
Effects of salinity on embryonic development and early larvae in ovate pompano Trachinotus ovatus
-
摘要:
观察比较了10种不同盐度(5.0~50.0)条件下卵形鲳鲹(Trachinotus ovatus)受精卵的沉浮性、孵化率和畸形率,以及8种不同盐度(10.0~45.0)下仔鱼存活情况及不投饵存活系数(survival activity index,SAI)。结果表明,受精卵在不同盐度下表现出不同的沉浮性,在盐度为25.0以下的海水中呈沉性;在盐度30.0时为半沉浮性;在盐度为35.0的海水中呈浮性。盐度低于10.0或高于50.0,胚胎均不发育;在盐度15.0~45.0范围内,卵形鲳鲹胚胎能完成发育过程孵出仔鱼,但盐度对早期仔鱼的成活和畸形率的影响比较大。统计分析了总孵化率与盐度、畸形率与盐度的相关关系,求得胚胎孵化的预测适盐范围为14.9~39.4,最适盐度为26.0~28.2。在盐度25.0~30.0和无投饵状态下,仔鱼的SAI值为44.19~47.44。
Abstract:The effects of salinity on the buoyancy, hatching and abnormal rate of fertilized eggs of ovate pompano (Trachinotus ovatus) at 10 different salinities ranging from 5.0 to 50.0 were studie. Moreover, the survivorship as well as survival activity index (SAI) of larvae T.ovatus at 8 different salinities ranging from 10.0 to 45.0 were also observed. The results showed that different buoyancies of fertilized eggs were exhibited under different salinity conditions: all eggs sank at salinity bellow 25.0; most of eggs suspended in the middle of water at 30.0;all eggs floated on the surface at salinity above 35.0. Eggs did not hatch out at salinity below 10.0 or above 50.0, and the salinities for their hatching out ranged from 15.0 to 45.0. However, the effects of salinity on the survival rate and abnormal rate of early larval were significant. The relationship between total hatching rate and salinity variation as well as that between abnormal rate and salinity was analyzed statistically. Expected suitable salinity for embryonic development was estimated as 14.9~39.4 and the optimal salinity was 26.0~28.2. SAI value of early larvae was calculated as 4.19~47.44 under non-feeding condition and at salinities ranging from 25.0 to 30.0.
-
Keywords:
- ovate pompano (Trachinotus ovatus) /
- salinity /
- embryonic development /
- larvae
-
钝缀锦蛤 (Tapes dorsatus) 俗称“沙包螺”,为软体动物门、瓣鳃纲、帘蛤目、帘蛤科、缀锦蛤属的海洋贝类动物,多生活在潮间带下区和潮下浅水区的泥砂底,广泛分布于西太平洋和印度洋水域。其个体大,足部与软体部肌肉发达,肉质鲜美,是沿海地区居民经常食用的水产品之一,具有极高的经济价值[1]。迄今为止,国内外关于钝缀锦蛤的研究报道主要集中于苗种孵化生产[2]、性状关联及遗传力分析[3-4]、生态习性 (如温度、盐度、氨氮及pH等生态环境因子对其生长和存活的影响)[5-9]、繁殖生物学 (包括繁殖习性和胚胎、幼虫、稚贝的生长发育过程等) [10-12]等方面。
目前,钝缀锦蛤几乎全部为天然野生资源,由于近年来海区环境恶化和大肆捕捞,导致其自然资源遭到严重破坏。因此,开展钝缀锦蛤人工育苗技术研究,对实现种苗规模化生产具有重要意义。迄今有关钝缀锦蛤苗种培育的报道仅见于曾志南等[13]的苗种繁育技术和彭银辉等[14]的人工育苗方法研究。本研究在曾志南等[13]的研究基础上进行了改进,在亲贝选择、促熟、催产以及稚贝培育、采苗方式上进行了创新。此外,本研究尝试在冬季进行反季节育苗,以解决钝缀锦蛤反季节人工育苗中亲本性腺不成熟、常规育苗单位面积出苗量低、种苗生长慢、耗水量大等问题,弥补冬季市场空缺。通过开展钝缀锦蛤苗种繁育关键技术研究,可为其大规模人工育苗提供参考,并克服规模化养殖面临的种苗供应问题,促进产业发展。
1. 材料与方法
1.1 材料
实验于2020年10月在广东省雷州市覃斗镇贝类苗种培育基地进行,11月15日从广西防城港市购进越南同批人工种苗同一地点养成的钝缀锦蛤群体,其中随机购买100 kg未经选择的个体,作为非选育繁育群体;再按5%留种率上选最大体质量个体300 kg,选择强度为2.06[15],作为选育繁育群体,用于群体选育研究。在两个繁育群体中各随机测量50个个体的性状参数 (表1)。
表 1 钝缀锦蛤两个发育群体性状参数Table 1. Traits parameters of two T. dorsatus developmental populations性状参数
Traits parameter壳长
Shell length/mm壳高
Shell height/mm壳宽
Shell width/mm体质量
Body mass/g非选育繁育群体
Non-selective breeding population64.9±3.8 (5.86) 43.6±2.5 (5.73) 26.3±0.22 (8.37) 31.6±4.8 (15.19) 选育繁育群体
Selective breeding population70.9±1.7 (2.52) 47.6±1.2 (2.52) 28.7±0.7 (2.44) 38.8±3.3 (8.51) 性状选择差
Selection differential of traits (S)6.0 4.0 2.4 7.2 注:括号里的数值表示繁育群体各性状的变异系数 (%)。 Note: The values in parentheses are the variable coefficients (%) of each trait of breeding population. 1.2 实验设计
本研究钝缀锦蛤人工育苗包括选育、促熟、促产、封闭育苗、采苗和稚贝培养共6个关键技术环节,每个环节都在前期育苗实践经验上进行了创新,并将分散独立的技术集成贯穿到整个育苗生产中。同时,为探究每个技术在实际生产中的效果,设置了相应的对照组进行验证,其中促熟和促产两个技术可通过促熟率与促产率直接验证;选育、封闭育苗、采苗3个技术均通过对比最终的育苗效果进行验证,实验方案设计如下:
1) 选育:在同批次、同一地点养成的钝缀锦蛤繁育基础群中,按5%留种率上选最大体质量个体建立选育繁育群体,作为实验组,同时将剩余未经选育的个体建立非选育繁育群体,作为对照组1。保证其他因素不变,每组设3个平行进行后续实验。
2) 促熟:将钝缀锦蛤亲贝吊挂于室内10 m3水泥池,吊养密度为150 个·m−3,每池约1 500个,水温从常温逐步升至26 ℃恒温,每天等温换水1/2。促熟期间每日定时投饵,投饵量视水色变化加以调节,保证亲本充足摄食。为比较不同饵料对促熟效果的影响,设置不同的实验组别,其中实验组在促熟期间以虾塘藻为饵料,虾塘藻按对虾养殖的常规方法进行培育,包含多种优质复合藻类,如角毛藻 (Chaetoceros sp.)、菱形藻 (Nitzschia sp.)、卵形藻 (Cocconeis sp.)、舟形藻 (Navicula sp.) 等,为亲本促熟提供大量营养互补的饵料,以满足其性腺发育的营养需求。对照组促熟期间仅投喂浓缩拟微球藻 (Nannochloropsis gaditana),保证其他因素不变,每组设3个平行。
3) 促产:为提高促产率,比较温度对促产效果的影响,设立了实验组与对照组。实验组采用升温阴干悬浮静置方法进行促产,将亲贝从26 ℃水温促熟池中取出,移入室内30 ℃恒温阴干6~8 h,再移入26 ℃水温催产池中的悬浮塑料筛里,停止充气,悬浮静置待产。待大部分个体开始产精排卵,即开始微波状充气,通过水微循环分散卵子,当卵子密度达到3~4粒·mL−1时,及时将亲贝移入其他催产池继续排精产卵。对照组采用“室内自然温度 (22~26 ℃) 阴干悬浮静置促产”方式进行催产,亲贝未经升温,在室内自然温度下阴干,其他条件与实验组相同,每组设3个平行。
4) 封闭育苗:为避免频繁换水导致育苗失败,本实验采用不换水的封闭育苗法,育苗期间水温控制在24~26 ℃。为保证封闭育苗期间的水质,通过添加微生物制剂的方式提高分解有机物的效率,消除腐败,抑制病原菌,形成适于动植物生长的良好环境。为验证微生物制剂在封闭育苗中的作用,分3个不同的组别进行实验,实验组在封闭育苗过程中添加EM菌 (Effective microorganisms,又名“益生菌”),另设2个对照组,分别为对照组2和3;对照组2不添加微生物制剂,对照组3添加光合细菌。此外,3组每天均加入适量的维生素B、维生素C和葡萄糖,以促进水体微生态系统稳定,保持其他因素不变,每组设3个平行。
5) 采苗:为提高采苗效率,本实验采用“立体采苗器+池底”采苗模式,每串采苗器长度110 cm,由12层聚乙烯塑料板构成,层距10 cm,板规格为30 cm×30 cm,每串面积为1.08 m2,投放密度为3串·m−2,30 m2的池子投放90串,采苗器总采苗面积为97.2 m2·池−1;池底采苗面积为30 m2,采苗器采苗面积是池底的3.24倍,整池采苗面积为127.2 m2。采苗前,采苗器经过消毒清洗,池底通过虹吸法刮净脏物,提高池底采苗效果。当幼虫70%发育至壳长220 μm的成熟期时,即投放采苗器。为验证立体采苗的效率,设置对照组4,不投放采苗器,仅采用池底采苗法进行采苗,保证其他因素不变,每组设3个平行。
6) 稚贝培养:幼虫附着后会变态成为稚贝,附着7 d后,稚贝长大,抗逆性增强,开始投喂虾塘复合藻。由于投喂虾塘藻时带来大量虾塘水,水体处于对流状态,因此不再使用EM菌。为使稚贝出池后适宜冬季的低温,育苗池水温在附着后第11至第20天降至24 ℃,第21至第30天降至22 ℃,30 d后停止加温,让池内水温逐渐接近自然水温。附着后经过30~40 d的培育,稚贝壳长达到2 mm左右即可出苗,育苗工作完成。
1.3 数据处理
1) 性状选择差S=选择亲本性状−非选择亲本性状;
2) 性腺促熟率= (性腺完全成熟个数/促熟个数)×100%,抽测样本数量100个;
3) 催产率=解剖法检查产卵排精总个数/(用于催产总数×性腺促熟率)×100%,抽测样本数量100个;
4) 孵化率= (D形幼虫数/受精卵数)×100%;
5) 浮游幼虫期成活率=(投放采苗器前壳顶后期幼虫数量/D形幼虫数量)×100%;
6) 稚贝育成率=(稚贝出苗量/D形幼虫数量)×100%;
7) 稚贝育成提高率=(立体采苗稚贝育成率−平面采苗稚贝育成率)/平面采苗稚贝育成率×100%;
8) 选育稚贝性状提高率=(选育稚贝性状−非选育稚贝性状)/非选育稚贝性状×100%;
9) RL=
$\dfrac{{L}_{1}-{L}_{0}}{t}$ ;10) RH=
$\dfrac{{H}_{1}-{H}_{0}}{t};$ 式中:RL为壳长生长率 (μm·d−1);RH为壳高生长率 (μm·d−1);L0为各个发育阶段起始时壳长 (μm);L1为各个发育阶段结束时壳长 (μm);H0为各个发育阶段起始时壳高 (μm);H1为各个发育阶段结束时壳高 (μm);t为各个发育阶段的发育时间 (d);幼虫和稚贝的壳长、壳高用目微尺在10×10镜头下测量,每次测量30粒随机个体。
11) 选择反应:SR=
$ \dfrac{\mathrm{B}\mathrm{S}-\mathrm{B}\mathrm{C}}{{\sigma }_{\mathrm{B}\mathrm{C}}} $ ;12) 现实遗传力:h2=
$\dfrac{\mathrm{B}\mathrm{S}-\mathrm{B}\mathrm{C}}{{i\sigma }_{\mathrm{B}\mathrm{C}}}$ ;由于贝类的生长受环境影响很大,其个体大小会随环境而变化,故参考Zheng等[16]的方法计算不同生长期的选择反应和现实遗传力,消除环境因素对遗传力的影响[17]。式中:BS为选育后稚贝的平均壳长;BC为对照组稚贝的平均壳长;σBC为对照组的标准差;i为选择强度。
实验数据采用“平均值±标准差 (
$\overline { X}\pm { \rm {SD}} $ )”表示,使用SPSS 26.0软件对数据进行单因素方差分析 (One-way ANOVA),采用Duncan's多重比较法来检验处理组间的差异显著性,用Origin 2021软件作图。2. 结果
2.1 不同促熟方式对促熟效果的影响
采用不同的方式进行促熟,实验组投喂虾塘复合藻,对照组投喂浓缩拟微球藻,同时促熟19 d后,实验组亲本的性腺成熟率 [(88.0±2.9)%] 显著高于对照组 [(23.0±1.5)%, P<0.05],前者比后者提高了282.6%。
2.2 催产方式对催产率的影响
采用不同的方式对亲贝进行催产,实验组亲本开始排放精卵的时间为 (62±3) min,对照组亲本为 (78±2) min,显著慢于实验组 (P<0.05),前者比后者时间缩短了20.5%。同时实验组的催产率 [(83.6±2.8)%] 显著高于对照组 [(55.4±2.4)%, P<0.05],比对照组提高了50.9%。
2.3 微生物制剂对育苗效果的影响
采用EM菌的实验组,浮游期成活率、幼虫生长速度、稚贝育成率、出池稚贝壳长均显著大于不采用微生物制剂的对照组2和使用光合细菌的对照组3 (P<0.05,表2);而使用光合细菌的对照组3,浮游期成活率、幼虫生长速度、稚贝育成率、出池稚贝壳长均显著大于不采用微生物制剂的对照组2。
表 2 封闭式育苗采用不同微生物制剂的育苗效果对比Table 2. Effects of different microbial agents on closed seedling breeding组别
Group浮游期成活率
Survival rate in
pelagic period/%幼虫生长速度
Larval growth
rate/(μm·d−1)稚贝育成率
Juvenile rearing
rate/%出池稚贝壳长
Juvenile shell of length
out-of-pool spats/mm实验组 Test group 89.3±2.91a 9.33±0.25a 25.60±1.18a 1.31±0.026a 对照组2 Control group 2 38.6±1.35c (131.3) 7.45±0.21c (25.2) 7.86±0.35c (225.7) 1.03±0.033b (27.2) 对照组3 Control group 3 67.1±2.02b (33.1) 8.05±0.22b (15.9) 10.52±0.42b(143.3) 1.09±0.030b (20.2) 注:括号里的数值表示实验组相对于对照组的提高率 (%);同列中不同字母表示存在显著性差异 (P<0.05)。 Note: The values in parentheses are the improvement rates (%) of the test group compared to the control group. Values with different letters within the same column indicate significant differences (P<0.05). 2.4 立体采苗方式对育苗效果的影响
实验组采用“立体采苗器+池底”采苗模式,对照组4仅采用池底采苗的方法。结果表明:1) 实验组立体采苗器和池底的采苗密度均小于对照组;2) 用立体采苗器进行采苗的采苗量显著大于池底采苗量,实验组的采苗量比对照组提高了76.5%;3) 实验组两种采苗方式附着的苗种生长速度均大于对照组,实验组的稚贝平均壳长比对照组提高了21.9%;4) 实验组大部分稚贝附着于采苗器上,减轻了池底稚贝的生存压力,稚贝育成率达到25.6%,而对照组4池底附着密度高、死亡率高,稚贝育成率仅14.5%,前者比后者提高了76.6% (表3)。
表 3 不同采苗方式的育苗效果对比Table 3. Seedling breeding effects of different seedling collection methods指标
Index采苗方式
Seedling collection method实验组
Test group对照组4
Control group 4采苗密度
Seedling density/(万粒·m−2)采苗器
池底18.00±0.81(b)
23.58±1.13a(a)
46.40±1.51b (−49.2)采苗量
Seedling quantity/(万粒·池−1)采苗器
池底
合计1 749.6±78.6(a)
707.4±33.9b(b)
2 457.0±75.6a
1 392.0±45.3b (−49.2)
1 392.0±45.3b (76.5)稚贝壳长
Juvenile shell length/mm采苗器
池底
加权平均1.33±0.09(a)
1.16±0.08a(b)
1.28
1.05±0.03b (10.5)
1.05±0.03 (21.9)稚贝育成率
Juvenile rearing rate/%25.6±1.18a 14.5±0.61b (76.6) 注:上标不同小写字母表示各组之间差异显著 (P<0.05),其中不带括号的为组间分析,带括号的为组内分析;对照组的相应数据后括号内的数值表示实验组相对于对照组的提高率 (%)。 Note: Different lowercase letters indicate significant differences among the groups (P<0.05). Those without parentheses are between-group analyses, and those with parentheses are within-group analyses. The values in parentheses after the control group indicate the improvement rate (%) of the test group compared to the control group. 2.5 选育与否对育苗效果的影响
由表4可见,选育组培育出的稚贝壳长比未选育组提高了6.67%,体质量提高了17.03%,稚贝育成率提高了8.43%,壳长和体质量的变异系数分别降低了29.13%和55.29%,现实遗传力分别达到0.65和0.72。
表 4 选育与不选育的育苗效果对比Table 4. Comparison of seedling breeding effects between selection breeding and non-selection breeding组别
Group壳长
Shell
length/mm体质量
Body
mass/mg壳长变异系数
Variable coefficient of
shell length/%体质量变异系数
Variable coefficient of
body mass/%稚贝育成率
Juvenile rearing
rate/%实验组稚贝
Test group juvenile1.28±0.03a 2.13±0.11a 7.81 5.16 25.60±1.18a 对照组1稚贝
Control group 1 juvenile1.20±0.06b (6.67) 1.82±0.21b (17.03) 11.02 (−29.13) 11.54 (−55.29) 23.61±1.12a (8.43) 选择反应
Selective reaction1.33 1.48 现实遗传力 h2 0.65 0.72 注:括号里的数值表示实验组相对于对照组的提高率 (%);同列中不同字母表示存在显著性差异 (P<0.05)。 Note: The values in parentheses are the improvement rates (%) of the test group compared to the control group. Values with different letters within the same column indicate significant differences (P<0.05). 2.6 技术集成下幼虫及稚贝生长规律
通过对技术集成方法下培育的钝缀锦蛤胚胎、幼虫及稚贝发育阶段进行连续观察,发现钝缀锦蛤早期生长发育速度呈现“快-慢-快”的变化趋势 (表5)。发育各个时期的生长情况如图1所示,最慢的阶段在由变态期幼虫发育到无管期稚贝期间,为期3 d,壳长和壳高增长率分别为2.03和1.40 μm·d−1,其他阶段的壳长和壳高增长率随个体发育不断增加。
表 5 钝缀锦蛤幼虫期至稚贝期生长率Table 5. Growth rate in larval and juvenile stages of T. dorsatus发育阶段
Developmental stage发育时间
Development time/d壳长生长率
Shell length growth
rate/(μm·d−1)壳高生长率
Shell height growth
rate/(μm·d−1)D形幼虫—壳顶初期幼虫
D-shape larvae−Early umbo larvae5.33 9.76 10.38 壳顶初期幼虫—壳顶中期幼虫
Early umbo larvae−Medium term umbo larvae3 10.63 11.23 壳顶中期幼虫—壳顶后期幼虫
Medium term umbo larvae−Post umbo larvae4 6.85 7.48 壳顶后期幼虫—变态期幼虫
Post umbo larvae−Metamorphosis larvae4 3.83 3.93 变态期幼虫—无管稚贝
Metamorphosis larvae−Non siphon juvenile3 2.03 1.40 无管稚贝—单管稚贝
Non siphon juvenile−Single siphon juvenile20 12.52 10.99 单管稚贝—双管稚贝
Single siphon juvenile−Double siphon juvenile10 9.33 9.05 双管稚贝—实验结束
Double siphon juvenile−End of experiment15 27.10 24.57 ![]() 图 1 钝缀锦蛤幼虫期至稚贝期生长曲线Figure 1. Growth curve of larval and juvenile stages of T. dorsatus
图 1 钝缀锦蛤幼虫期至稚贝期生长曲线Figure 1. Growth curve of larval and juvenile stages of T. dorsatus3. 讨论
3.1 钝缀锦蛤选择育种中目标性状的选择
与海洋贝类生长相关的性状如壳形态和质量性状等,被认为是选择育种中重要的经济性状[18-19]。在选育过程中,体质量是最直接、最能反映贝类生产性能的测量指标,但由于贝类的质量性状相对不易测量,目前主要采用相关性分析和通径分析来确定影响体质量的主要形态性状,因此可通过壳形态性状的选择达到选择育种的目的[20-22]。在以往关于钝缀锦蛤的研究中,巫旗生等[3]通过对钝缀锦蛤野生群体的形态性状和活体质量进行通径分析,明确了壳宽为影响其活体质量的主要性状,得出以壳宽作为首要选育目标的结论。不同月龄的贝,其大小不同,生长性状的相关性不同[23]。聂振平等[4]的研究表明,2龄钝缀锦蛤的壳宽对体质量直接作用最大,与前述报道一致。因此,钝缀锦蛤的选育可以考虑以壳宽为首,预计能取得较大的遗传进展。但本研究以体质量作为首选目标,主要基于下述考虑:1) 在追求体质量高产的生产体系中,育种目标首先是体质量,体质量可直接测量而无需用某一体尺替代。同时,体质量受多个体尺的综合影响,没有哪一个体尺可以100%影响体质量。因此,对体质量直接选育是最有效的。2) 体质量的变化受体尺的影响,因此体质量的变异系数往往大于其他性状。变异系数是进行品种选育的重要依据,其数值越大则说明选择潜力越大[3]。本研究中,钝缀锦蛤繁育基础群各性状变异系数分别为壳长5.86%、壳高5.73%、壳宽8.37%和体质量15.19%,对体质量进行选育,稚贝的壳长和体质量明显提高,现实遗传力h2分别达到0.65和0.72,因此直接以体质量为目标进行选育可获得较大的遗传进展。
3.2 投喂虾塘复合藻及升温对亲贝促熟效果的影响
亲贝的促熟技术关系到生殖腺的发育程度,直接影响最终的育苗效果,是虾夷扇贝 (Patinopecten yessoensis) 人工育苗的基础[24]。本研究表明,在室内水泥池人工控温条件下,采用室外虾塘复合藻作为饵料,其促熟效果显著好于采用纯种单胞藻。尤颖哲[25]的研究证实,用室外土池人工促熟法,波纹巴非蛤 (Paphiaun dulata) 亲贝性腺发育的成熟情况好、死亡率低,且孵化率得到有效提高。其原理在于贝类促熟需要两个基本要素,即积温和营养,只有两个要素都满足了才能完成性腺的成熟发育[26]。其中,温度是影响海洋生物生殖最重要的因素,合适的温度对贝类的性腺发育与繁殖有积极作用[27-28]。本研究采用室内逐渐升温至26 ℃恒温,能保证亲贝促熟期间获得有效积温,与巫旗生等[10]报道的钝缀锦蛤的性腺增殖最适温度为24~28 ℃的结论相一致。饵料是亲贝性腺发育的物质基础,饵料的种类和数量对亲贝的性腺发育起着至关重要的作用[29]。研究表明,亲贝促熟期的食性越杂,育苗效果越好[24,30]。传统的促熟方法采用纯种单胞藻,由于品种少,营养互补水平低,促熟效果差。张红云[31]研究发现在栉江姚 (Atrina pectinata) 亲贝繁育期间以混合藻为饲料促熟效果最好;虾夷扇贝[24]、波纹巴非蛤[25]、长牡蛎 (Crassostrea gigas) [32]及香港巨牡蛎 (C. hongkongensis)[33]促熟培育过程中均以多种藻类作为饵料。本研究采用的虾塘复合藻,包含各种浮游单胞藻,如角毛藻、菱形藻、卵形藻、舟形藻等硅藻和各种绿藻等,饵料生物密度高且品种多、质量好,营养互补效果好,极大地满足了亲贝性腺发育所需的营养,亲贝肥满快,性腺促熟率相比于投喂单一纯种单胞藻提高了269.7%。这与常亚青[34]认为饵料的质量和数量直接影响贝类繁殖的结论一致。
3.3 实施封闭式育苗的关键措施
EM菌是一种新型复合微生物活性菌剂[35],由5大类微生物——光合菌群、酵母菌群、乳酸菌群、放线菌群及发酵系的丝状菌群组成。其主要功能是高效分解有机物、消除腐败和抑制病原菌,形成适于动植物生长的良好环境。在水产养殖中,EM菌对改善水质、提高成活率和相对生长率及增强水产动物免疫力有重要作用[36]。在养殖过程中添加EM菌可起到净化水质的作用[37];在凡纳滨对虾 (Litopenaeus vannamei) 养殖池中添加EM菌后可显著降低水中氮和磷的浓度[38]; EM菌可显著提升尼罗罗非鱼 (Oreochromis niloticus) 的生长性能和营养利用率[39];EM菌可显著提高凡纳滨对虾幼体的成活率和体内消化酶活性[40];水体中添加EM菌有助于提高中华绒螯蟹 (Eriocheir sinensis) 抵抗氧化应激的能力,调控其免疫基因的表达和相关酶活性,增强其非特异性免疫力[41]。此外,有些EM菌可以改变甲壳动物的消化道菌群,增强其对病原体的抵抗力[42-43]。本研究在使用EM菌的同时添加了维生素B、维生素C和葡萄糖等,目的在于为EM菌提供营养补充,促进水体微生态系统稳定,使其真正发挥作用。但须注意的是,由于EM菌菌种的变异和退化,应每3~4 d补充1次以维持其菌种优势。本研究结果显示,EM菌实验组稚贝育成率为不添加微生物制剂组的3.26倍,为单投光合细菌组的2.43倍。EM菌实验组的效果好于单投光合细菌组的原因在于前者含有5大功能不同的菌种,净化水质和抑制病原菌的功能更加强大。由于使用EM菌,在不换水的条件下仍能保持良好的水质,避免了频繁换水对幼体的损伤,进而提高了幼体成活率,这是“封闭育苗法”的关键之一;其次,幼虫密度控制在3 个·mL−1,可减少对水体的污染物负荷,使水体微生态平衡更易控制;再者,采用浓缩拟微球藻投喂,可大大减少藻类培养液带来的原生动物、藻类排泄物和病原细菌等的污染。在雷州半岛,因换水导致育苗失败的较为常见,因此,采用封闭育苗法育苗既可节约人力、物力和电力,也可显著提高育苗成功率,大幅提升经济效益。
3.4 立体采苗的效果
钝缀锦蛤生活在潮间带中、低潮区至潮下带沙、沙泥和泥沙底质中,营埋栖生活,且对底质有较强的适应能力[5]。营底栖潜居生活的双壳贝类采用立体采苗法进行采苗,能提高稚贝的附着量,有利于稚贝的生长,也方便对其筛选、冲洗和移植,有效地减轻了稚贝移植过程中的机械损伤[44]。张晓燕等[45]研究发现,在泥蚶 (Tegillarca granosa) 人工育苗中,采用立体附着的效果明显好于池底平面附着,单位水体出苗量可提高3.4~5.3倍;刘德经等[44]研究指出,西施舌 (Coelomactra antiquata) 人工立体采苗的采苗量为传统池底采苗量的1.6倍,且具有成活率高、生长速度快等优势。本研究采用立体采苗方法,出苗量比平面采苗提高了76.5%,与前人的研究一致,但与前两项研究相比,本研究中立体采苗法的出苗量相对对照组的提升率较小,这可能与品种、浮游幼虫投放密度和稚贝收获规格不同有关。本研究立体采苗的稚贝育成率为25.6%,平面采苗的对照组仅为14.5%,其原因主要是对照组池底因高密度附着带来的饵料、空间、溶解氧的竞争导致稚贝大量死亡,而实验组大部分稚贝附着于附着板上,减轻了池底稚贝的生存压力,从而提高了成活率。由此可见,立体采苗法能显著提升稚贝的数量和质量,更适合应用于生产实践。
4. 结论
本研究在室内水池人工控温促熟条件下,采用虾塘复合藻代替纯种单胞藻对亲本进行促熟,解决了亲本高效促熟的问题。采用空气升温加阴干诱导及控制产卵环境等一系列措施,可显著提高亲本催产率。通过添加EM复合菌,育苗期间不换水,可显著提高幼虫浮游期成活率、生长速度和稚贝育成率。采用立体采苗器结合池底进行立体附苗,可提高稚贝的育成率及单位面积产量,同时提高稚贝的生长速度。对体质量进行选育,稚贝的壳长和体质量明显提高,现实遗传力h2分别达0.65和0.72。采用本研究的集成创新技术进行钝缀锦蛤人工育苗,可获得显著的经济效益,为实现种苗的规模化生产奠定了基础。
-
![]()
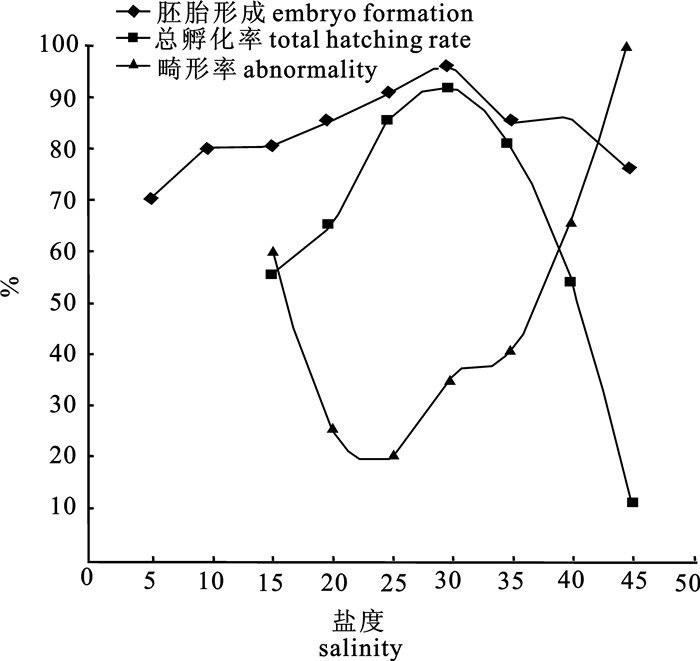
图 1 盐度对卵形鲳鲹胚胎形成、总孵化率、畸形率的影响
Figure 1. Effects of salinity on embryo formation, total hatching rate and abnormality of T.ovatus
表 1 卵形鲳鲹受精卵在不同盐度下的沉浮情况
Table 1 Buoyancy observation of T.ovatus fertilized eggs at different salinities
盐度 salinity 5.0 10.0 15.0 20.0 25.0 30.0 35.0 40.0 45.0 50.0 状态 state 沉 沉 沉 沉 沉 半浮沉 浮 浮 浮 浮  下载: 导出CSV
下载: 导出CSV
表 2 盐度10.0~45.0条件和饥饿状态下卵形鲳鲹仔鱼成活率和生存活力指数
Table 2 Survival rate and SAI of larval T.ovatus under starvation at salinity of 10.0~45.0
盐度
salinity仔鱼不同天数的成活率/%
the survival rate of larval on different daysSAI 1 2 3 4 5 6 7 8 9 10 11 12 10.0 100 52 37 8 0 0 0 0 0 0 0 0 3.17 15.0 100 88 74 69 54 45 13 2 0 0 0 0 16.43 20.0 100 93 90 85 84 83 54 34 16 2 0 0 26.18 25.0 100 98 95 95 93 88 86 73 70 60 4 0 44.19 30.0 100 99 98 98 98 89 89 86 85 60 12 0 47.44 35.0 100 96 93 90 89 80 70 15 0 0 0 0 24.66 40.0 100 85 70 63 49 35 13 6 0 0 0 0 13.26 45.0 100 0 0 0 0 0 0 0 0 0 0 0 1.00
下载: 导出CSV
-
[1] FERRANRO S P. Embryonic development of Altantic menhaden (Brevoortia tyrannus) and a fish embryo age estimation[J]. Fish Bull, 1980, 77(4): 943-949. https://www.semanticscholar.org/paper/EMBRYONIC-DEVELOPMENT-OF-ATLANTIC-MENHADEN%2C-AND-A-Ferrar/8fa8ca4455356b587cb081e25ac51cef6c584b58
[2] 雷霁霖, 孙鲁宁, 陈学豪. 盐度对黑鲷胚胎发育和早期仔鱼发育影响的初步观察[J]. 海洋水产研究, 1986(7): 143-147. [3] 王永新, 陈建国, 孙帼英. 温度和盐度对花鲈胚胎及前期仔鱼发育影响的初步报告[J]. 水产科技情报, 1995, 22(2): 54-57. doi: 10.16446/j.cnki.1001-1994.1995.02.002 [4] 黄永春, 郑建辉, 周泽斌. 盐度对NFDBF状黄姑鱼胚胎发育和仔鱼成活的影响[J]. 福建水产, 1997, (1): 34-37. doi: 10.14012/j.cnki.fjsc.1997.01.007 [5] 葛国昌, 宋卫红. 盐度对梭鱼卵和初孵仔鱼的作用[J]. 山东海洋学院学报, 1985, 15(1): 93-97. doi: 10.16441/j.cnki.hdxb.1985.01.011 [6] MURASHIGE R, BASS P, WALLACE L, et al. The effect of salinity on the survival and growth of striped mullet (Mugil cephalus) larvae in the hatchery[J]. Aquac, 1991, 96(3/4): 249-254. doi: 10.1016/0044-8486(91)90155-Z
[7] KARAS P, KLINGSHEIM V. Effects of temperature and salinity on embryonic development of turbot (Scophthalrnus maximus L. ) from the North Sea, and comparisons with Baltic populations[J]. Helgolander Meeresuntersuchungen, 1997, 51(2): 241-247. doi: 10.1007/BF02908710
[8] 麦贤杰, 黄伟健, 叶富良, 等. 海水鱼类繁殖生物学和人工繁育[M]. 北京: 海洋出版社, 2005: 1-345. https://d.wanfangdata.com.cn/periodical/Ch9QZXJpb2RpY2FsQ0hJTmV3UzIwMjQxMTA1MTcxMzA0Eg5RSzIwMDUwMjI5ODU1MxoIeGE4bGR0amo%3D [9] 李加儿, 李正森, BANNO J E. 遮目鱼受精卵和早期仔鱼对盐度的耐受性[M]//海洋水产科学研究文集. 广州: 广东科技出版社, 1999: 147-151. [10] 成庆泰, 郑葆珊. 中国鱼类系统检索[M]. 北京: 科学出版社, 1987: 341-342. https://baike.baidu.com/item/%E4%B8%AD%E5%9B%BD%E9%B1%BC%E7%B1%BB%E7%B3%BB%E7%BB%9F%E6%A3%80%E7%B4%A2/59235508 [11] 陈伟洲, 许鼎盛, 王德强, 等. 卵形鲳鲹人工繁殖及育苗技术研究[J]. 台湾海峡, 2007, 26(3): 435-442. doi: 10.3969/j.issn.1000-8160.2007.03.019 [12] 区又君, 李加儿. 卵形鲳鲹的早期胚胎发育[J]. 中国水产科学, 2005, 12(6): 787-789. doi: 10.3321/j.issn:1005-8737.2005.06.019 [13] 南京农学院. 田间试验和统计方法[M]. 北京: 农业出版社, 1979: 85-92. [14] 谢仰杰, 翁朝红, 林锦宗, 等. 盐度对花尾胡椒鲷胚胎和仔鱼的影响[J]. 台湾海峡, 2000, 19(1): 31-35. https://kns.cnki.net/kcms2/article/abstract?v=HbazLCXbuSWEHNmm4Oc4E3hYJ5JJUGoWrUchaUOGM7myK1wownFFPBkGtcY4lIzOo-8I4XDhSAMfsZaMmPn3DfiENGDJqMGBJaarxXoxwb3ojqQLd8JXHF0X8jz99vNkveC84WTG8hxZnTiPC1wOkP89ZXgAMAbjNKUc-vbx0qqg8kevI7rINqlaPPw22j9h&uniplatform=NZKPT&language=CHS [15] 施兆鸿, 陈波, 彭士明, 等. 盐度胁迫下点胚胎点带石斑鱼(Epinephelus malabaricus)及卵黄囊仔鱼的形态变化[J]. 海洋与湖沼, 2008, 39(3): 222-226. doi: 10.3321/j.issn:0029-814X.2008.03.005 [16] 陈昌生, 纪荣兴, 黄佳鸣, 等. 高体NFDA4胚胎、早期仔鱼发育与盐度的关系[J]. 上海水产大学学报, 1997, 6(1): 5-10. https://kns.cnki.net/kcms2/article/abstract?v=HbazLCXbuSVHRZTpzxeCEq7LrYeL83jP5DeRrPlWnP0wiwiOeb4aAw8C5mlZXUIaixyIWV4bqXr9s4lXkg7QDaya6MrKeqryiIcmdq8FP2J0liOUo1AH0KGtCIaWeC8wP1db7O0yWNd6K9oBx0BrW5SGXx_95s0JMKJQyjkmO9jpWa8mJRIJViQ3EUF_3i5l&uniplatform=NZKPT&language=CHS [17] 施兆鸿, 夏连军, 王建钢, 等. 盐度对黄鲷胚胎发育及早期仔鱼生长的影响[J]. 水产学报, 2004, 28(5): 599-602. doi: 10.3321/j.issn:1000-0615.2004.05.021 [18] 孙丽华, 陈浩如, 王肇鼎. 盐度对军曹鱼胚胎和仔鱼发育的影响[J]. 生态科学, 2006, 25(1): 48-51. doi: 10.3969/j.issn.1008-8873.2006.01.013 [19] SWANSON C. Early development of milkfish: Effects of salinity on embryonic and larval metabolism, yolk absorption and growth[J]. Fish Biol, 1996, 48(3): 405- 421. doi: 10.1111/j.1095-8649.1996.tb01436.x
[20] 张海发, 刘晓春, 王云新, 等. 温度、盐度及pH对斜带石斑鱼受精卵孵化和仔鱼活力的影响[J]. 热带海洋学报, 2006, 25(2): 3l-37. doi: 10.3969/j.issn.1009-5470.2006.02.006 [21] 鵜川正雄. 赤点石斑鱼产卵习性和初期生活史[J]. 鱼类学杂志, 1966, 13(4/6): 156-161. [22] 王涵生, 方琼珊, 郑乐云. 盐度对赤点石斑鱼受精卵发育的影响及仔鱼活力的判断[J]. 水产学报, 2002, 26(4): 344-351. doi: 10.3321/j.issn:1000-0615.2002.04.009






计量
- 文章访问数: 4641
- HTML全文浏览量: 166
- PDF下载量: 3222
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
