
Auto-feeding system for deep-water net cage based on PLC
-
摘要:
为提高深水网箱养殖投饵的自动化和工业化水平,设计并开发出适合深水网箱养殖投饵管理的自动投饵系统。该系统采用PLC为控制核心,继电接触器控制各个设备的启停,实现精确投饵。文章介绍了系统的工作原理和设计框架,阐述了系统控制电路、系统工作流程与PLC硬件结构,设计了系统关键设备控制方案并编写了相关控制程序。检测结果表明,在投饵输送距离为300 m时,投饵量为1 200 kg·h-1,系统风量为5.56 m3·min-1,系统风压为49 kPa;吨料能耗最大为8.4 kW·h·t-1(平均为5.0 kW·h·t-1),投饵破碎率小于0.7%,喷投距离达11.3 m;系统可实现动态定时、定点和定量投喂饲料,完全满足深水网箱集群养殖的要求。
Abstract:To promote feeding automation and industrialization, an auto-feeding system is designed and developed for management of culture and feeding by deep-water net cage. PLC is used as control core, and the start-stop of each equipment is controlled by relay contactors to realize precise feeding. We introduce the working principle and design framework of the system, describe the control circuit, working process of the system and PLC hardware structure, design the control methods of key equipments and write related control program. The results show that at feeding transportation distance of 300 m, feeding quantity is 1 200 kg·h-1, and air volume of system is 5.56 m3·min-1 and air pressure of system is 49 kPa. Maximum energy consumption reaches 8.4 kW·h·t-1 (5.0 kW·h·t-1 averagely), and breaking ratio of feed is less than 0.7% and spraying distance is 11.3 m. With this system, feeding can be carried out by dynamic timing at designated points in fixed quantity, which meets the requirement of cluster culture by deep-water net cage.
-
Keywords:
- PLC /
- automatic feeding /
- deep-water net cage
-
杜氏枪乌贼(Uroteuthis duvaucelii,也常称为Loligo duvauceli)隶属于头足纲、蛸亚纲、枪形目、闭眼亚目、枪乌贼科,广泛分布于太平洋和印度洋沿岸,在红海、阿拉伯海和印度沿岸资源量较大[1-3],同时也广泛分布于泰国沿岸及我国东海和南海海域[4-6]。随着我国渔业资源严重小型化,该物种也逐渐成为我国沿海重要的经济性头足类之一[7]。头足类的角质颚是重要的摄食器官,包裹于口球中,分为上、下颚,作为一种具有稳定形态和丰富生态信息的硬组织,得到了许多研究者的关注[8],常常被用于年龄鉴定、种群划分、食性评估和资源丰度等研究中[9-11]。据报道,处于不同性成熟度的头足类,由于摄食种类的差异,其角质颚的形态和生长规律也有所差异,因此角质颚的形态可以用于表征头足类性成熟的特征[12]。已有针对多种大洋性头足类如柔鱼(Ommastrephes bartramii)[13]、茎柔鱼(Dosidicus gigas)[14]、阿根廷滑柔鱼(Illex argentinus)[15]等以及近海头足类如剑尖枪乌贼(U.edulis)[16]、太平洋褶柔鱼(Todarodes pacificus)[17]、火枪乌贼(Loliolus beka)[18]等角质颚的形态和生长特性研究,尚未见对杜氏枪乌贼角质颚的形态和生长的研究。因此,本研究根据我国拖网船2015年9—10月在南海北部海域进行渔业生产期间采集的杜氏枪乌贼样本,测定其角质颚的形态生长规律,利用主成分分析得到形态参数,通过比较不同胴长组和不同性成熟度角质颚的形态参数,分析性成熟对杜氏枪乌贼角质颚形态和生长的影响,为其种群区分和资源可持续合理利用提供技术支撑。
1. 材料与方法
1.1 采集时间和范围
样本主要从南海北部大陆架海域拖网船中获取,采集时间为2015年9—10月。最终共捕获个体512尾(雌性281尾,雄性231尾)。捕获的样品在船上直接冷冻,运回实验室进行后续分析。
1.2 基础生物学测量
样本运回实验室解冻后,对杜氏枪乌贼进行生物学测定,包括胴长(mantel length,LM)、体质量(body mass,M)、性别、性腺成熟度等。测量胴长用卷尺进行,测定精确至1 mm;测定体质量用电子天平进行,精确至1 g。性成熟度的划分参照Lipiński等[19]根据性腺的特征变化划分Ⅰ、Ⅱ、Ⅲ、Ⅳ、Ⅴ五期,其中Ⅰ和Ⅱ为未成熟,Ⅲ期以上为成熟个体。用镊子从口球中取出角质颚,并存放于盛有75%乙醇溶液的50 mL离心管中。最后得到完整角质颚样本共477对,其中雌性258对(未成熟94对,性成熟164对),雄性219对(均为未成熟个体)。
1.3 角质颚外形测量
将角质颚从乙醇中取出,用清水再次清洗并晾干。利用数显游标卡尺进行形态测量,首先沿水平和垂直2个方向进行校准,然后对角质颚的上头盖长(upper hood length,LUH)、上脊突长(upper crest length,LUC)、上喙长(upper rostrum length,LUR)、上侧壁长(upper lateral wall length,LULW)、上翼长(upper wing length,LUW)、下头盖长(lower hood length,LLH)、下脊突长(lower crest length,LLC)、下喙长(lower rostrum length,LLR)、下侧壁长(lower lateral wall length,LLLW)、下翼长(lower wing length,LLW) 10项形态参数进行测量(图1),测量结果精确至0.01 mm。
![]() 图 1 杜氏枪乌贼角质颚外部形态测量示意图 (左图为上颚,右图为下颚)AB. 头盖长;AC. 脊突长;AE. 喙长;ED. 翼长;AF. 侧壁长Figure 1. Scheme of morphometric measurements of U.duvaucelii beak (left: upper beak; right: lower beak)AB. hood length; AC. crest length; AE. rostrum length; ED. wing length; AF. lateral wall length
图 1 杜氏枪乌贼角质颚外部形态测量示意图 (左图为上颚,右图为下颚)AB. 头盖长;AC. 脊突长;AE. 喙长;ED. 翼长;AF. 侧壁长Figure 1. Scheme of morphometric measurements of U.duvaucelii beak (left: upper beak; right: lower beak)AB. hood length; AC. crest length; AE. rostrum length; ED. wing length; AF. lateral wall length1.4 数据处理方法
1)采用频度分析法分雌雄分析渔获物胴长及体质量组成,组间距分别为20 mm和20 g。
2)由于不同性别和性成熟度下的个体差异较大,因此将不同性别和性成熟度个体分开分析。首先检验数据的正态性,不满足正态分布的则进行平方根转化的标准化处理[20];然后利用方差分析(ANOVA)对雌雄个体胴长、体质量以及角质颚形态进行差异性检验,以检测是否存在性别差异。同时建立胴长与体质量的生长方程,其公式[21]为:
$$ \quad\quad\quad\quad\quad\quad\quad\quad\quad\;M= a {L _{\rm {M}}}^{b} $$ (1) 式中M为体质量(g);LM为胴长(mm);a、b为估算参数。
3)参考韩青鹏等[22],利用不同的生长模型(线性、幂函数、指数函数、对数函数)拟合角质颚形态参数与胴长之间的关系。
利用赤池信息量准则(Akaike information criterion,AIC),选取值最小的为最适生长模型,计算公式为:
$$ \quad\quad\quad\quad\quad\quad\; {\rm AIC} = {\rm{2}} \times k + n \times \ln (\frac{{\rm RSS}}{n}) $$ (2) 式中k为方程中参数常数的数量,n为样本量,RSS为剩余平方和。
4)为检验不同胴长组和不同性成熟度对雌性个体角质颚形态差异的影响,运用方差分析对不同胴长组和不同性成熟雌性个体的角质颚各项参数值进行差异性检验。对于存在极显著性差异的(P<0.01),采用Tukey法进一步进行组间多重比较[23],以便分析不同胴长和性成熟度的角质颚形态的具体差异所在。
所有统计分析采用SPSS 17.0软件进行。
2. 结果
2.1 胴长、体质量组成及其关系
统计表明,雌性未成熟个体胴长、体质量范围分别为36~110 mm、4~51 g,对应的优势胴长和体质量为60~100 mm、0~20 g,占总数的92.55%、72.34%;雌性成熟个体胴长、体质量范围分别为36~134 mm、15~104 g,对应的优势胴长和体质量为60~100 mm、20~40 g,占总数的89.02%、88.41%;雄性未成熟个体胴长、体质量范围分别为36~164 mm、3~84 g,对应的优势胴长、体质量为60~100 mm、20~40 g,占总数的85.32%、88.58%。以上3组个体的胴长(ANOVA,F=10.654,P<0.01)和体质量(ANOVA,F=4.79,P<0.01)均存在显著差异(图2)。通过多重比较发现,仅不同性别未成熟个体间胴长存在显著差异(P<0.01);仅不同性成熟度雌性个体间体质量存在显著差异(P<0.01)。
![]() 图 2 不同性别杜氏枪乌贼胴长与体质量大小组成分布图Figure 2. Distribution of mantle length and body mass of U.duvaucelii for different sexes
图 2 不同性别杜氏枪乌贼胴长与体质量大小组成分布图Figure 2. Distribution of mantle length and body mass of U.duvaucelii for different sexes根据上述方差分析(ANOVA)的结果,本研究将不同性别和性成熟度个体分开讨论。经过拟合(图3),不同性别和性成熟度胴长与体质量关系为:
![]() 图 3 不同性别杜氏枪乌贼胴长与体质量的关系虚线为成熟个体与胴长关系,实线为未成熟个体与胴长关系,后图同此Figure 3. Relationship between mantle length and body mass of U.duvaucelii for different sexesDotted line represent mature individuals, and solid line represent immature individuals. The same case in the following figures.
图 3 不同性别杜氏枪乌贼胴长与体质量的关系虚线为成熟个体与胴长关系,实线为未成熟个体与胴长关系,后图同此Figure 3. Relationship between mantle length and body mass of U.duvaucelii for different sexesDotted line represent mature individuals, and solid line represent immature individuals. The same case in the following figures.雌性未成熟:M=0.001 2LM2.223(n=94,R2=0.761,P<0.01)
雌性成熟:M=0.013 5LM1.757 2(n=164,R2=0.902,P<0.01)
雄性未成熟:M=0.001 1LM2.249 1(n=219,R2=0.893,P<0.01)
2.2 不同性别和性成熟度个体角质颚形态差异
不同性别杜氏枪乌贼角质颚形态值见表1。其中雌性成熟个体的角质颚形态的各项参数均大于其他2组。而不同性别未成熟个体的角质颚形态较为相似,雄性略大于雌性。
表 1 不同性别和性成熟度杜氏枪乌贼角质颚形态值Table 1. Beak morphometrics of U.duvaucelii for different sexes and maturity stages参数
parameter雌性未成熟
immature female雌性成熟
mature female雄性未成熟
mature female极值 extremum 均值±标准差 $ {\overline X \pm {\rm SD}}$ 极值 extremum 均值±标准差 $ {\overline X \pm {\rm SD}}$ 极值 extremum 均值±标准差 $ {\overline X \pm {\rm SD}}$ LUH 2.49~8.69 5.22±1.47 3.62~9.58 6.07±1.11 2.67~10.87 5.53±1.44 LUC 4.44~11.91 7.33±1.90 4.23~14.09 8.38±1.54 4.25~13.03 7.73±1.83 LUR 0.98~3.04 1.68±0.44 1.17~2.90 1.96±0.30 0.45~2.86 1.76±0.43 LULW 3.47~9.05 5.67±1.35 4.09~10.92 6.48±1.24 3.07~9.89 5.95±1.35 LUW 1.17~3.67 2.24±0.56 1.27~5.94 2.45±0.69 1.08~5.03 1.34±0.65 LLH 1.32~4.00 2.09±0.49 1.47~3.89 2.32±0.44 1.21~3.87 2.13±0.50 LLC 2.31~6.38 4.00±1.07 1.38~7.76 4.73±0.90 1.30~7.17 4.18±1.13 LLR 0.59~2.26 1.52±0.34 0.79~2.95 1.55±0.45 0.45~4.93 1.56±0.47 LLLW 1.51~8.00 4.82±1.37 3.30~9.71 5.79±1.05 1.69~8.68 5.06±1.33 LLW 1.13~5.58 3.37±0.86 2.28~7.56 3.64±0.95 1.18~6.26 3.64±0.89 通过方差分析可以发现,除了下喙长以外,不同性别和性成熟度个体间的各项角质颚参数值均存在显著差异(P<0.05)。利用多重比较分析(Tukey HSD)进一步分析发现,除了下喙长和下翼长外,不同性成熟度雌性个体间的各项角质颚参数值均存在显著差异(P<0.05),雌性成熟个体与雄性未成熟个体间也有类似的关系。而在不同性别未成熟个体间,所有的角质颚形态参数值均不存在显著差异(P>0.05,表2)。
表 2 不同性别和性成熟度杜氏枪乌贼角质颚形态值方差分析Table 2. Beak morphometrics variation of U.duvaucelii in different sexes and maturity stages参数
parameter整体比较
whole comparisonTukey-HSD 雌未-雌成熟 FI-FM 雌未-雄未 FI-MI 雌成熟-雄未 FM-MI F P SE P SE P SE P LUH 13.68 <0.01 0.17 <0.01 0.16 >0.05 0.14 <0.01 LUC 11.97 <0.01 0.23 <0.01 0.22 >0.05 0.19 <0.01 LUR 18.77 <0.01 0.05 <0.01 0.04 >0.05 0.04 <0.01 LULW 13.45 <0.01 0.17 <0.01 0.16 >0.05 0.13 <0.01 LUW 3.32 <0.05 0.08 <0.05 0.08 >0.05 0.06 >0.05 LLH 9.17 <0.01 0.06 <0.01 0.06 >0.05 0.05 <0.01 LLC 19.03 <0.01 0.13 <0.01 0.13 >0.05 0.11 <0.01 LLR 0.254 >0.05 0.06 >0.05 0.05 >0.05 0.04 >0.05 LLLW 23.22 <0.01 0.16 <0.01 0.15 >0.05 0.13 <0.01 LLW 3.38 <0.05 0.12 >0.05 0.11 <0.05 0.09 >0.05 注:SE. 标准误;雌未. 雌性未成熟个体;雄未. 雄性未成熟个体;雌成熟. 雌性成熟个体;P<0.01. 差异极显著;P<0.05. 差异显著;P>0.05. 差异不显著 Note: SE. standard error; FI. immature female; MI. immature male; FM. mature female; P<0.01. very significant difference; P<0.05. significant different; P>0.05. insignificant difference 2.3 角质颚形态参数与胴长的关系
结合上述差异分析,本研究选取了头盖长(LUH和LLH),脊突长(LUC和LLC)和侧壁长(LULW和LLLW)分别与胴长进行拟合分析。通过尝试不同模型的拟合后发现,根据AIC准则,所有上颚参数值与胴长的关系符合线性模型(图4)。
![]() 图 4 不同性别和性成熟度杜氏枪乌贼上颚形态值与胴长的关系Figure 4. Relationship between upper beak morphometircs and mantle length of U.duvaucelii for different sexes
图 4 不同性别和性成熟度杜氏枪乌贼上颚形态值与胴长的关系Figure 4. Relationship between upper beak morphometircs and mantle length of U.duvaucelii for different sexes雌性未成熟:
LUH=0.066 6LM+0.771 6 (n=94,R2=0.763,P<0.01)
LUC=0.086 0LM+1.587 2 (n=94,R2=0.762,P<0.01)
LULW=0.061 0LM+1.590 0 (n=94,R2=0.761,P<0.01)
雌性成熟:
LUH=0.043 9LM+3.188 2 (n=164,R2=0.697,P<0.01)
LUC=0.059 9LM+4.440 8 (n=164,R2=0.669,P<0.01)
LULW=0.045 9LM+3.468 2 (n=164,R2=0.610,P<0.01)
雄性未成熟:
LUH=0.050 2LM+1.733 7 (n=219,R2=0.893,P<0.01)
LUC=0.065 0LM+2.820 1 (n=219,R2=0.774,P<0.01)
LULW=0.042 4LM+2.741 2 (n=219,R2=0.608,P<0.01)
与上颚和胴长的关系相同,根据AIC准则,所有下颚参数值与胴长的关系符合线性模型(图5)。
![]() 图 5 不同性别和性成熟度杜氏枪乌贼下颚形态值与胴长的关系Figure 5. Relationship between lower beak morphometircs and mantle length of U.duvaucelii for different sexes
图 5 不同性别和性成熟度杜氏枪乌贼下颚形态值与胴长的关系Figure 5. Relationship between lower beak morphometircs and mantle length of U.duvaucelii for different sexes雌性未成熟:
LLH=0.018 9LM+0.833 0 (n=94,R2=0.545,P<0.01)
LLC=0.046 4LM+0.899 6 (n=94,R2=0.694,P<0.01)
LLLW=0.055 0LM+1.143 2 (n=94,R2=0.599,P<0.01)
雌性成熟:
LLH=0.012 6LM+1.493 5 (n=164,R2=0.356,P<0.01)
LLC=0.032 4LM+2.600 8 (n=164,R2=0.573,P<0.01)
LLLW=0.040 4LM+3.135 9 (n=164,R2=0.653,P<0.01)
雄性未成熟:
LLH=0.016 1LM+0.918 2 (n=219,R2=0.639,P<0.01)
LLC=0.038 2LM+1.286 6 (n=219,R2=0.696,P<0.01)
LLLW=0.044 2LM+1.722 3 (n=219,R2=0.681,P<0.01)
2.4 不同胴长组和不同性成熟度对雌性角质颚形态的影响
由于雌性个体差异较大,因此本研究根据胴长分布,以20 mm为间距,将胴长分为8个组。同时也将性腺成熟度细化分为Ⅰ、Ⅱ、Ⅲ和Ⅳ 4个等级,利用双因素方差分析(Two-way ANOVA)来分析雌性个体大小和性成熟度以及其交互作用对角质颚形态的影响。在剔除上文方差分析结果中差异不显著的形态值LLR后,其余形态参数无论上下颚,在不同的胴长组间均存在显著差异(P<0.01)。在结合性成熟度分析后,结果显示除了下翼长,其余形态参数均受到胴长和性成熟交互作用的影响(P<0.01,表3)。
表 3 不同胴长组和性成熟度对雌性杜氏枪乌贼角质颚形态的影响Table 3. Influence of mantle length groups and maturity stages on beak morphometric parameters of female U.duvaucelii参数
parameter主体间效应 main effect 胴长组 mantle length group 性成熟度 maturity stage group 交互作用 interaction F P F P F P LUH 108.05 <0.01 3.57 <0.01 4.57 <0.01 LUC 90.57 <0.01 3.18 <0.01 3.75 <0.01 LUR 9.72 <0.01 4.75 <0.01 2.50 <0.01 LULW 76.10 <0.01 5.10 <0.01 3.13 <0.01 LUW 36.72 <0.01 1.25 <0.05 1.81 <0.05 LLH 35.80 <0.01 0.31 <0.05 4.33 <0.01 LLC 70.64 <0.01 3.59 <0.01 2.76 <0.01 LLLW 72.51 <0.01 5.94 <0.01 3.34 <0.01 LLW 39.06 <0.01 1.04 >0.05 1.89 >0.05 注:P<0.01. 差异极显著;P<0.05. 差异显著;P>0.05. 差异不显著 Note: P<0.01. very significant difference; P<0.05. significant difference; P>0.05. insignificant difference 根据多重比较进一步发现,在不同胴长组中,除了40~60 mm与60~80 mm的上喙长外,其余组在上颚形态值间均存在显著差异(P<0.01);除了40~60 mm与60~80 mm的下头盖长外,其余组在下颚形态值间均存在显著差异(P<0.01)。在不同的性成熟度中,所有上颚形态值仅在Ⅲ期和Ⅳ期间不存在差异(P>0.05),其余性成熟度间均存在显著差异(P<0.01),其中上喙长在Ⅱ期与Ⅲ期间也不存在显著差异(P>0.05);下颚与上颚有所不同,所有下颚形态值仅在Ⅰ期和Ⅱ期间存在显著差异(P<0.01),其余性成熟度间均不存在显著差异(P>0.05)。
3. 讨论
3.1 个体大小特征
在我国东海和南海的相关渔业资源调查中,杜氏枪乌贼在渔获物的头足类中所占比例较大,是重要的优势种(IRI>500)[5-6],因此了解其基础生物学对开发该资源有着重要的意义。本研究中所采集的样本雌雄比例较为接近,总体雄性个体大于雌性个体,雌性在个体较小时已经达到性成熟。目前该种类在国内公开发表的基础生物学报道较少。对比国外同种类的研究发现,印度孟买沿岸[24]和红海北部海域[2]所捕获的杜氏枪乌贼,均有超过200 mm的个体发现,平均胴长都在100 mm以上,平均体质量也在40 g以上。本研究中个体相对较小,这与其所处的不同环境中饵料和营养物有着较大的关系[25]。从胴长体质量关系来看,其中未成熟个体的系数b为2.22~2.25,而雌性成熟个体仅为1.75,这在头足类的生长中属于较慢类型。对比其他海域的生长系数,在埃及沿岸[2]和泰国沿岸[4]的雄性个体,以及印度西海岸的雌雄个体[26]均出现系数低于2 (1.61~1.95)的情况。性成熟与未成熟个体生长的差异,也直接说明了枪乌贼生长的策略,即幼体期间保持个体较快生长,达到性成熟后,能量主要供给于性腺发育。总体而言,由于受到近岸海洋环境的影响,杜氏枪乌贼的胴长体质量关系处于一种较为平缓的异速生长模式[2]。
3.2 不同性别和性成熟度角质颚形态特征
头足类的角质颚是主要的摄食器官,其形态差异可以反映出个体的生长情况和摄食习性。本研究中对角质颚各项形态参数方差分析的结果发现,不同性成熟阶段雌性角质颚形态存在显著差异。头足类在生长过程中,由于个体生长发育的需要,食性会发生一定的变化,往往是从较小的甲壳类变为与其体型相似的鱼类,甚至同类[27]。因此在个体从未成熟向性成熟转变的过程中,角质颚的形态也会发生较大的变化,这在柔鱼、阿根廷滑柔鱼等种类中也有所发现。雌雄二态性(sexual dimorphism)在头足类中较为常见,其中角质颚的形态也有类似的特征[28-29]。本研究中对比未成熟个体,雌雄间角质颚形态不存在显著差异,这与其他的大洋性头足类角质颚特征有所不同[13-15]。此情况产生的原因可能有如下2点:1)由于杜氏枪乌贼栖息于沿岸海域,无论何种性别,其栖息范围均处于一个相似的环境中,因此摄食对象和影响的环境因子均相似,这样雌雄个体间角质颚形态则不会产生较大的差异;2)从个体大小来看,在同一环境中,雌性性成熟与未成熟个体均存在,而雄性则未发现性成熟个体,可以认为雄性和雌性在相同时间处于不同的生长阶段,成熟个体由于食性的变化,角质颚形态已经发生改变,而未成熟个体仍然保持相对稳定的形态。
3.3 角质颚生长特征
本研究发现,角质颚的各项形态值与胴长的关系均符合线性模型,不同性成熟度雌性角质颚的生长也各有不同。这种生长模型较为常见,在其他种类中也有类似的发现[13-15,22]。其生长模型在不同性别中无差异,而在雌性不同性成熟度个体间则表现出明显的差别。在头足类个体发育阶段,个体唯有依靠大量摄食以保证个体和角质颚的生长,因此该阶段无论是个体本身还是角质颚都生长迅速;随着个体的性成熟,所摄入的能量主要供给性腺的发育,因此个体和角质颚的生长就逐渐趋缓。本研究中,雌性个体表现出了2种不同的生长模式,这应该与其生殖策略有关。Rocha等[30]介绍了不同头足类的生殖策略,其中多数大洋性经济头足类 [如柔鱼、茎柔鱼、鸢乌贼 (Symplectoteuthis oualaniensis)] 和枪乌贼类均属于单循环产卵模式(monocyclic spawning),再次细分发现大洋性头足类为多次产卵(multiple spawning),个体在不同的产卵季节间仍旧保持生长;而枪乌贼类为间歇性产卵(intermittent terminal spawning),个体在不同的产卵季节间不再继续生长[30]。正是因为上述原因,直接造成了雌性个体大小相似的情况下,性成熟度大不相同,也影响了角质颚形态的变化。
3.4 不同胴长组和性成熟度雌性角质颚形态变化
雌性杜氏枪乌贼个体不同胴长组之间,角质颚的形态值均存在差异,结合上述性成熟度差异分析后也发现,大多数角质颚形态值受到胴长和性成熟度交互作用的影响。以往的相似研究中均有分胴长组和性成熟度进行分析,均发现了个体和性成熟度对头足类角质颚的影响[13-15,22],但是并没有将2个原因进行综合考虑。本研究中发现相似大小雌性个体呈现出完全不同的性成熟情况,经过分析也发现,个体大小(胴长)和性成熟度存在交互作用,共同影响角质颚参数。这也与上述枪乌贼类的产卵策略有很大的关系[30]。因此,针对杜氏枪乌贼,不能仅仅从角质颚的大小来直接推断出胴长大小或性成熟度的高低。多数学者往往通过建立胴长和角质颚形态值的关系,来推算出未知个体的大小[31-33]。而本研究中出现的特殊情况,采样原有的估算方法往往会造成较大的误差,需要在今后的研究中进一步分析。
本研究分析了性别及性成熟度对南海北部海域杜氏枪乌贼角质颚形态的影响,发现了角质颚形态可能受到个体大小和性成熟度共同作用的影响。杜氏枪乌贼在南海为重要的优势种,其潜在的资源量大,今后有较大的开发潜力。枪乌贼类作为我国重要的近岸种类,有着独特的生长模式和繁殖策略,随着我国近海渔业资源的不断开发,也需要对其进行更深入的了解。本研究中未获得成熟的雄性个体,可能由采样时间较短造成。后续研究应延长采样时间,并建议从环境因子的角度考虑,综合胃含物分析,探究影响杜氏枪乌贼个体成熟和角质颚形态变化的主要原因。
-
![]()
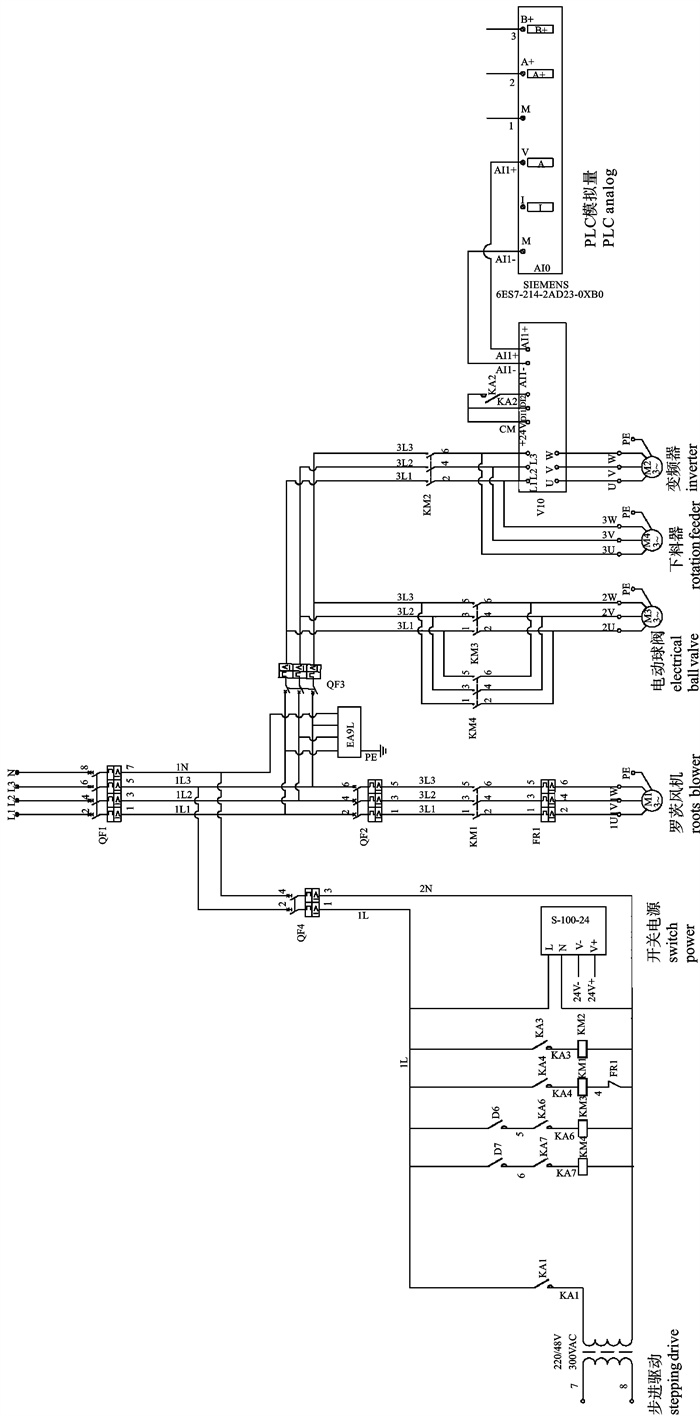
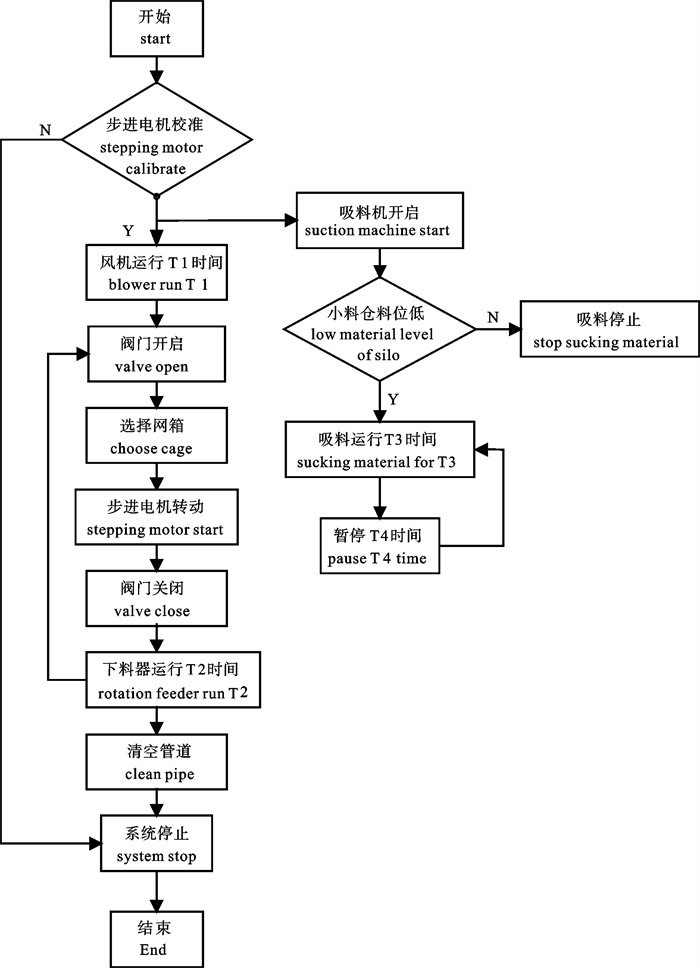
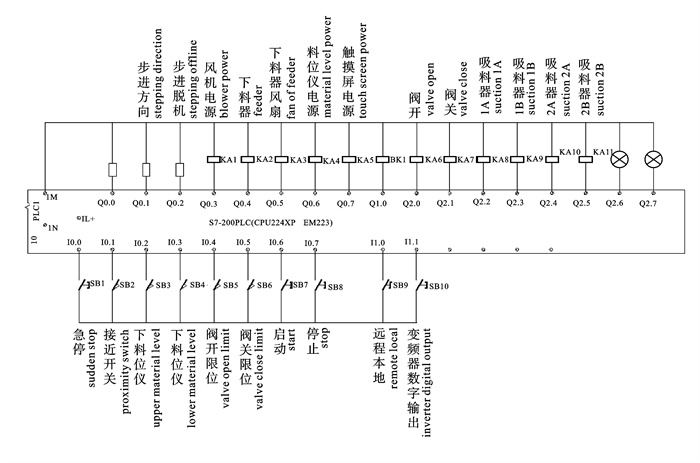
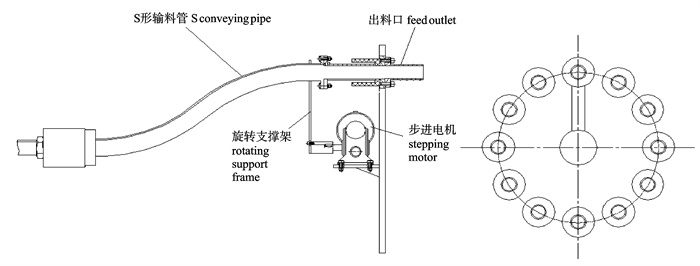
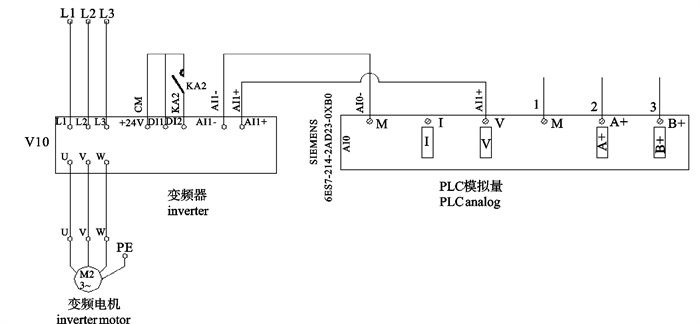
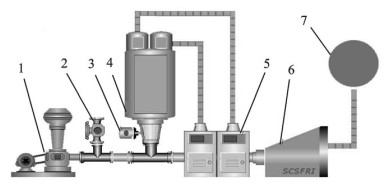
图 1 深水网箱自动投饵系统工作原理图
1. 罗茨风机;2. 球阀;3. 旋转下料器;4. 料仓;5. 吸料器;6. 分配器;7. 深水网箱
Figure 1. Working principle of auto-feeding system for deep-water net cage
1. roots blower; 2. ball valve; 3. rotation feeder; 4. silo; 5. sunction machine; 6. distributor; 7. deep-water net cage
-
[1] 郭根喜, 庄保陆, 王良运, 等. 基于PLC的远程气力输送自动投饵控制系统的设计与实现[J]. 南方水产, 2008, 4(6): 7-16. doi: 10.3969/j.issn.1673-2227.2010.02.001 GUO Genxi, ZHUANG Baolu, WANG Liangyun, et al. Design of automatic control system with remote pneumatic conveying feeding system based on PLC[J]. South China Fish Sci, 2010, 6(2): 1-6. (in Chinese) doi: 10.3969/j.issn.1673-2227.2010.02.001
[2] 郭根喜. 我国深水网箱养殖产业化发展存在的问题与基本对策[J]. 南方水产, 2006, 2(1): 66-70. doi: 10.3969/j.issn.2095-0780.2006.01.014 GUO Genxi. The existing problem and basic countermeasure in the industrialization development of deep water net cage culture in China[J]. South China Fish Sci, 2006, 2(1): 66-70. (in Chinese) doi: 10.3969/j.issn.2095-0780.2006.01.014
[3] 庄保陆, 郭根喜. 水产养殖自动投饵装备研究进展及其应用[J]. 南方水产, 2008, 4(4): 67-72. doi: 10.3969/j.issn.2095-0780.2008.04.013 ZHUANG Baolu, GUO Genxi. The evolution and application of automatic feeding system in aquaculture[J]. South China Fish Sci, 2008, 4(4): 67-72. (in Chinese) doi: 10.3969/j.issn.2095-0780.2008.04.013
[4] 郭根喜, 陶启友. 我国深水网箱养殖技术及发展展望(下)[J]. 科学养鱼, 2004(9): 10-11. https://qikan.cqvip.com/Qikan/Article/Detail?id=10389367 GUO Genxi, TAO Qiyou. Culture technology and development prospect of deep water cage in China[J]. Sci Fish Farming, 2004(9): 10-11. (in Chinese) https://qikan.cqvip.com/Qikan/Article/Detail?id=10389367
[5] 关长涛, 林德芳, 黄滨, 等. 深海抗风浪网箱养殖设施与装备技术的研究进展[J]. 现代渔业信息, 2007, 22(4): 6-8. doi: 10.3969/j.issn.1004-8340.2007.04.002 GUAN Changtao, LIN Defang, HUANG Bin, et al. Progress on research of farming equipment and technology of anti-storm sea net cage[J]. Mod Fish Info, 2007, 22(4): 6-8. (in Chinese) doi: 10.3969/j.issn.1004-8340.2007.04.002
[6] QIESTAD V, PEDERSEN T, FOLKVORD A, et al. Automatic feeding and harvesting of juvenile Atlantic cod (Gadus movhua L. ) in a pond[J]. Model Ident Contral, 1987, 8(1): 39-46. doi: 10.4173/mic.1987.1.5
[7] PAPANDROULAKIS N, DIMITRIS P, PASCAL D. An automated feeding system for intensive hatcheries[J]. Aquac Eng, 2002, 26(1): 13-26. doi: 10.1016/S0144-8609(01)00091-7
[8] CHO C Y, DOMINIQUE P. Reduction of waste output from salmonid aquaculture through feeds and feeding[J]. Prog Fish-Cult, 1997, 59(2): 155-160. doi: 10.1577/1548-8640(1997)059<0155:ROWOFS>2.3.CO;2
[9] MAL B C. Performance of Hawaii-type automated fish feed dispenser[J]. Aquac Eng, 1996, 15(2): 81-90. doi: 10.1016/0144-8609(95)00011-9
[10] ALANARA A. The use of self-feeders in rainbow trout (Oncorhynchus mykiss) production[J]. Aquaculture, 1996, 145(1): 1-20. doi: 10.1016/S0044-8486(96)01346-4
[11] 周晓林, 姬广闻, 焦仁育, 等. 网箱养殖自动控制投饵机的设计与应用[J]. 淡水渔业, 2003, 33(2): 36-37. https://lib.cqvip.com/Qikan/Article/Detail?id=7664000&from=Qikan_Article_Detail ZHOU Xiaolin, JI Guangwen, JIAO Renyu, et al. Design and application of automatic feeding machine in cage culture[J]. Freshwater Fish, 2003, 33(2): 36-37. (in Chinese) https://lib.cqvip.com/Qikan/Article/Detail?id=7664000&from=Qikan_Article_Detail
[12] 宋协法, 路士森. 深水网箱投饵机设计与试验研究[J]. 中国海洋大学学报, 2006, 36(3): 405-409. doi: 10.3969/j.issn.1672-5174.2006.03.013 SONG Xiefa, LU Shisen. Design and experimental study of a new feeder for deep sea cages[J]. Periodic Ocean Univ China, 2006, 36(3): 405-409. (in Chinese) doi: 10.3969/j.issn.1672-5174.2006.03.013
[13] 宋协法. 网箱养殖配套设备的设计与试验研究[D]. 青岛: 中国海洋大学, 2006: 9-15. https://www.doc88.com/p-262184785896.html SONG Xiefa. The farming auxiliary equipments of net cage design and research with experiment[D]. Qingdao: Ocean Univerity of China, 2006: 9-15. (in Chinese) https://www.doc88.com/p-262184785896.html
[14] 刘兆明, 吴子岳. 深水网箱需求式自动投饵装置的初步研究[J]. 上海水产大学学报, 2004, 13(4): 367-370. LIU Zhaoming, WU Ziyue. Preliminary study on the needed automatic feeding equipment in sea-cage[J]. J Shanghai Fish Univ, 2004, 13(4): 367-370. (in Chinese)
[15] 虞宗敢, 高翔, 虞宗勇. 气力投饲系统的研制[J]. 渔业现代化, 2006(2): 45-46. doi: 10.3969/j.issn.1007-9580.2006.02.019 YU Zonggan, GAO Xiang, YU Zongyong. Development of pneumatic feeding system[J]. Fish Modernization, 2006(2): 45-46. (in Chinese) doi: 10.3969/j.issn.1007-9580.2006.02.019
[16] 周晓林, 焦仁育, 胡亚东. 自动投饵系统设计[J]. 渔业现代化, 2005(6): 41-43. doi: 10.3969/j.issn.1007-9580.2005.06.022 ZHOU Xiaolin, JIAO Renyu, HU Yadong. Design of auto feeding system[J]. Fish Modernization, 2005(6): 41-43. (in Chinese) doi: 10.3969/j.issn.1007-9580.2005.06.022
[17] 何伟. 自动投饵机原理及应用[J]. 河南水产, 2004(3): 10-11. HE Wei, The principle and application of auto feeding machine[J]. Henan Fish, 2004(3): 10-11. (in Chinese)
[18] 杨后川, 张瑞. 西门子S7-200 PLC应用100例[M]. 北京: 电子工业出版社, 2009: 88-89. https://xueshu.baidu.com/usercenter/paper/show?paperid=846e8eff9ac93c05fe24d08317e157a1&site=xueshu_se&hitarticle=1 YANG Houchuan, ZHANG Rui. 100 examples of Siemens S7-200 PLC application[M]. Beijing: Publishing House of Electronics Industry, 2009: 88-89. (in Chinese) https://xueshu.baidu.com/usercenter/paper/show?paperid=846e8eff9ac93c05fe24d08317e157a1&site=xueshu_se&hitarticle=1
[19] 李晓宁. 例说西门子PLC S7-200[M]. 北京: 人民邮电出版社, 2008: 9-12. https://xueshu.baidu.com/usercenter/paper/show?paperid=be83aae91253cfb60eb6ad13311f8d0c&site=xueshu_se&hitarticle=1 LI Xiaoning. Example illustration of Siemens PLC S7-200[M]. Beijing: People's Posts & Telecommunications Publishing House, 2008: 9-12. (in Chinese) https://xueshu.baidu.com/usercenter/paper/show?paperid=be83aae91253cfb60eb6ad13311f8d0c&site=xueshu_se&hitarticle=1
[20] 蔡红斌. 电器与PLC控制技术[M]. 北京: 清华大学出版社, 2007: 119-138. CAI Hongbin. Apparatus and PLC control technology[M]. Beijing: Tsinghua University Press, 2007: 119-138. (in Chinese)
-
期刊类型引用(7)
1. 卜心宇,刘必林,欧利国,李建华,方舟,刘岩. 富山武装乌贼个体发育期角质颚表型变化分析. 南方水产科学. 2022(01): 144-152 .  本站查看
本站查看
2. 吕玉晗,张弛,戴了疑,何卫彤,郑小东. 西太平洋杜氏枪鱿(Uroteuthis duvaucelii)群体形态差异分析. 海洋与湖沼. 2022(03): 768-777 . 百度学术
3. 何静茹,陆化杰,陈炫妤,刘凯,王洪浩,陈新军. 西北印度洋鸢乌贼角质颚外形变化的影响因素. 应用生态学报. 2021(05): 1881-1889 . 百度学术
4. 陈炫妤,陆化杰,何静茹,王洪浩,刘凯,陈子越,陈新军. 日本海舍氏贝乌贼角质颚形态特征分析. 动物学杂志. 2021(06): 918-928 . 百度学术
5. 何静茹,陆化杰,陈炫妤,刘凯,王洪浩,陈新军. 冬春季西北印度洋鸢乌贼角质颚外部形态及生长特性. 应用生态学报. 2020(08): 2775-2784 . 百度学术
6. 林仪菲,匡天旭,侯琅,陈伟源,李育森,刘丽,周磊. 珠江流域■和南方拟■的几何形态分析. 淡水渔业. 2020(06): 23-29 . 百度学术
7. 宁欣,陆化杰,张旭,任品,刘凯,陈子越,陈新军. 西北太平洋北方拟黵乌贼角质颚外部形态生长特性. 应用生态学报. 2019(12): 4259-4266 . 百度学术
其他类型引用(5)

 下载:
下载:






计量
- 文章访问数: 4126
- HTML全文浏览量: 153
- PDF下载量: 2437
- 被引次数: 12
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
