
Isolation, identification and phylogenetic analysis of pathogen from Haliotis diversicolor Reeve with withering syndrome
-
摘要:
近几年中国南方杂色鲍(Haliotis diversicolor Reeve)养殖场频繁爆发严重的肌肉萎缩症,该病能感染各种规格杂色鲍,死亡率很高。病鲍症状主要表现为外套膜和内脏团萎缩、无生气,附着力下降,最终死亡。该试验从病鲍组织分离到1株优势菌WS,人工感染试验证实其对健康杂色鲍有很强致病性,且与自然发病的杂色鲍症状相同。对菌株WS进行形态学观察和生理生化检测,结果显示与哈维氏弧菌(Vibrio harveyi)最为接近。为了进一步确定其分类地位,采用1对通用引物扩增其16S rDNA序列片断进行序列分析和同源性对比,构建系统发育树,结果表明,菌株WS与弧菌属的V.harveyi[AY750577]聚为1个分支,且同源性达99.70%。综合上述3种鉴定结果,确定该菌为哈维氏弧菌。
Abstract:In recent years, there have often been serious withering syndrome in many breeding farms of Haliotis diversicolor Reeve in South China. It is infectious to all sizes of abalones with high mortality. The symptoms of sick abalones are: mantle and visceral mass shrink and become lethargic, weak in adhesion and finally die. A bacterial strain WS was isolated from the tissues of cultured H.diversicolor with withering syndrome. Artificial infection test proves that it has strong pathogenicity to healthy H.diversicolor and has symptoms similar to those which get sick spontaneously. We carried out a morphological observation and a test on the physiological and biochemical characteristics of strain WS, which indicates that it is most similar to Vibrio harveyi. In order to define its phylogenetic position, we sequenced, analyzed and compared the 16S rDNA genes whose partial fragments were amplified by a pair of universal primers, and finally constructed a molecular phylogenetic dendrogram based on the genetic distance. The result shows that strain WS and V.harveyi[AY750577] group into one cluster, with a 99.70% homology. According to morphological, physiological, biochemical and phylogenetic analyses, strain WS is proved to be V.harveyi.
-
杂色鲍(Haliotis diversicolor Reeve),又称九孔鲍、珍珠鲍等,是一种热带和亚热带暖水性小型鲍,在中国主要分布于东南沿海省市。中国杂色鲍养殖历史较短,20世纪90年代开始大规模工厂化养殖并迅速发展,成为中国南方海水养殖主要经济品种之一[1]。随着鲍高密度立体养殖模式的推广和生态环境的日益恶化,其病害也接踵而至,并呈现逐年加重的趋势,严重威胁中国鲍鱼养殖业的发展[2-4]。近年来,中国南方杂色鲍养殖场陆续出现一种名为“肌肉萎缩症”的疾病,据笔者调查,该病发生于高温季节,幼鲍及成鲍均可发病,在感染的1~2个月内,养殖场杂色鲍死亡率高达80%以上。症状主要表现为足部肌肉异常消瘦,颜色加深,部分腹足变得僵硬,内脏团和外套膜萎缩,反应较为迟钝,触角伸向壳内,用手轻触即从养殖笼内壁脱落;与正常相同日龄的杂色鲍相比,鲍软体部分与壳长比例严重失调;濒死鲍脱落于养殖笼底部,足肌朝上,但无明显病灶。为了弄清该病的病因,笔者对广东深圳多家患肌肉萎缩症的杂色鲍养殖场进行现场考察,并从病鲍组织分离到1株优势菌,经人工回归感染试验证实其具有很强的致病原性,对该株菌进行形态学观察和常规的生理生化检测,通过测定16S rDNA序列构建系统发育树。
1. 材料与方法
1.1 材料来源
自然发病的杂色鲍取自广东深圳某杂色鲍养殖场,用于病原分离;健康杂色鲍采自另一家鲍养殖场,暂养于玻璃缸供人工回归感染试验用。细菌分离用Zobell 2216E培养基(酵母膏1 g,蛋白胨5 g,磷酸铁0.1 g,氯化钠30 g,蒸馏水1 L,pH 7.6~7.8,琼脂20 g);细菌微量生化鉴定管购自广东环凯生物科技有限公司;16S rDNA扩增通用引物由上海英俊生物技术有限公司合成;细菌基因组DNA提取试剂盒为TIANGEN产品;TaqDNA聚合酶及PMD18-T载体等为TaKaRa产品。
1.2 病鲍解剖观察和病原分离
取不同病程的患肌肉萎缩症杂色鲍,取体表粘液、鳃、肝胰腺和消化道等组织制成水浸片,置于光学显微镜下观察寄生虫和细菌等微生物的情况;将腹足、外套膜、肝胰腺、鳃和消化道加入液氮碾磨后,经梯度离心后制成负染样品,电镜检查有无病毒;在无菌条件下取上述病鲍组织,碾磨成浆,灭菌海水梯度稀释后,涂抹ZoBell 2216E平板,28 ℃培养48 h,把形态特征一致的优势菌落重复划线纯化,直至获得1株纯培养菌(编号WS),接种于斜面培养待用。
1.3 人工感染试验
菌株WS经纯化培养,挑取单菌落接种至Dobell 2216E液体培养基,28 ℃生化培养箱振荡培养24 h,离心后采用活菌平板计数和比浊法,利用过滤海水将活菌浓度调至2.5×106 cfu · mL-1。试验共分3组,每组20只健康杂色鲍,放入预先由患病杂色鲍体内分离到的优势菌株WS制备的2.0×106 cfu · mL-1菌液中浸泡养殖,同时设平行组,每日更换新鲜海水并添加菌液,以保持养殖水体中菌体浓度,并设不加菌对照组。感染期间不投喂饲料,全天候充气,水温控制在(25±1)℃,盐度25~30,时间2周。每天记录鲍鱼的发病情况和死亡数量,若有濒死鲍,则进行病原菌再分离,并挑选与原感染病原菌一致的菌株进行再感染试验。
1.4 病原菌形态学观察和生理生化特征检测
将优势菌株WS划线接种于TCBS平板培养基,28 ℃培养24 h观察生长情况和菌落形态,同时进行革兰氏染色和电镜负染,观察菌体形态特征,其他各项生理生化指标的测定参照文献[5]进行。
1.5 病原菌16S rDNA序列测定和分析
1.5.1 PCR模板的制备
按照TIANGEN公司的细菌基因组DNA提取试剂盒说明书进行总DNA制备,并取DNA溶液进行琼脂糖凝胶电泳检查制备的DNA完整性,若符合要求置于-20 ℃保存备用。
1.5.2 16S rDNA序列扩增、克隆及测序
用于16S rDNA扩增的通用引物,其正向引物8F: 5’-A-GAGTTTGATCCTGGCTCAG-3’ (对应于E.coli 16S rDNA第8~27个碱基位置),反向引物1492R: 5’ -GGTTACCTTGTTACGACTT-3’ (对应于E.coli 16S rDNA的第1 492~1 510个碱基位置)[6]。24 μL PCR反应体系中:10×PCR缓冲液2.5 μL;MgCl2 1.5 mmol · L-1;4×dNTP混合物各200 μmol · L-1;引物各10 μmol · L-1;TaqDNA聚合酶0.5 μL(2.5 U · μL-1)和DNA模板1 μL。PCR反应条件:94 ℃预变性5 min;94 ℃变性45 s;52 ℃复性40 s;70 ℃延伸1 min,35个循环,最后70 ℃温育10 min。琼脂糖凝胶电泳获得特异性条带,凝胶产物回收后经进一步PCR验证与pMD18-T载体连接,转化至E.coli JM109感受态细胞进行培养,经蓝白斑筛选,挑取若干白色菌落至液体培养基过夜培养,提取重组质粒,酶切和PCR验证后,产物交由上海英俊生物技术公司进行纯化和测序。
1.5.3 序列分析和系统发育树构建
将获得的16S rDNA基因序列提交到GenBank核苷酸序列数据库进行同源序列检索,选取同源性较高的已知菌株,调出其16S rDNA基因序列,用DNAstar软件进行多序列对比和系统发育树构建。
2. 结果
2.1 病鲍解剖观察和病原分离结果
病鲍各组织的水浸片显微观察结果显示,大多数病鲍鳃、消化道和肝胰腺组织处有活动的短杆状细菌存在,另外除发现少量纤毛虫和车轮虫外,未见其他微生物存在。多次电镜负染结果均未发现病毒粒子。利用平板涂抹法从病鲍组织分离到1株优势菌株,经多次纯化后接种于斜面保存,并利用优势菌株WS进行人工感染试验。
2.2 人工感染试验
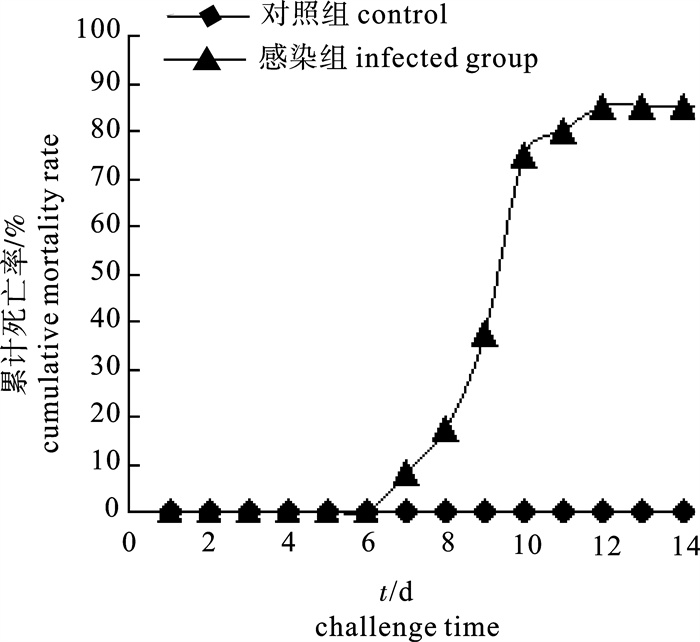
WS菌液浸泡感染期间,杂色鲍从第5天开始出现发病症状,表现为运动缓慢,反应迟钝,附着力下降,腹足和外套膜萎缩,内脏团收缩,养殖水也变得较为混浊;第7天开始部分鲍鱼从水族箱内壁脱落,腹足朝上,最终死亡。后期死亡的杂色鲍萎缩症状非常明显,与自然发病症状相似,对照组无发病和死亡情况。从濒死鲍体内分离到菌株WSA和再感染分离到菌株WSA1,其菌落和菌体形态均与从自然发病杂色鲍体内分离到的菌株WS一致,充分证明菌株WS为此次南方杂色鲍肌肉萎缩症的病原菌(浸泡感染试验结果见图 1)。
2.3 菌形态特征和理化特性
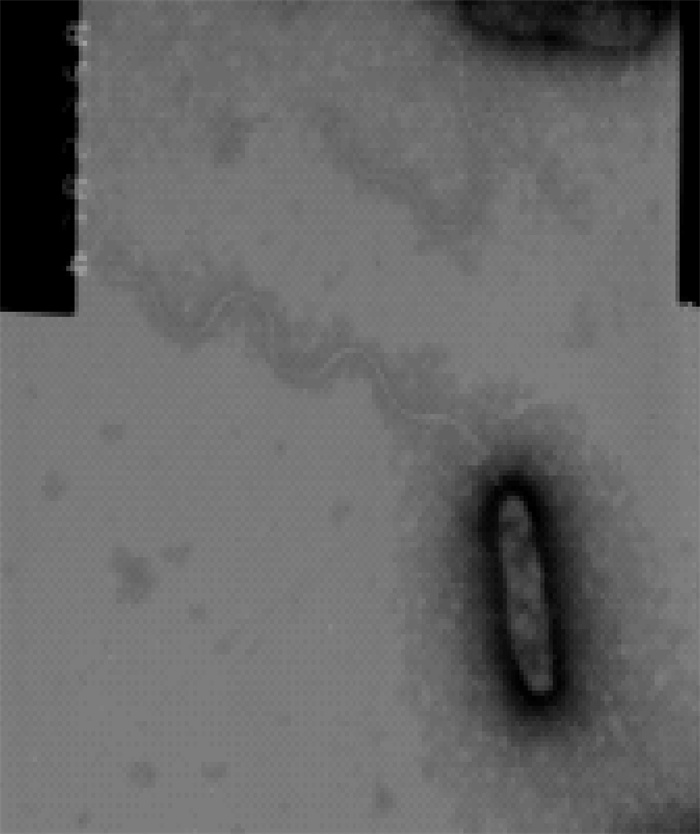
菌株WS在TCBS培养基上生长良好,菌落呈绿色,圆形,边缘整齐,表面光滑湿润,稍隆起;革兰氏染色阴性,短杆状,稍弯曲;电镜负染极生单鞭毛(图 2)。各项生理生化反应结果见表 1。
表 1 菌株WS的生理生化特性与哈维氏弧菌标准菌株ATCC14126[7]比较Table 1 Comparison of physiological and biochemical characteristics between WS and V.harveyi ATCC14126鉴定项目
test itemsWS V.harveyi
ATCC14126鉴定项目
test itemsWS V.harveyi
ATCC14126革兰氏染色gram stain ( ( 明胶酶gelatinase ( + 运动性motility + + 尿素酶urease ( ( 氧化酶oxidase + + 淀粉酶amylase ( + 接触酶catalase + + 脂酶lipase + + O/129(150 μg) s s 几丁质酶chitinase + + 发光luminescence ( ( H2S ( ( TCBS生长growth on TCBS g g V-P test ( ( 色素产生pigmentation ( ( 水杨素salicin ( ( 0%NaCl生长growth at 0%NaCl ( ( 吲哚indole ( ( 3%NaCl生长growth at 3%NaCl + + 柠檬酸盐citrate + + 6%NaCl生长growth at 6%NaCl + + 肌醇inosito ( ( 8%NaCl生长growth at 8%NaCl + + 纤维二糖cellobio ( + 10%NaCl生长growth at 10%NaCl ( ( 阿拉伯糖arabinose ( ( 4 ℃生长growth at 4 ℃ ( ( 木糖xylos ( ( 30 ℃生长growth at 30 ℃ + + 蔗糖saccharose ( ( 35 ℃生长growth at 35 ℃ + + 乙醇ethanol ( ( 42 ℃生长growth at 40 ℃ ( ( 葡萄糖glucose + + β-半乳糖苷β-galactosid + + 甘露糖mannose + + 葡萄糖产气gas from glucose ( ( 甘露醇mannitol + + 硝酸盐还原nitrate reduction + + 乳糖lactose ( ( 赖氨酸脱羧酶lyadeearboxylaes + + 蜜二糖melibiose ( ( 鸟氨酸脱羧酶ornithine decarboxylase ( + 鼠李糖rhamnose ( ( 精氨酸双水解酶arginine dihydrolase ( ( 山梨糖sorbitol ( ( 色氨酸脱氨酶tryptophane deaminase ( ( 半乳糖galactose ( + 注: +.阳性;(.阴性;S.敏感;G.绿色
Note: +.positive; (.negative; S.sensitive; G.green2.4 16S rDNA序列测定和系统发育树构建
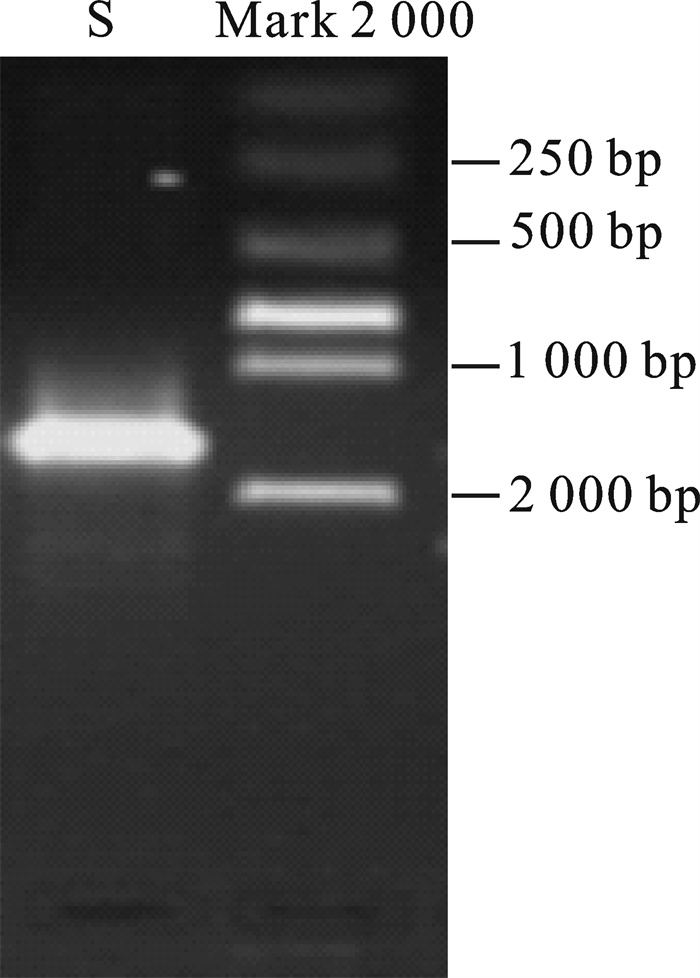
DNA模板经16S rDNA通用引物扩增,琼脂糖凝胶电泳得到约1 500 bp条带(图 3),PCR产物凝胶回收纯化后经载体连接、转化后扩增培养,提取重组质粒,阳性克隆双酶切后电泳得到2条带(图 4)。
![]() 图 4 重组质粒双酶切验证P.酶切质粒Fig. 4 Restriction digestion identification of recombinant plasmid P.Plasmid of restriction digestion
图 4 重组质粒双酶切验证P.酶切质粒Fig. 4 Restriction digestion identification of recombinant plasmid P.Plasmid of restriction digestion挑取阳性克隆经双向测序得到的16S rDNA片段长度为1 548 bp,登陆GenBank数据库进行同源性检索,登陆序号为lcl|2069,结果与弧菌属细菌的16S rDNA基因序列自然聚类。在相近的100个检索结果中,弧菌占89%。从中选取同源性较高的弧菌属菌株,从GenBank下载其16S rDNA序列,用DNAstar软件构建系统树(图 5)。结果表明该菌与V.harveyi[AY750577]聚为1个分支,其相似性达99.70%,结合形态学和生理生化特征鉴定其为哈维氏弧菌(Vibrio harveyi)。
3. 讨论
笔者对多家患肌肉萎缩症的杂色鲍养殖场进行现场调查发现,杂色鲍发病期间养殖海水的温度、盐度、pH、溶解氧和氨氮(NH3-N)等各种理化因子与以往养殖周年相比较,均未出现异常变化,重金属含量检测也未超出水质检测标准,因此,基本可以排除非生物因素作为杂色鲍发病主要病因的可能。中国南方杂色鲍肌肉萎缩症爆发期为每年的8~9月,此时期养殖水温较高,哈维氏弧菌代谢旺盛、生长迅速[8],粘附作用相关基因的表达能力和毒力基因表达产物的活性随之增强[9],加之现今的高密度立体养殖模式导致鲍抵抗力减弱,当水中致病菌含量达到一定值时便可能引起鲍鱼发病。BEVERLY等[10]认为升高水温能显著加速鲍肌肉萎缩症的进程,而环境因素不会直接引起鲍发病,而厄尔尼诺现象、不合理的饲养管理和环境的污染也可加速肌肉萎缩症进程,由此可见,环境因素对杂色鲍肌肉萎缩症的发生起外在诱导作用。
2005年5月,中国北方养殖的皱纹盘鲍(H.discus hannai Ino)发生肌肉萎缩症,造成大规模死亡[11]。近几年来,中国南方杂色鲍养殖场陆续爆发肌肉萎缩症,死亡率很高,许多养殖场面临关闭的困境,给鲍养殖业造成巨大经济损失。笔者对患病杂色鲍进行解剖观察,并从体内分离到1株优势菌WS,首先对菌株WS进行形态学观察和生理生化测定,与哈维氏弧菌标准菌株做比较,结果在是否发光、尿氨酸脱羧酶、淀粉酶、明胶酶、纤维二糖和半乳糖等几个方面存在差异,无法鉴定到种。目前,PCR技术广泛应用于水产动物细菌性病原鉴定,16S rDNA分子量适中,具有普遍性、保守性和相对可变性等特点,序列变化与进化距离相适应,序列分析的重复性极高,在细菌分子鉴定中应用普遍[12-13]。因此,对菌株WS的16S rDNA进行PCR扩增、克隆和测序,与相似菌株的16S rDNA序列进行同源性比较和分析,并构建系统发育树,结果与V.harveyi[AY750577]聚为1个分支,且同源性达99.70%,最终将该菌株鉴定为哈维氏弧菌。
哈维氏弧菌广泛分布于自然海水环境和养殖海水环境,并以养殖环境的数量居多,是海水养殖动物的重要致病原[14],由于可遗传的毒力因子不同,哈维氏弧菌不同菌株间的致病力存在很大差异[15]。THOMPSON等[16]研究证实,哈维氏弧菌在暖水季节数量尤为丰富,通过入侵机体引起生理机能发生紊乱导致生物体发病。对杂色鲍肌肉萎缩症进行病理学研究时发现,病鲍的肝胰腺、鳃和肠道均呈现不同程度的病变,这使鲍正常的消化、吸收和呼吸等生理功能受到障碍,新陈代谢能力减弱,最终死亡[17]。在国外,哈维氏弧菌曾引起日本一家皱纹盘鲍养殖场大规模发病,鲍发病前除摄食量减少外,无其他征兆,该病集中发生在每年的8、9月,水温低于20 ℃时不会发生[18]。另有报道哈维氏弧菌引起法国自然海域欧洲鲍(H. tuberculata)大量死亡[19]。到目前为止,国内未见哈维氏弧菌引起鲍发病的相关报道,此次从患肌肉萎缩症杂色鲍组织分离到哈维氏弧菌,并通过人工感染试验证实其对健康鲍具有很强致病性,在国内尚属首例,说明哈维氏弧菌对中国鲍养殖业具有潜在危害性,应引起相关部门和水产养殖业的重视。
-
![]()
图 4 重组质粒双酶切验证P.酶切质粒
Figure 4. Restriction digestion identification of recombinant plasmid P.Plasmid of restriction digestion
表 1 菌株WS的生理生化特性与哈维氏弧菌标准菌株ATCC14126[7]比较
Table 1 Comparison of physiological and biochemical characteristics between WS and V.harveyi ATCC14126
鉴定项目
test itemsWS V.harveyi
ATCC14126鉴定项目
test itemsWS V.harveyi
ATCC14126革兰氏染色gram stain ( ( 明胶酶gelatinase ( + 运动性motility + + 尿素酶urease ( ( 氧化酶oxidase + + 淀粉酶amylase ( + 接触酶catalase + + 脂酶lipase + + O/129(150 μg) s s 几丁质酶chitinase + + 发光luminescence ( ( H2S ( ( TCBS生长growth on TCBS g g V-P test ( ( 色素产生pigmentation ( ( 水杨素salicin ( ( 0%NaCl生长growth at 0%NaCl ( ( 吲哚indole ( ( 3%NaCl生长growth at 3%NaCl + + 柠檬酸盐citrate + + 6%NaCl生长growth at 6%NaCl + + 肌醇inosito ( ( 8%NaCl生长growth at 8%NaCl + + 纤维二糖cellobio ( + 10%NaCl生长growth at 10%NaCl ( ( 阿拉伯糖arabinose ( ( 4 ℃生长growth at 4 ℃ ( ( 木糖xylos ( ( 30 ℃生长growth at 30 ℃ + + 蔗糖saccharose ( ( 35 ℃生长growth at 35 ℃ + + 乙醇ethanol ( ( 42 ℃生长growth at 40 ℃ ( ( 葡萄糖glucose + + β-半乳糖苷β-galactosid + + 甘露糖mannose + + 葡萄糖产气gas from glucose ( ( 甘露醇mannitol + + 硝酸盐还原nitrate reduction + + 乳糖lactose ( ( 赖氨酸脱羧酶lyadeearboxylaes + + 蜜二糖melibiose ( ( 鸟氨酸脱羧酶ornithine decarboxylase ( + 鼠李糖rhamnose ( ( 精氨酸双水解酶arginine dihydrolase ( ( 山梨糖sorbitol ( ( 色氨酸脱氨酶tryptophane deaminase ( ( 半乳糖galactose ( + 注: +.阳性;(.阴性;S.敏感;G.绿色
Note: +.positive; (.negative; S.sensitive; G.green 下载: 导出CSV
下载: 导出CSV
-
[1] 苏天凤. 杂色鲍与九孔鲍种质资源研究进展[J]. 南方水产, 2006, 2(2): 8-12. doi: 10.3969/j.issn.2095-0780.2006.02.012 [2] 李太武, 丁明进, 相建海, 等. 皱纹盘鲍对河流弧菌-Ⅱ苗免疫的研究[J]. 海洋与湖沼, 1997, 28(1): 27-32. [3] FRIEDMAN C S, GRINDLEY R, KEOGH J A. Isolation of a fungus from shell lesions of New Zealand abalone, Haliotis iris Martyn and H. australis Gmelin[J]. Mollus Res, 1997, 18: 313-324. doi: 10.1080/13235818.1997.10673704
[4] WANG Jiangyong, GUO Zhixun, FENG Juan, et al. Virus infection in cultured abalone, Haliotis diversicolor Reeve in Guangdong province, China[J]. J Shellfish Res, 2004, 23(4): 1163-1168. https://eurekamag.com/research/012/742/012742327.php
[5] 东秀珠, 蔡妙英. 常见细菌系统鉴定手册[M]. 北京: 科学出版社, 2001: 364-398. https://xueshu.baidu.com/usercenter/paper/show?paperid=92be5766cfb591d39903d20547aabf25&site=xueshu_se&hitarticle=1 [6] POLZ M F, CAVANAUGH C M. Bias in template-to-product ratios in multitemplate PCR[J]. Appl Environ Microbiol, 1998, 64(10): 3724-3730. doi: 10.1128/AEM.64.10.3724-3730.1998
[7] LIU P C, LEE K K, YII K C, et al. Isolation of Vibrio harveyi from diseased Kuruma prawn Penaeus japonicus[J]. Curr Microbiol, 1996, 33(2): 129-133. doi: 10.1007/s002849900087
[8] LOPEZ L M, TYLER P A, VIANA M T. The effect of temperature and artificial diets on growth rates of juvenile Haliotis tuberculate[J]. J Shellfish Res, 1998, 17(3): 657-662. https://www.researchgate.net/publication/236222880_The_effect_of_temperature_and_artificial_diets_on_growth_rates_of_juvenile_Haliotis_tuberculata_Linnaeus_1758
[9] GOARANT C, HERLIN J, BRIZARD R, et al. Toxic factors of Vibrio strains pathogenic to shrimp[J]. Dis Aquat Org, 2000, 40(2): 101-107. doi: 10.3354/dao040101
[10] BEVERLY A B, JAMES D M, THEA T R. Health and survival of red abalone, Haliotis rufescens, under varying temperature, food supply, and exposure to the agent of withering syndrome[J]. Invertebr Pathol, 2005, 89(3): 219-231. doi: 10.1016/J.JIP.2005.06.004
[11] 于金海, 王品虹, 李春艳, 等. 皱纹盘鲍"足萎缩症"幼鲍超微结构研究[J]. 海洋环境科学, 2007, 26(5): 461-465. https://xueshu.baidu.com/usercenter/paper/show?paperid=1r6j0p802p1g04c03w7p0p60xv141459&site=xueshu_se&hitarticle=1 [12] YOSHIVUKI Y, YOSHIKO K, HISATSUGU W. Phylogenetic intrarelationshlips of a typical Aeramonas salmonicida isolated in Japan as determined by 16S rDNA sequencing[J]. Fish Pathol, 2000, 35(1): 35-40. https://www.jstage.jst.go.jp/article/jsfp1966/35/1/35_1_35/_pdf/-char/ja
[13] GAUGER E G, GOMEZ C M. 16S ribosomal DNA sequencing confirms the synonymy of Vibrio harveyi and V. carchariae[J]. Dis Aquat Org, 2002, 52(1): 39-46. doi: 10.3354/dao052039
[14] 陈吉祥, 杨慧, 杨显辉, 等. 致病性哈维氏弧菌溶血素基因克隆及其检测[J]. 中国水产科学, 2005, 12(5): 580-587. doi: 10.3321/j.issn:1005-8737.2005.05.009 [15] MONTERO A B, AUSTIN B. Characterization of extracellular products from an isolate of Vibrio harveyi recovered from diseased post-larval Peneaus vannamei (Bonne)[J]. J Fish Dis, 1999, 22(5): 377-386. doi: 10.1046/j.1365-2761.1999.00189.x
[16] THOMPSON J R, RANDA L A, MARCELINO A, et al. Diversity and dynamics of a North Atlantic coastal Vibrio community[J]. Appl Environ Microbiol, 2004, 70: 4103-4110. doi: 10.1128/AEM.70.7.4103-4110.2004
[17] 孙秀秀, 苏友禄, 冯娟, 等. 杂色鲍肌肉萎缩症的组织病理学研究[J]. 安徽农业科学, 2009, 37(3): 1098-1101. doi: 10.3969/j.issn.0517-6611.2009.03.082 [18] NISHIMORI E, HASEGAWA O, NUMATA T, et al. Vibrio carcharia causes mass mortalities in Japanese abalone, Sulculus diversicolor supratexta[J]. Fish Pathol, 1998, 33(4): 495-502. doi: 10.3147/JSFP.33.495
[19] NICHOLAS J L, BASUYAUX O, MAZURIE J, et al. Vibrio carchariae, a pathogen of the abalone Haliotis tuberculata[J]. Dis Aquat Org, 2002, 50(1): 35-43. doi: 10.3354/dao050035
-
期刊类型引用(17)
1. 熊向英,曾梦清,陈瑞芳. 盐酸多西环素和恩诺沙星对织锦巴非蛤肠道微生物的影响. 南方农业学报. 2023(01): 273-280 .  百度学术
百度学术
2. 王瑞旋,王江勇,李韵萍,郭子晗,李炳,李云,朱慧. 致病性海洋弧菌对氨基糖苷类药物的耐药传递机制初步研究. 海洋科学. 2020(10): 81-90 . 百度学术
3. 梁健,高山,李永仁,郭永军,黄亚东,邢克智. 方斑东风螺肿吻病致病菌分离及中草药对其的预防效果. 江苏农业科学. 2016(11): 267-269 . 百度学术
4. 程蝶,柴方超,蔡怡,周前进,陈炯. 环介导等温扩增联合横向流动试纸条可视化检测哈维氏弧菌的研究. 生物技术通报. 2016(06): 60-68 . 百度学术
5. 赵曼曼,姜敬哲,何健,孙永婵,王江勇. 哈维弧菌和鲍类疱疹病毒刺激对杂色鲍免疫相关因子的影响. 海洋科学. 2015(11): 39-48 . 百度学术
6. 辜良斌,徐力文,冯娟,苏友禄,刘广峰,郭志勋. 豹纹鳃棘鲈尾部溃烂症病原菌的鉴定与药敏试验. 南方水产科学. 2015(04): 71-80 . 本站查看
7. 高玉龙,冯娟,苏友禄,徐力文,郭志勋. 广东阳江市闸坡网箱养殖区弧菌耐药性分析. 海洋环境科学. 2014(01): 32-35 . 百度学术
8. 杨蕊,姜敬哲,王江勇,赵旺. 杂色鲍低温病毒病研究进展. 广东农业科学. 2013(02): 114-117+2 . 百度学术
9. 韩焘,姜敬哲,王江勇. 杂色鲍血淋巴双向电泳与质谱分析. 海洋科学. 2013(03): 102-108 . 百度学术
10. 房沙沙,林壮炳,邱礽,王劭雯,刘晓. 粤东养殖区分离的2株海洋弧菌及其胞外产物对皱纹盘鲍致死毒性的初步分析. 海洋科学. 2013(08): 16-22 . 百度学术
11. 王江勇,王瑞旋,苏友禄,吴开畅,郭志勋,姜敬哲,刘广锋,赵旺. 方斑东风螺“急性死亡症”的病原病理研究. 南方水产科学. 2013(05): 93-99 . 本站查看
12. 黄贻涛,蔡秀红,张子平,王国栋,邹志华,王淑红,王艺磊. 杂色鲍紫色酸性磷酸酯酶基因克隆及应激下的表达. 中国水产科学. 2013(05): 939-949 . 百度学术
13. 姜敬哲,张彬彬,张远,苏友禄,王江勇. 杂色鲍Fosmid基因组文库构建与血蓝蛋白基因筛选. 华南农业大学学报. 2012(02): 192-196+202 . 百度学术
14. 王瑞旋,耿玉静,王江勇,冯娟,李国平. 水产致病菌耐药基因的研究. 海洋环境科学. 2012(03): 323-328 . 百度学术
15. 姜敬哲,韩焘,王江勇,杨慧英,刘金叶. 杂色鲍血蓝蛋白多克隆抗体的制备与血蓝蛋白35kDa片段鉴定. 海洋科学. 2012(03): 67-73 . 百度学术
16. 王瑞旋,耿玉静,冯娟,王江勇. 杂色鲍哈维弧菌耐药质粒的鉴定和分析. 南方水产科学. 2012(02): 1-6 . 本站查看
17. 张岩,吴燕燕,李来好,杨贤庆,邵征翌. 16S rDNA序列分析鉴定一株合浦珠母贝共附生乳酸菌. 南方水产科学. 2012(06): 9-15 . 本站查看
其他类型引用(22)




计量
- 文章访问数: 4791
- HTML全文浏览量: 192
- PDF下载量: 3044
- 被引次数: 39
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
