85. Maximum greenhouse gas emission scenarios.
图 6 褐拟鳞鲀未来不同气候情景下的潜在分布注:WM. 概率加权平均组合模型;CA. 投票平均组合模型;RCP26. 最低温室气体排放情景;RCP60. 较高温室气体排放情景;RCP85. 最高温室气体排放情景。Figure 6. Potential future distribution of B. viridescens under different climate scenariosNote: WM. Weighted Mean of Probabilities Ensemble Model; CA. Committee Averaging Ensemble Model; RCP26. Minimum greenhouse gas emission scenarios; RCP60. Higher greenhouse gas emission scenarios; RCP85. Maximum greenhouse gas emission scenarios.3. 讨论
3.1 物种分布组合模型在适宜生境研究中的应用
多数研究表明,使用物种分布模型对海洋生物的潜在分布进行模拟时,各类单一模型的运行过程及算法存在不确定性和差异性,会降低预测的精度[26]。采用本研究构建的组合模型评估当前及未来3种不同气候情景下长棘海星与褐拟鳞鲀的潜在适生区动态变化,可有效降低单一模型造成的偏差。真实技巧统计值与受试者工作特征曲线是评价模型常用的指标[27]。研究结果显示,长棘海星与褐拟鳞鲀组合模型的TSS和ROC值高于大多数单一模型的结果,表明组合模型对两个物种潜在适生区的模拟结果准确。Weinert等[28]使用Biomod2程序包的6种单模型构建组合模型研究北海底栖生物的分布变化情况,结果也表明组合模型可降低模型的不确定性。本研究的模型预测结果与已知的长棘海星和褐拟鳞鲀的分布范围大致符合 (主要分布在北印度洋、西太平洋海域)。但本研究使用的建模方式仍存在一定的局限性。首先,物种扩散能力与种群动态息息相关,长棘海星与褐拟鳞鲀的扩散受到洋流及自身游泳能力的限制,但本研究并未考虑扩散的影响[29]。其次,人为因素对海洋生物的影响加剧[30],本研究只考虑了部分重要环境因素的影响。此外,受限于未来环境变量的获取,以及在实际情况下存在各种限制因素,本研究可能会高估长棘海星和褐拟鳞鲀的分布范围。生物间的相互作用也会对物种产生重要影响,褐拟鳞鲀作为长棘海星的重要捕食者,对长棘海星的种群动态也会产生干扰,并未纳入此次研究的范围。
3.2 影响长棘海星与褐拟鳞鲀适宜生境分布的主要因素
长棘海星与褐拟鳞鲀均生活在珊瑚礁生境,其种群动态与珊瑚礁生态系统密切相关。模型预测影响长棘海星空间分布的主要因素是温度和离岸距离,其中温度是最重要的因子。对褐拟鳞鲀分布有重要影响的环境变量为温度、溶解氧浓度与离岸距离,同样温度为最重要的因子。温度与海洋生物的新陈代谢有关,从而对生物的种群动态产生影响,模型结果显示,当温度高于15 ℃,长棘海星的生境适宜性增加。温度对长棘海星的产卵及幼虫成活率有重要影响,海水温度达27 ℃阈值时,长棘海星才开始产卵。其幼虫最佳发育温度在不同发育阶段有所差异,胚囊幼虫期为13~34 ℃,羽腕幼虫期为14.5~32 ℃,中晚期幼虫为26~30 ℃,当高于32 ℃时幼虫会死亡,低于25 ℃时则停止发育并退回早期发育阶段[31-32]。海水温度对中上层鱼类的水平和垂直移动有重要影响[33]。海温、叶绿素与上升流是影响鱼类空间分布的主要驱动因素,如鳀鱼 (Engraulis ringens) 等[34]。由此可见,温度可能是最主要和最普遍的鱼类分布驱动因子。离岸距离影响长棘海星分布,其原因可能与珊瑚礁形成有关,但未见相关的研究报道。长棘海星以珊瑚为食,在很大程度上珊瑚决定长棘海星的空间分布,而珊瑚对于生长环境的要求比较严格,主要分布在热带浅水海域,离海岸的距离较近。陆架边缘鱼类分布广泛、种类多,许多鱼类会在繁殖季节回到近岸海域产卵,因此离岸距离是影响鱼类栖息地的重要环境因子。一些鱼类的分布可能与特定生境的距离高度相关[35]。此外,溶解氧是海洋鱼类获得氧气的主要来源[36],对褐拟鳞鲀的栖息地分布起着重要作用。由于温室气体大量排放,未来全球气候变暖极有可能使当前适宜生存的环境因素超出物种的耐受阈值,诱发长棘海星和褐拟鳞鲀迁移到符合其生境偏好的新水域或在当前适宜生境做出适应性改变或急剧减少,从而引起两个物种的生境范围发生变化[37]。
3.3 不同气候情景下长棘海星与褐拟鳞鲀潜在适生区的变化
气候变暖导致全球海洋生物的丰度呈现向极地方向增加且向赤道方向减少的趋势[38]。本研究结果表明,当前环境下长棘海星的适宜栖息地主要分布于印度-太平洋海域、中国南海到澳大利亚海岸线、红海和阿拉伯海,与Sill和Dawson[18]的研究结果相似。本研究对长棘海星和褐拟鳞鲀当前和未来的栖息地分布做出了较为可靠的预测,可以为制定长棘海星暴发管理的策略提供一些参考。例如可针对长棘海星未来生境适宜性较高的海域,定期开展长棘海星的跟踪监测,对可能出现长棘海星暴发的海域进行预警[39]。当前环境下褐拟鳞鲀极适宜生境分布在40°N—40°S,其分布区域大部分覆盖了长棘海星的分布范围,可能是由于褐拟鳞鲀会捕食长棘海星,长棘海星存在的地方可以为它们提供部分食物来源,适合其生存[40]。未来不同气候情景下,长棘海星和褐拟鳞鲀的共性是两者的总体适生区范围均呈现出一定程度向高纬度地区扩大的趋势,模型预测温度是影响两个物种空间分布的最重要因素,未来全球气候变暖致使水温升高,促进了长棘海星与褐拟鳞鲀的生长发育。长棘海星具有逃离天敌存在区域的生物特性[41],因此可在其可能暴发的海域大量投放成本较低的长棘海星幼体和成体捕食者,从而减少长棘海星暴发的风险。在升温2 ℃的情况下,长棘海星幼虫的成活率可能会提高240%,在食物供应不变的情况下,发育速度会提高30%[42],从而导致其分布范围发生变化。本研究结果也符合未来条件下物种生存范围的变化趋势。研究显示气候变化改变了127种海洋生物的分布范围,其中大部分物种的潜在分布向高纬度海域移动[43]。在非洲南部海岸、美洲中部海岸,长棘海星的适宜生境明显比褐拟鳞鲀更广泛,这可能是因为在不适合褐拟鳞鲀栖息的地区,由于捕食者压力的潜在释放,会出现更为丰富的长棘海星种群[44]。到2050s,2种生物的潜在适宜生境在热带及亚热带海域的部分缩减,推测可能因气候变化使得海洋温度上升,海洋生物可能会向更适宜生存的极地迁徙[45]。在本研究记录数据以外的部分非洲和美洲沿岸海域也发现了褐拟鳞鲀的适宜栖息地,可能是由于气候变暖导致这些海域的理化性质改变,从而变得适合褐拟鳞鲀的生长。此外,控制海洋环境改变以及减少人为活动的干扰对减缓长棘海星暴发周期也具有重要作用[46],通过控制陆源污染以改善珊瑚礁区水质、设置禁渔或保护区以保护鱼类种群丰度和多样性,同时减少对长棘海星天敌的过度捕捞等都是维护珊瑚礁生态系统稳定和可持续发展的必要措施。
4. 结论
本研究基于组合物种分布模型,预测了不同气候情景下长棘海星与褐拟鳞鲀全球范围内的潜在分布格局,并分别探讨了2个物种分布的主要影响因子。结果表明,组合模型的结果优于单一模型,一定程度上能够提高预测的准确性。影响长棘海星空间分布的主要因素是温度和离岸距离,而影响褐拟鳞鲀空间分布的主要因素是温度、溶解氧浓度和离岸距离。长棘海星与褐拟鳞鲀在未来气候情景下的潜在分布均有扩大且向高纬度地区移动的趋势。本研究结果将为长棘海星生态管理的应用、维护生态系统多样性、制定未来气候适应管理决策提供科学依据。
-
表 1 培养基的碳源及添加质量浓度
Table 1 Carbon source and its concentration in medium
g · L-1 碳源
carbon source葡萄糖
glucose蔗糖
sucrose糖蜜
molasses葡萄糖+蔗糖
glucose+ sucrose葡萄糖+糖蜜
glucose+ molasses蔗糖+糖蜜
sucrose+ molasses添加质量浓度
concentration20 20 20 10+10 10+10 10+10  下载: 导出CSV
下载: 导出CSV
表 2 培养基的氮源及添加质量浓度
Table 2 Nitrogen source and its concentration in medium
g · L-1 氮源
nitrogen source蛋白胨
peptone酵母膏
yeast extract硫酸铵
(NH4)2SO4氯化铵
NH4Cl豆粕
soybean meal蛋白胨+酵母膏
peptone+ yeast extract蛋白胨+硫酸铵
peptone+ (NH4)2SO4酵母膏+硫酸铵
yeast extract +(NH4)2SO4添加质量浓度
concentration10 10 10 10 10 5+5 5+5 5+5
下载: 导出CSV
表 3 培养基的无机盐及添加质量浓度
Table 3 Inorganic salt and its concentration in medium
g · L-1 无机盐
inorganic salt磷酸二氢钾
KH2PO4硫酸镁
MgSO4氯化钙
CaCl2磷酸二氢钾+硫酸镁
KH2PO4+ MgSO4磷酸二氢钾+硫酸镁+氯化钙
KH2PO4+ MgSO4+ CaCl2添加质量浓度
concentration0.5 0.5 0.5 0.25+0.25 0.5+0.25+0.1
下载: 导出CSV
表 4 正交试验因素水平
Table 4 Factors and levels of orthogonal test
水平
level因素factor (A)硫酸镁/g·L-1
MgSO4(B)葡萄糖/g·L-1
glucose(C)酵母膏/g·L-1
yeast extract(D)蛋白胨/g·L-1
peptone1 0.25 15 5 5 2 0.50 20 10 10 3 0.75 25 15 15
下载: 导出CSV
表 5 碳源对RH1菌量的影响
Table 5 Effect of carbon sources onbiomass of RH1
108 cfu · mL-1 碳源carbon source 菌量biomass 葡萄糖glucose 6.79±0.79a 蔗糖sucrose 4.78±0.48bc 糖蜜molasses 5.43±0.56abc 葡萄糖+蔗糖glucose+sucrose 4.99±0.67bc 葡萄糖+糖蜜glucose+molasses 5.73±0.90ab 蔗糖+糖蜜sucrose+molasses 3.64±0.94c 注:表格中的数值是3次平行试验的平均值,数据后不同上标字母表示5%显著水平。后表同此
Note:Values are expressed as average of 3 replicates; values with different superscript letters within the same column are significantly different(P<0.05). The same case in the following tables.
下载: 导出CSV
表 6 氮源对RH1菌量的影响
Table 6 Effect of nitrogen sources on biomass of RH1
108 cfu · mL-1 氮源nitrogen source 菌量biomass 蛋白胨peptone 4.13±0.64a 酵母膏yeast extract 6.89±0.68b 硫酸铵(NH4)2SO4 1.34±0.63c 氯化铵NH4Cl 1.18±0.76c 豆粕soybean meal 1.35±0.49c. 蛋白胨+酵母膏peptone+yeast extract 7.13±1.18b 蛋白胨+硫酸铵peptone+(NH4)2SO4 3.87±0.78a 酵母膏+硫酸铵yeast extract+(NH4)2SO4 4.04±0.92a
下载: 导出CSV
表 7 无机盐对RH1菌量的影响
Table 7 Effect of inorganic salts on biomass of RH1
108 cfu · mL-1 无机盐inorganic salt 菌量biomass 磷酸二氢钾KH2PO4 4.43±0.33a 硫酸镁MgSO4 6.14±1.17ab 氯化钙CaCl2 4.22±0.76a 磷酸二氢钾+硫酸镁
KH2PO4+MgSO46.90±0.47b 磷酸二氢钾+硫酸镁+氯化钙
KH2PO4+MgSO4+CaCl25.27±1.27ab
下载: 导出CSV
表 8 正交试验结果分析
Table 8 Results of analysis of orthogonal test
108 cfu · mL-1 编号
No.A B C D 菌量
biomass1 1 1 1 1 6.04 2 1 2 2 2 8.44 3 1 3 3 3 4.60 4 2 1 2 3 4.08 5 2 2 3 1 6.76 6 2 3 1 2 5.92 7 3 1 3 2 7.32 8 3 2 1 3 3.64 9 3 3 2 1 5.56 k1 6.36 5.82 5.20 6.12 k2 5.58 6.28 6.02 7.22 k3 5.50 5.36 6.22 4.10 R 0.86 0.92 1.02 3.12
下载: 导出CSV
-
[1] BON J A, LEATHERS T D, JAYASWAL R K. Isolation of astaxanthin-overproducing mutants of Phaffia rhodozymal[J]. Biotechnol Lett, 1997, 19(2): 109-112. doi: 10.1023/A:1018391726206
[2] 刘秀莲, 王宇光. 海洋红酵母的研究进展[J]. 生物技术通讯, 2008, 19(2): 293-295. doi: 10.3969/j.issn.1009-0002.2008.02.039 LIU Xiulian, WANG Yuguang. Advance in the research of the Rhodotorula benthica[J]. Lett Biotechnol, 2008, 19(2): 293-295. (in Chinese) doi: 10.3969/j.issn.1009-0002.2008.02.039
[3] 李臣, 阮榕生, 林向阳, 等. 红酵母菌的性质及其应用研究[J]. 农产品加工学刊, 2006(5): 20-25. https://xueshu.baidu.com/usercenter/paper/show?paperid=fa4a63f19e6e7d35176171adfd4a8ee8&site=xueshu_se&hitarticle=1 LI CHEN, RUAN Rongsheng, LIN Xiangyang, et. al. Advances in research of properties and application of Rhodotorula[J]. Acad Periodic Farm Prod Process, 2006(5): 20-25. (in Chinese) https://xueshu.baidu.com/usercenter/paper/show?paperid=fa4a63f19e6e7d35176171adfd4a8ee8&site=xueshu_se&hitarticle=1
[4] 赵述淼, 韩继宏. 谈酵母在水产养殖上的应用[J]. 中国水产, 2003(7): 68-69. doi: 10.3969/j.issn.1002-6681.2003.07.052 ZHAO Shumiao, HAN Jihong. The application of marine red yeast in aquaculture[J]. China Fish, 2003(7): 68-69. (in Chinese) doi: 10.3969/j.issn.1002-6681.2003.07.052
[5] 王伟霞, 李福后, 浦寅芳. 海洋红酵母产类胡萝卜素的研究[J]. 食品研究与开发, 2007, 28(10): 55-58. doi: 10.3969/j.issn.1005-6521.2007.10.018 WANG Weixia, LI Fuhou, PU Yinfang. Investigation on Carotenoids from marine Rhodotorula sp. [J]. Food Res Dev, 2007, 28(10): 55-58. (in Chinese) doi: 10.3969/j.issn.1005-6521.2007.10.018
[6] 王宇光, 雷禄旺, 刘秀莲, 等. 海南海域海洋红酵母的分离鉴定和应用性能评价[J]. 中国农学通报, 2009, 25(22): 324-328. https://xueshu.baidu.com/usercenter/paper/show?paperid=c06033e9e502a2e636e932738625631e&site=xueshu_se&hitarticle=1 WANG Yuguang, LEI Luwang, LIU Xiulian, et. al. Isolation and identification of Rhodotorula in Hainan sea area and evaluation of its application performance[J]. Chin Agric Sci Bull, 2009, 25(22): 324-328. (in Chinese) https://xueshu.baidu.com/usercenter/paper/show?paperid=c06033e9e502a2e636e932738625631e&site=xueshu_se&hitarticle=1
[7] 童应凯, 韦东胜, 刘婷, 等. 海洋红酵母菌培养条件的研究[J]. 食品研究与开发, 2006, 27(8): 75-79. doi: 10.3969/j.issn.1005-6521.2006.08.023 TONG Yingkai, WEI Dongsheng, LIU Ting, et al. Study on fermentation conditions in Rhodotorual benthica[J]. Food Res Dev, 2006, 27(8): 75-79. (in Chinese) doi: 10.3969/j.issn.1005-6521.2006.08.023
[8] 王普, 裘娟萍, 郑裕国, 等. 高产虾青素的红发夫酵母菌种的选育[J]. 微生物学通报, 2002, 29(1): 15-19. doi: 10.3969/j.issn.0253-2654.2002.01.004 WANG Pu, QIU Juanping, ZHENG Yuguo, et al. Breeding of Phaffia rhodozyma for astaxanthin over-production[J]. Microbiology, 2002, 29(1): 15-19. (in Chinese) doi: 10.3969/j.issn.0253-2654.2002.01.004
[9] 吴江, 刘子贻. 法夫红酵母高产虾青素菌株的研究概况[J]. 生物技术通报, 1999(4): 23-25. doi: 10.3969/j.issn.1002-5464.1999.04.006 WU Jiang, LIU Ziyi. The survey of astaxanthin high-producing stains of Phattia rhodozyma[J]. Biotechnol Info, 1999(4): 23-25. (in Chinese) doi: 10.3969/j.issn.1002-5464.1999.04.006
[10] MISAWA N, SHIMADA H. Metabolic engineering for the production of carotenoids in non-carotenogenic bacteria and yeasts[J]. Biotechnology, 1998, 59(3): 169-181. https://xueshu.baidu.com/usercenter/paper/show?paperid=bba15988eb99de03bd640859916aee0b&site=xueshu_se
[11] NAGAHAMA T, HAMAMOTO M, NAKASE T, et al. Rhodotorula benthica sp. nov. and Rhodotorula calyptogenae sp. nov., novel yeast species from animals collected from the deep-sea floor, and Rhodotorula lysiniphia sp. nov., which is related phylogentically[J]. Int J Syst Evol Microbiol, 2003, 53(3): 897-903. doi: 10.1099/ijs.0.02395-0
[12] 陈昌福, 姚鹃, 肖雪生. 海洋1号红酵母饲养日本对虾幼虾的效果[J]. 淡水渔业, 2004, 34(6): 55-57. doi: 10.3969/j.issn.1000-6907.2004.06.020 CHEN Changfu, YAO Juan, XIAO Xuesheng. The effect of the No. 1 marine red yeast on feeding juvenile Japanese prawn[J]. Freshwater Fish, 2004, 34(6): 55-57. (in Chinese) doi: 10.3969/j.issn.1000-6907.2004.06.020
[13] 李爱娟, 何伯峰, 李光. 海洋红酵母在水产养殖中的应用[J]. 齐鲁渔业, 2007, 24(12): 62. https://xueshu.baidu.com/usercenter/paper/show?paperid=15bebaa662142a362b35988dfb78bbc7&site=xueshu_se LI Aijuan, HE Bofeng, LI Guang. The application of marine red yeast in aquaculture[J]. Shandong Fish, 2007, 24(12): 62. (in Chinese) https://xueshu.baidu.com/usercenter/paper/show?paperid=15bebaa662142a362b35988dfb78bbc7&site=xueshu_se
[14] 杨秋明, 蔡慧农, 宁思扬, 等. 海洋红酵母产虾青素培养基优化的初步研究[J]. 微生物学杂志, 2007, 27(1): 72-75. doi: 10.3969/j.issn.1005-7021.2007.01.017 YANG Qiuming, CAI Huinong, NING Siyang, et. al. Medium optimization of astaxanthin by oceanic red yeast[J]. J Microbiol, 2007, 27(1): 72-75. (in Chinese) doi: 10.3969/j.issn.1005-7021.2007.01.017
[15] 张超, 刘军. SCP生产菌海洋红酵母摇瓶培养研究[J]. 四川轻化工学院学报, 2000, 13(3): 71-74. https://xueshu.baidu.com/usercenter/paper/show?paperid=d8cd7a827caac2562c59b3748b06d684&site=xueshu_se ZHANG Chao, LIU Jun. Study on the culture in flask scale of SCP-producing stain oceanic red yeast[J]. J Sichuan Inst Light Ind Chem Technol, 2000, 13(3): 71-74. (in Chinese) https://xueshu.baidu.com/usercenter/paper/show?paperid=d8cd7a827caac2562c59b3748b06d684&site=xueshu_se
[16] 张志军, 江晓路, 张婷婷, 等. 海洋红酵母Rhodotorula sp. 06产胡萝卜素的研究[J]. 食品工业科技, 2007, 28(11): 123-125, 131. doi: 10.3969/j.issn.1002-0306.2007.11.037 ZHANG Zhijun, JIANG Xiaolu, ZHANG Tingting, et al. The study on carotenoids production by marine Rhodotorula sp. 06[J]. Sci Technol Food Ind, 2007, 28(11): 123-125, 131. (in Chinese) doi: 10.3969/j.issn.1002-0306.2007.11.037
[17] 钱铭镛. 发酵工程最优化控制[M]. 南京: 江苏科学出版社, 1998: 306-309. https://xueshu.baidu.com/usercenter/paper/show?paperid=9cf151481a62d6ee949df004d4628168&site=xueshu_se&hitarticle=1 QIAN Mingyong. The optimization of fermentation engineering[M]. Nanjing: Jiangsu Science Press, 1998: 306-309. (in Chinese) https://xueshu.baidu.com/usercenter/paper/show?paperid=9cf151481a62d6ee949df004d4628168&site=xueshu_se&hitarticle=1
[18] 李宁, 柯晓静, 林场, 等. 产β胡萝卜素红酵母的筛选及其发酵废糖蜜的研究[J]. 农产品加工学刊, 2009(6): 43-45, 48. https://xueshu.baidu.com/usercenter/paper/show?paperid=5775c8dd5da5d893a6226c6651816dd5&site=xueshu_se&hitarticle=1 LI Ning, KE Xiaojing, LIN Chang, et al. Screening of Rhodotorula producing β-carotene and fermentation with wasted molasses[J]. Acad Periodic Farm Prod Process, 2009(6): 43-45, 48. (in Chinese) https://xueshu.baidu.com/usercenter/paper/show?paperid=5775c8dd5da5d893a6226c6651816dd5&site=xueshu_se&hitarticle=1
[19] 张超. 补料分批培养海洋红酵母的研究[J]. 四川轻化工学院学报, 2000, 13(2): 26-30. https://xueshu.baidu.com/usercenter/paper/show?paperid=163x0gt0fj4d08t04c2v0ju0tk332020&site=xueshu_se&hitarticle=1 ZHANG Chao. Study on fed-batch culture of oceanic red yeast[J]. J Sichuan Inst Light Ind Chem Technol, 2000, 13(2): 26-30. (in Chinese) https://xueshu.baidu.com/usercenter/paper/show?paperid=163x0gt0fj4d08t04c2v0ju0tk332020&site=xueshu_se&hitarticle=1
[20] BHOSALE P, GADRE R V. Production of β-carotene by a Rhodotorula guutinis mutant in sea water medium[J]. Bioresour Technol, 2001, 76(1): 53-55. doi: 10.1016/S0960-8524(00)00075-4






计量
- 文章访问数: 4212
- HTML全文浏览量: 194
- PDF下载量: 2493
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
