
Influence of illumination,oxygen,pH and salinity on growth and nitrite removal effect of Rhodopseudomonas palustris strain 2-8
-
摘要:
以1株具有亚硝酸盐消除能力的沼泽红假单胞菌(Rhodopseudomonas palustris)2-8菌株(简称2-8菌株)为材料, 研究了不同光照和氧组合、pH、盐度对菌株生长和亚硝酸盐消除能力的影响。结果发现, 光照厌氧条件最利于2-8菌株的生长, 该条件下菌株亚硝酸盐消除能力最强, 25h消除率达(91.33±1.27)%;菌株在pH7.0时生长和亚硝酸盐消除能力最强, 25h亚硝酸盐消除率达到(95.58±4.34)%, 在pH9.0以上和5.0以下时基本不生长;菌株在w(NaCl)为0和0.4%的培养基中生长和亚硝酸盐消除能力最强, 在w(NaCl)为0.8%~2.0%的培养基中, 生长和亚硝酸盐消除能力随w(NaCl)增高而减弱。测定的4个环境因子主要通过影响菌株的生长来影响其对亚硝酸盐的消除能力。
Abstract:We used Rhodopseudomonas palustris strain 2-8, a denitrifying non-sulfur photosynthetic bacterium, to study its growth and nitrite removal effect with different illumination-oxygen combinations, pH level and NaCl concentrations.The results show that illumination without oxygen is best for the strain′s growth and nitrite removal, with a nitrite removal efficiency of (91.33±1.27)% after 25 h inoculation;pH 7.0 is best for its growth and nitrite removal, and the nitrite removal efficiency reaches (95.58±4.34)% after 25 h inoculation, however, it does not grow at pH below 5.0 or above 9.0.Moreover, the strain grows best and removes nitrite most quickly with NaCl culture medium of 0.0% or 0.4%, but with a 0.8%~2.0% NaCl concentration, its growth and nitrite removal effect weaken with the increasing concentration.These four environmental factors influence the nitrite removal effect of stain 2-8 by influencing its growth.
-
Keywords:
- nitrite removal /
- Rhodopseudomonas palustris /
- growth
-
亚硝酸盐是水产养殖中一种重要的污染物,容易积累甚至严重超标而引发养殖动物病害或死亡[1-2]。亚硝酸盐可以引起水产养殖动物呼吸作用、离子调节、心血管压力调节、内分泌及排泄等多种生理代谢紊乱[3],低质量浓度时对凡纳滨对虾(Litopenaeus vannanei)的影响则是降低其免疫能力[4]。亚硝酸盐在自然界的消除主要通过具有硝化能力的自养细菌和具有反硝化作用或产铵异化硝酸盐还原能力的异养细菌来完成[5]。光合细菌中的沼泽红假单胞菌(Rhodopseudomonas palustris)是一种新陈代谢机制多样的紫色非硫细菌,广泛存在于土壤和水体中[6],具有编码同化和异化亚硝酸盐还原酶的基因[7],能够消除亚硝酸盐[8],并在畜禽饲养、水产养殖、农业种植、污水处理、医疗保健、能源和新材料等领域广泛应用[9-12],水产养殖中主要用于控制水体化学需氧量(chemical oxygen demand,COD)、氨氮和亚硝酸盐[8, 13-14]。
珠海市农业科学研究中心的1株专利菌株沼泽红假单胞菌2-8菌株(R.palustris strain 2-8,简称2-8菌株)具有很强的亚硝酸盐消除能力,从2007年开始在斗门六乡应用于高密度凡纳滨对虾养殖生产实践,通过持续的田间应用证实其不仅能够有效净化养殖水体的亚硝酸盐,还具有增产的效果,2007~2008年冬棚养殖对虾6个月的单产折合14 844 kg · hm-2,2008年露天养殖对虾5个月的产量折合11 235 kg · hm-2(未发表数据)。由于各地养殖水体存在差异,水体条件如光照、溶氧、盐度和pH等会在不同的养殖时间或时期发生变化,而这些因素会对沼泽红假单胞菌的生长[15-17]和氮、磷消除功能产生影响[18-20]。为了探讨养殖水体光照、氧气、盐度和pH等生态因子波动对2-8菌株生长和亚硝酸盐消除的影响,了解菌株的适用水环境范围以扩大其田间应用,笔者就相关问题在室内开展了模拟研究。
1. 材料与方法
1.1 试验材料
1.1.1 供试菌株
2-8菌株由珠海市农业科学研究中心农业微生物学实验室保藏。
1.1.2 培养基和检测试剂
参考文献[9]配制Van Niel(VN)培养基,w (NH4 Cl) 0.1%,w (NaCl)0.2%,w (Na2HPO4) 0.05%,w (MgCl2)0.02%,w (蛋白胨) 0.2%,将各组分溶解到水中,121 ℃蒸汽灭菌20 min,分别制备下列溶液并过滤除菌,1)w(NaHCO3)10%;2)乙醇;3)0.1 mol · L-1 H3PO4。在基础培养基中加入NaHCO3溶液50 mL,乙醇2.0 mL,并用H3PO4调节pH 7.0。参考文献[21]配制Griess试剂用于亚硝酸盐测定。
1.2 试验方法
1.2.1 光照和氧气组合对菌株生长和亚硝酸盐消除的影响
光照和氧气组合方式参考张李阳和吴向华[20]的方法,包括光照厌氧、光照有氧、黑暗厌氧和黑暗有氧4个处理。试验开始时,将菌浓度为1010 cfu · mL-1的2-8菌株种子液使用新鲜VN培养基稀释到106 cfu · mL-1并分装,其中厌氧处理用磨口三角瓶中加满培养液混合物模拟,有氧处理用50 mL三角瓶中加入25 mL混合物模拟;黑暗处理用锡箔纸和黑色塑料袋包裹三角瓶模拟。每个处理设3个重复,在30 ℃、光照强度1 800 lx的恒温箱中培养,每隔24 h测定1次不同处理培养液的光密度(OD660),以未接菌的VN培养基为空白对照调零。亚硝酸盐消除生测参考喻国辉等[22]的方法,组合方式、菌体初始浓度和培养条件同上,仅在体系中添加亚硝酸钠使其终质量浓度为2.4 mg ·L-1以模拟亚硝酸盐胁迫,培养16 h后开始检测培养基中亚硝酸盐剩余值,根据剩余值计算消除的亚硝酸盐值和消除率。
1.2.2 pH对菌株生长和亚硝酸盐消除的影响
生长和亚硝酸盐消除生测体系和方法同光照厌氧处理,用1 mol · L-1的NaOH溶液和HCl溶液将体系的pH分别调节为3.0、4.0、5.0、6.0、7.0、8.0、9.0和10.0,在30 ℃、光照强度1 800 lx的恒温箱中培养,并按时测定OD660和亚硝酸盐消除情况。
1.2.3 盐度对菌株生长和亚硝酸盐消除的影响
生长和亚硝酸盐消除生测体系和方法同光照厌氧处理,使用NaCl母液将体系的w (NaCl)分别调整为0、0.2%、0.6%、0.8%、1.0%、1.2%、1.6%和2.0%以模拟盐度梯度,培养条件和测定方法同上。
1.2.4 体系亚硝酸盐的测定
测定时吸取1.5 mL培养液到2 mL离心管中,离心去除菌体,上清吸到玻璃试管中,加入适量固体Griess试剂显色5 min,测定OD520,以不加菌、其他条件相同的处理为对照校正系统误差,以不含亚硝酸盐的培养基加入Griess试剂调零。
$$ \text { 亚硝酸盐消除率(%) }=\left(\frac{\rho(\text { 对照亚硝酸盐 })-\rho(\text { 处理亚硝酸盐 })}{\rho(\text { 时照亚硝酸盐 })}\right) \times 100 \% $$ 1.3 数据分析
将获得的数据在SPSS 13.0软件包中利用单因素方差分析和LSD多重比较,分析不同处理下菌株的生长差异和对亚硝酸盐消除率的差异,并在Excel软件中作图。
2. 结果与分析
2.1 光照和氧气组合对菌株生长和亚硝酸盐消除的影响
2.1.1 光照和氧气对2-8菌株生长的影响
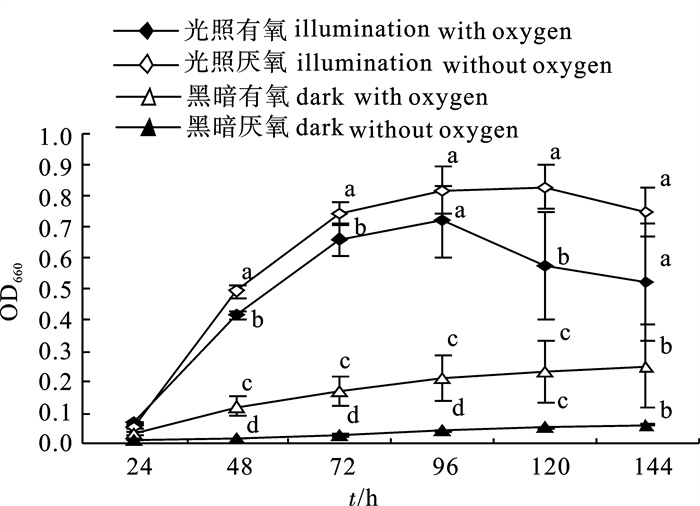
在光照厌氧条件下,2-8菌株生长最好,最高OD660达0.83±0.07,其次是光照有氧,而在黑暗条件下生长速率都显著低于光照条件(图 1)。显示光照有利于2-8菌株的生长。
![]() 图 1 光照和氧气组合对2-8菌株生长的影响注:同一时间不同处理上的小写字母表示处理间SPSS方差分析和LSD多重比较差异显著(P<0.05),后图同此Fig. 1 Influence of different illumination-oxygen combinations on growth of strain 2-8Note:Different lowercase letters on the same time column indicate different treatments influence the strain′s growth significantly (P<0.05, LSD, SPSS 13.0). The same case in the following figures.
图 1 光照和氧气组合对2-8菌株生长的影响注:同一时间不同处理上的小写字母表示处理间SPSS方差分析和LSD多重比较差异显著(P<0.05),后图同此Fig. 1 Influence of different illumination-oxygen combinations on growth of strain 2-8Note:Different lowercase letters on the same time column indicate different treatments influence the strain′s growth significantly (P<0.05, LSD, SPSS 13.0). The same case in the following figures.2.1.2 光照和氧气对2-8菌株亚硝酸盐消除能力的影响
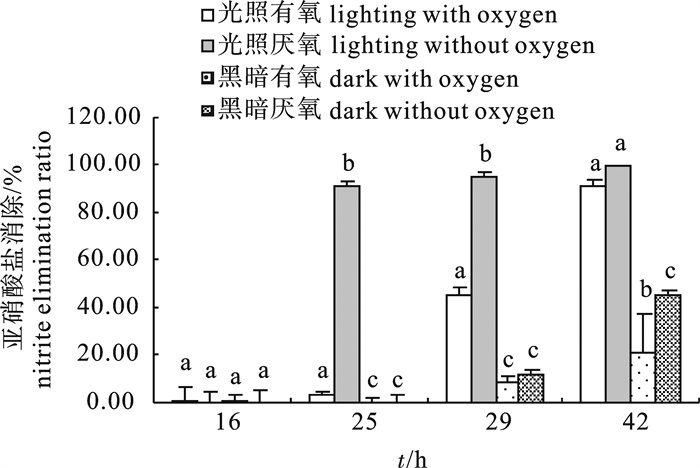
光照和氧的不同组合对2-8菌株的亚硝酸盐消除能力影响显著(图 2),光照厌氧条件最利于菌株消除亚硝酸盐,在此条件下,菌株25 h的亚硝酸盐消除率达到(91.33±1.27)%,显著高于其他处理;29 h时的测定结果显示,该条件下亚硝酸盐消除率已经达到(94.46±2.75)%,而光照有氧条件下的亚硝酸盐消除率仅为(45.50±2.80)%,2种黑暗条件的亚硝酸盐消除率分别为(8.34±2.89)%和(12.18±1.96)%;42 h时,光照厌氧条件下的亚硝酸盐消除率为100%,而光照有氧条件下的亚硝酸盐消除率也达(91.32±2.69)%,黑暗条件的消除率则分别仅为(21.55±15.28)%和(45.22±2.00)%。
![]() 图 2 光照和氧气的不同组合对2-8菌株亚硝酸盐消除能力的影响Fig. 2 Influence of different illumination-oxygen combinations on nitrite removal effect of strain 2-8
图 2 光照和氧气的不同组合对2-8菌株亚硝酸盐消除能力的影响Fig. 2 Influence of different illumination-oxygen combinations on nitrite removal effect of strain 2-82.2 pH对菌株生长和亚硝酸盐消除的影响
2.2.1 pH对2-8菌株生长的影响
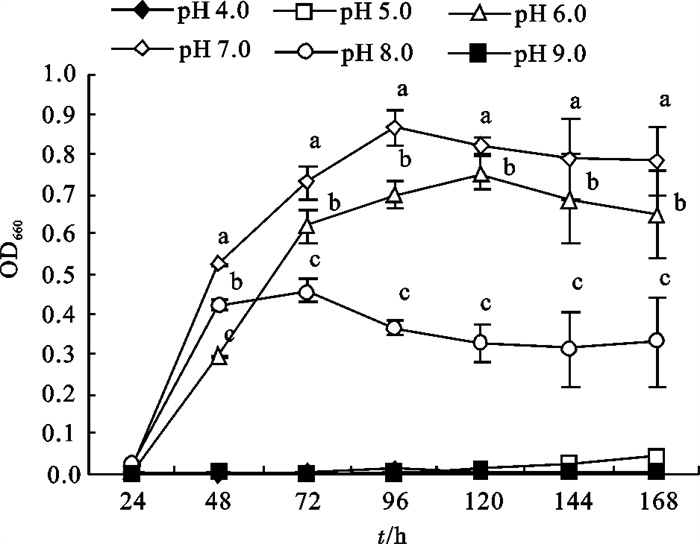
2-8菌株在pH 7.0的环境条件下生长良好,其最高OD660为0.87±0.04,其次是pH 6.0,偏酸偏碱都不利于2-8菌株的生长,pH为4.0、5.0和9.0时,2-8菌株几乎不能生长(图 3)。
2.2.2 pH对2-8菌株亚硝酸盐消除能力的影响
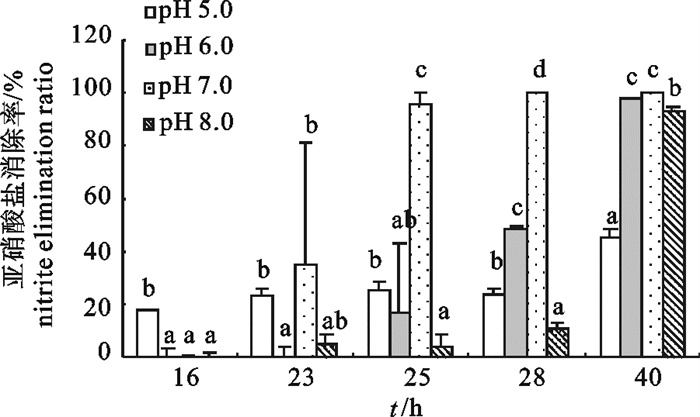
生测体系的pH对2-8菌株亚硝酸盐消除能力影响显著,偏酸和偏碱环境不利于菌株的亚硝酸盐消除(图 4)。生测体系pH为4.0和9.0时,菌株基本不利用亚硝酸盐;生测体系pH为5.0时,菌株可以消除亚硝酸盐,但消除能力受到严重影响,到40 h时,亚硝酸盐的消除率仍然只有(44.87±3.16)%,显著低于其他处理;菌株在pH 7.0的环境条件下亚硝酸盐消除能力最强,在第25小时,体系的亚硝酸盐消除率已经达到(95.58±4.34)%,显著高于其他处理;pH 6.0和8.0对2-8菌株的亚硝酸盐消除有一定的影响,主要是使体系亚硝酸盐的消除时间延长,第28小时pH 6.0条件下菌株的亚硝酸盐消除率为(48.25±1.58)%,pH 8.0条件下菌株的亚硝酸盐消除率为(10.67±2.03)%,而此时pH 7.0条件下菌株的亚硝酸盐消除率达到了100%;到第40小时,这2个处理的消除率分别达到(97.52±0.59)%和(93.45±1.12)%。
![]() 图 4 pH对2-8菌株亚硝酸盐消除能力的影响Fig. 4 Influence of different pH levels on nitrite removal effect of strain 2-8
图 4 pH对2-8菌株亚硝酸盐消除能力的影响Fig. 4 Influence of different pH levels on nitrite removal effect of strain 2-82.3 盐度对菌株生长和亚硝酸盐消除的影响
2.3.1 w (NaCl)对2-8菌株生长的影响
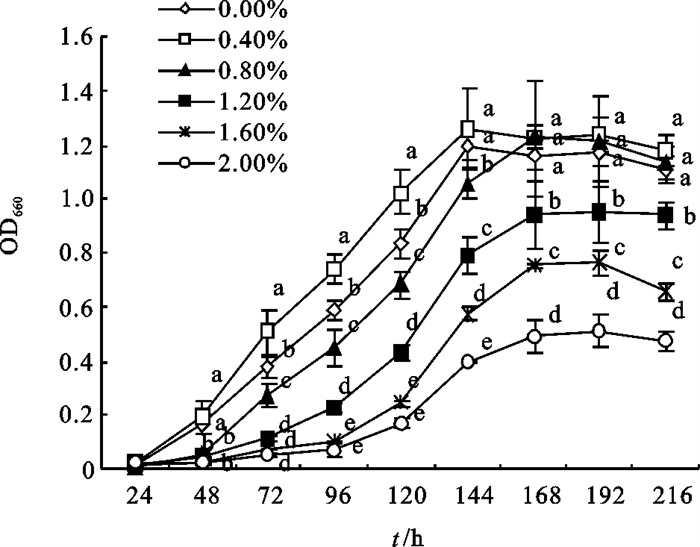
2-8菌株在w (NaCl)为0%~0.80%范围内生长良好且在0.40%生长最好,其最高OD660为1.26±0.14,随着w (NaCl)的升高,2-8菌株生长状况直线下降,在w (NaCl)为2.00%时,其最高OD660仅为0.51±0.065(图 5)。
![]() 图 5 w (NaCl)对2-8菌株生长的影响Fig. 5 Influence of different NaCl concentrations on growth of strain 2-8
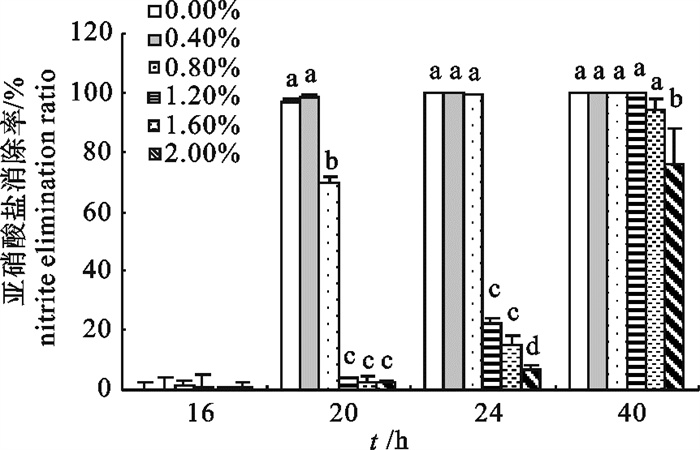
图 5 w (NaCl)对2-8菌株生长的影响Fig. 5 Influence of different NaCl concentrations on growth of strain 2-82.3.2 w (NaCl)对2-8菌株亚硝酸盐消除能力的影响
用w (NaCl)模拟的盐度水平测试显示,虽然菌株在所测定的盐度范围内最后都能将体系的亚硝酸盐消除,但高盐度的环境延缓了消除效果的发挥(图 6)。w (NaCl)在0%~0.8%的水平下,菌株在第24小时可以将体系中亚硝酸盐消除98%以上,而此时w (NaCl)1.2%~2.0%处理的消除率仅为6.93%~21.99%,到第40小时,w (NaCl)为1.2%、1.6%和2.0%处理的亚硝酸盐消除率分别为100%、(94.28±3.30)%和(75.80±12.37)%。
![]() 图 6 w (NaCl)对2-8菌株亚硝酸盐消除能力的影响Fig. 6 Influence of different NaCl concentrations on nitrite removal effect of strain 2-8
图 6 w (NaCl)对2-8菌株亚硝酸盐消除能力的影响Fig. 6 Influence of different NaCl concentrations on nitrite removal effect of strain 2-83. 讨论
亚硝酸盐污染是凡纳滨对虾养殖生产实践中仅次于病害的第二大制约因子,常与病害同时出现,严重影响了对虾养殖的健康发展。光合细菌在水产养殖中被广泛用于水质净化,发挥有益菌和生物饵料的作用[13]。文章研究的2-8菌株在光照厌氧环境中生长最好,但在光照有氧时的生长和对亚硝酸盐消除能力也较强,适合在对虾养殖水体中生长繁殖和应用;pH中性或偏酸时菌株的生长最好,与倪黎纲等[23]报道的相似;pH 7.0时亚硝酸盐消除功能最好,与张李阳和吴向华[20]的结果类似;但菌株在pH 8.0时40 h亚硝酸盐的消除率也可以达到(93.45±1.12)%,能够满足生产实践的需要,可见2-8菌株的生长和功能发挥的pH水平与养殖水体的实际pH水平能够吻合;2-8菌株在低盐度的水体中繁殖能力较强,盐度超过12,繁殖能力明显受到抑制,亚硝酸盐的消除能力在24 h内受到盐度的影响较大,并有随盐度增加而所受影响增大的现象,显示菌株较适应珠三角的池塘养殖,尤其是养殖后期,水体盐度在0~4之间波动,与菌株生长和亚硝酸盐功能发挥的最佳盐度水平非常吻合。
-
![]()
图 1 光照和氧气组合对2-8菌株生长的影响
注:同一时间不同处理上的小写字母表示处理间SPSS方差分析和LSD多重比较差异显著(P<0.05),后图同此
Figure 1. Influence of different illumination-oxygen combinations on growth of strain 2-8
Note:Different lowercase letters on the same time column indicate different treatments influence the strain′s growth significantly (P<0.05, LSD, SPSS 13.0). The same case in the following figures.
![]()
图 2 光照和氧气的不同组合对2-8菌株亚硝酸盐消除能力的影响
Figure 2. Influence of different illumination-oxygen combinations on nitrite removal effect of strain 2-8
![]()
图 3 不同pH水平对2-8菌株生长的影响
Figure 3. Influence of different pH levels on growth of strain 2-8
![]()
图 4 pH对2-8菌株亚硝酸盐消除能力的影响
Figure 4. Influence of different pH levels on nitrite removal effect of strain 2-8
![]()
图 5 w (NaCl)对2-8菌株生长的影响
Figure 5. Influence of different NaCl concentrations on growth of strain 2-8
-
[1] 徐培峰,曾宪凯. 南美白对虾淡水养殖中亚硝酸盐危害分析与对策[J]. 渔业现代化,2006(2):38-39. doi: 10.3969/j.issn.1007-9580.2006.02.015 [2] 高明辉, 马立保, 葛立安, 等. 亚硝酸盐在水生动物体内的吸收机制及蓄积的影响因素[J]. 南方水产, 2008, 4(4): 73-79. doi: 10.3969/j.issn.2095-0780.2008.04.014 [3] JEBSEB F B. Nitrite disrupts multiple physiological functions in aquatic animals[J]. Comp Biochem Physiol A: Mol Integr Physiol, 2003, 135(1): 9-24. doi: 10.1016/S1095-6433(02)00323-9
[4] 黄翔鹄, 曾宪凯, 郑莲, 等. 亚硝酸盐氮对凡纳滨对虾毒性和抗病相关因子影响[J]. 水生生物学报, 2006, 30(4): 466-471. doi: 10.3321/j.issn:1000-3207.2006.04.016 [5] 王安利, 郑桂丽, 廖绍安, 等. 虾池中具有降解硝酸盐或亚硝酸盐能力的细菌多样性[J]. 生态学报, 2007, 27(5): 1937-1944. doi: 10.3321/j.issn:1000-0933.2007.05.033 [6] TABITA F R, HANSON T E. An oxygenic phototrophic bacteria[M]//FRASER C M, READ T D, NELSON K E. Microbial genomes. Totowa, New Jersey, USA: Humana Press, 2004: 225.
[7] LARIMER F W, CHAIN P, HAUSER L, et al. Complete genome sequence of the metabolically versatile photosynthetic bacterium Rhodopseudomonas palustris[J]. Nat Biotechnol, 2004, 22(1): 55-61. doi: 10.1038/nbt923
[8] 刘芳, 王敏, 杨慧, 等. 净化养殖水体紫色非硫光合细菌的筛选与鉴定[J]. 中国生物工程杂志, 2008, 28(8): 91-95. doi: 10.3969/j.issn.1671-8135.2008.08.017 [9] 刘如林, 刁虎欣, 梁凤来, 等. 光合细菌及其应用[M]. 北京: 中国农业科技出版社, 1991. [10] 杨启银, 曩潇潇, 何瑶, 等. 富硒沼泽红假单胞菌对D-半乳糖小鼠肝细胞修复作用的研究[J]. 食品科学, 2007, 28(10): 497-501. doi: 10.3321/j.issn:1002-6630.2007.10.126 [11] 岳文洁, 刘春, 张小凡. 沼泽红假单胞菌累积聚β-羟基丁酸的研究[J]. 环境科学与技术, 2007, 30(4): 26-28. doi: 10.3969/j.issn.1003-6504.2007.04.010 [12] 王永忠, 廖强, 朱恂, 等. 序批式培养沼泽红假单胞菌光照产氢的能量分析[J]. 太阳能学报, 2009, 30(3): 390-396. doi: 10.3321/j.issn:0254-0096.2009.03.022 [13] 刘朝阳, 孙晓庆. 生物控制法在水产养殖水质净化中的综合应用[J]. 南方水产, 2007, 3(1): 69-74. doi: 10.3969/j.issn.2095-0780.2007.01.012 [14] QI Zizhong, ZHANG Xiaohua, BOON N, et al. Probiotics in aquaculture of China-current state, problems and prospect[J]. Aquac, 2009, 290(1/2): 15-21. doi: 10.1016/j.aquaculture.2009.02.012
[15] 黄宁宇, 来琦芳. 三种光合细菌不同生态环境下的生长试验[J]. 海洋渔业, 2004, 26(3): 194-204. [16] 徐姗楠, 邱宏端, 林娟, 等. 紫色非硫光合细菌的分离鉴定及其功能研究[J]. 福州大学学报: 自然科学版, 2004, 32(2): 246-251. doi: 10.3969/j.issn.1000-2243.2004.02.027 [17] 周茂洪, 赵肖为, 周峙苗. 几种重金属离子对光合细菌生长的抑制效应[J]. 生态学杂志, 2002, 21(4): 6-11. https://d.wanfangdata.com.cn/periodical/stxzz200204002 [18] 周茂洪, 王荟沪, 岑宇. Hg2+、Pb2+、Cd2+和Cu2+对光合细菌沼泽红单胞菌的生长及净化氮、磷能力的影响[J]. 生态科学, 2002, 21(1): 55-58. [19] 蔡慧农, 倪辉, 苏文金. 沼泽红假单胞菌培养基的优化及降氨氮作用的研究[J]. 集美大学学报: 自然科学版, 2007, 12(3): 199-203. doi: 10.3969/j.issn.1007-7405.2007.03.002 [20] 张李阳, 吴向华. 不同条件光合细菌同化磷能力的初步探究[J]. 南京晓庄学院学报, 2004, 20(4): 1-4. doi: 10.3969/j.issn.1009-7902.2004.04.001 [21] 国家环保局《水和废水监测分析方法》编委会. 水和废水监测分析方法[M]. 3版. 北京: 中国环境科学出版社, 1989: 260-263. [22] 喻国辉, 陈远凤, 陈燕红, 等. 一株高效利用亚硝态氮的沼泽红假单胞菌及其应用: 中国, ZL200710089906.5[P]. 2007-10-10. https://www.zhangqiaokeyan.com/patent-detail/06120105249595.html [23] 倪黎纲, 王志鹏, 罗方妮, 等. 氧、光照时间、pH等理化因子影响光合细菌生长的研究[J]. 内陆水产, 2004(3): 41-42. doi: 10.3969/j.issn.1674-9049.2004.03.03 -
期刊类型引用(20)
1. 郭泽裕,刘兴国,陈哲,袁泽辉,陆诗敏. 基于Ecopath模型的渔光互补养殖池塘比较分析. 水产科学. 2025(03): 385-393 .  百度学术
百度学术
2. 刘明. 基于光合细菌红假单胞菌的废水处理与协同固碳作用研究. 生物质化学工程. 2024(02): 47-54 . 百度学术
3. 鲁晏宏,王蕊,王珺,胡婷,付维来,朱传忠,汪攀. 1株耐盐耐低温沼泽红假单胞菌的分离筛选及培养基优化. 饲料研究. 2023(20): 103-108 . 百度学术
4. 信艳杰,胡晓娟,曹煜成,徐煜,许云娜,苏浩昌,徐创文,文国樑,李卓佳. 光合细菌菌剂和沼泽红假单胞菌对实验水体氮磷营养盐和微生物群落的影响. 南方水产科学. 2019(01): 31-41 . 本站查看
5. 王蓉,龚世飞,金涛,刘章勇. 稻虾共作对稻田土壤反硝化细菌nosZ基因数量、多样性及群落结构的影响. 江苏农业科学. 2019(04): 246-251 . 百度学术
6. 张晓波,周广静,朱笔通,赵春贵,杨素萍. 光氧环境对紫色硫细菌YL28去除无机三态氮的影响. 微生物学通报. 2019(05): 997-1007 . 百度学术
7. 丁海涛,曹开银,钟伦锁,陈俊,金杰. 光合细菌处理黑臭水体影响因素的研究. 合肥学院学报(综合版). 2019(05): 51-56 . 百度学术
8. 文刚,汪彬,刘标,贺月林. 沼泽红假单胞菌R-3去除水体中氨氮的特性研究. 湖南农业科学. 2017(06): 49-51 . 百度学术
9. 陈文斌,潘鲁青,黄飞. 对虾生物絮团中亚硝态氮降解菌分离鉴定与安全性评价. 海洋湖沼通报. 2017(03): 121-129 . 百度学术
10. 易馨,杨开智,童晋,李明学,谢鸿观. 响应面法对光合细菌还原亚碲酸盐条件的优化. 江苏农业科学. 2017(10): 217-220 . 百度学术
11. 张晓波,朱笔通,产竹华,赵春贵,曾润颖,杨素萍. 有机碳对海洋着色菌YL28去除无机三态氮的影响. 微生物学通报. 2017(05): 1017-1027 . 百度学术
12. 周梦舟,吴珊,柳念,汪超,徐宁,胡勇,史文博,李冬生. 基于神经网络对枯草芽孢杆菌富硒过程的建模研究. 中国食品学报. 2016(12): 66-74 . 百度学术
13. 张晓波,朱笔通,熊慧,赵春贵,杨素萍. 沼泽红假单胞菌(Rhodopseudomonas palustris)CQV97对无机三态氮共存水体中氮素的去除效率及其影响因素. 氨基酸和生物资源. 2015(04): 38-45 . 百度学术
14. 蒋鹏,洪璇,赵春贵,杨素萍. 海洋着色菌Marichromatium gracile YL28静息细胞对无机三态氮的相互转化作用. 华侨大学学报(自然科学版). 2015(02): 185-189 . 百度学术
15. 王志杰,胡修贵,刘旭雅,宋晓玲,马甡,黄倢. 自生物絮团养殖池分离具有亚硝酸盐去除功能的细菌及其鉴定和特性. 渔业科学进展. 2015(02): 100-105 . 百度学术
16. 王雅楠,吴燕燕,任中阳,李来好,杨贤庆,周婉君. 亚硝酸盐还原酶对咸鱼中亚硝酸盐的降解条件与效果研究. 食品工业科技. 2015(08): 210-214+229 . 百度学术
17. 蒋鹏,赵春贵,杨素萍. 小分子有机碳、氮源对海洋着色菌(Marichromatium gracile)生长和去除高浓度无机三态氮的影响. 海洋与湖沼. 2014(06): 1218-1224 . 百度学术
18. 赵玥,赵春贵,陈龑,杨素萍. 沼泽红假单胞菌CQV97菌株对污染水体三氮去除特性研究. 山西大学学报(自然科学版). 2012(03): 557-562 . 百度学术
19. 纪敦敦,邱宏端,谢航. 采用原生质体融合技术选育提高氨氮降解效能的光合细菌. 大连海洋大学学报. 2011(05): 407-413 . 百度学术
20. 喻国辉,陈燕红,程萍,黎永坚,杨紫红,陈远凤. 几种金属离子对沼泽红假单胞菌2-8生长和亚硝酸盐消除的影响. 南方水产科学. 2011(04): 30-35 . 本站查看
其他类型引用(13)
 下载:
下载:
计量
- 文章访问数: 5194
- HTML全文浏览量: 233
- PDF下载量: 3123
- 被引次数: 33
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
