
Five kinds of isozyme in different tissues of Cheilinus undulatus
-
摘要:
运用聚丙烯酰胺垂直梯度凝胶电泳法对波纹唇鱼(Cheilinus undulatus)6组织(肌肉、心脏、肝脏、肾脏、脑和脾脏)的5种同工酶[酯酶(EST)、乳酸脱氢酶(LDH)、苹果酸脱氢酶(MDH)、苹果酸酶(ME)和天门冬氨酸氨基转移酶(AST)]进行了研究,并对其同工酶位点及其酶谱表型进行了分析。结果表明,波纹唇鱼5种同工酶系统具有不同程度的组织特异性。EST检测到2条酶带,由2个基因座位编码。LDH在6种组织中都存在,共检测到9条酶带,由3个基因座位编码,具有多态性现象。MDH在6种组织中均可检测到,可分为线粒体型和上清液型,各由1个基因座位编码,组织差异性不显著。ME检测到2条酶带,肝脏的含量比较丰富,线粒体型和上清液型的区分不明显。AST在6种组织中都没有发现。
Abstract:Five isozymes (EST, ME, AST, LDH and MDH) in six tissues (muscle, heart, liver, kidney, brain and spleen) of Cheilinus undulatus were studied by using vertical polyacrylamide gel gradient electrophoresis. The results showed that the five isozymes presented tissues specificity to some degrees. The number of EST bands was 2, which were encoded by 2 loci. LDH existed in all tissues studied. The number of LDH bands was 9, which were encoded by 3 loci, and the polymorphism was strong. Two types of MDH (m-MDH and s-MDH) were found and also existed in the six tissues. The number of ME bands was 2. ME in liver were rich, but it was not distinct between m-ME and s-ME. There was no AST in all six tissues.
-
Keywords:
- Cheilinus undulates /
- isozyme /
- expression /
- tissues specificity
-
卡拉胶是从麒麟菜(Eucheuma)、角叉菜(Chondrus)、杉菜(Gigartina)和沙菜(Hypnea)等属的海洋红藻中提取,由D-半乳糖残基α-(1→3)和β-(1→4)交替键合而成的重复结构的高分子多糖硫酸酯[1-3]。现在已知的卡拉胶类型有7种:κ-、μ-、ι-、ν-、λ-、θ-和ξ-卡拉胶。其中只有κ-和ι-卡拉胶具有胶凝性,其他类型的卡拉胶在水中都不能形成凝胶,但通过碱处理可以使μ-和ν-卡拉胶分别转变成κ-和ι-卡拉胶,提高其凝胶强度[4-6]。
凝胶强度是衡量卡拉胶质量的一个重要指标,碱处理能使卡拉胶中的硫酸基含量减少,306-内醚-半乳糖(306-AG)含量提高,使卡拉胶的类型发生改变,从而提高卡拉胶的凝胶强度[7]。碱处理是卡拉胶生产中必不可少的重要工序,目前碱处理方法主要有高温浓碱法、中温浓碱法和常温浓碱法。中、高温浓碱法的优点是生产周期短,缺点是卡拉胶流失较多、产率低、需耗热能、需特殊设备,常温浓碱法的优点是生产简单且易控制、产胶率高,无需耗能,设备简单,处理成本低,缺点是耗碱量较大,生产周期较长[8]。常温浓碱法是目前国内外卡拉胶加工企业重点关注的方法。卡拉胶提取过程中的碱处理工艺研究已有许多报道[9-11],基本均采用热碱法处理,温度范围为45~65 ℃,处理时间为2~15 h,卡拉胶产率为30%左右,凝胶强度1 000 g · cm-2以上(质量浓度15 g · L-1)。有关常温碱处理麒麟菜的优化工艺尚未见报道。文章采用常温浓碱法处理麒麟菜,以凝胶强度和产率为研究对象,并结合响应面分析法优化碱处理条件,确定碱处理最佳工艺,以为麒麟菜卡拉胶的工业生产提供依据。
1. 材料与方法
1.1 材料与仪器
1.1.1 异枝麒麟菜
由江门市丰正食品有限公司提供。
1.1.2 氢氧化钠(NaOH)、氯化钾(KCl)
均为分析纯。
1.1.3 仪器
Alpha 1-4 LSC型冷冻干燥机(德国Christ公司出品);QTS 25型质构仪[美国Brookfield(博力飞)公司出品];其他均为实验室常用仪器设备。
1.2 方法
1.2.1 卡拉胶提取工艺流程[12]
原料预处理→碱处理→洗至中性→漂白、脱氯→提胶→过滤→冷却凝胶→冻结脱水→干燥→成品。工艺流程简述:1)预处理。试验前用水洗去原料中的泥沙及其他杂物;2)碱处理。将麒麟菜放入一定浓度的NaOH溶液中常温浸泡一定时间,碱处理后排去碱液,用水反复浸洗至近中性;3)漂白、脱氯。选用次氯酸钠作为漂白剂,其中有效氯质量浓度为2 g · L-1,在溶液pH 4.5的条件下漂白5 min;漂白后用0.4%的硫代硫酸钠进行脱氯处理5~10 min;4)提胶。将处理好的原料放入2 L烧杯中,按m (麒麟菜) ∶ V (碱液)为1∶ 30(g · mL-1)的比例在90 ℃恒温水浴中提取1 h;5)过滤。用200目的尼龙布趁热过滤胶液,胶液用烧杯收集;6)冷却凝胶。加入胶液量0.2%的氯化钾溶液,充分搅拌,在室温下自动冷却形成凝胶;7)冻结脱水。将凝胶置-18 ℃冰箱中冷冻至完全冻结,自然解冻脱水;8)干燥、粉碎。将脱水后的卡拉胶在70 ℃下烘干,用粉碎机粉碎,过80目筛即得卡拉胶粉。
1.2.2 响应面法优化碱处理工艺
1) 单因素试验。选取对卡拉胶产率和凝胶强度有显著影响的3个因素,即碱液浓度、浸泡时间和浸泡温度进行单因素试验;2)响应曲面试验设计。根据单因素试验结果,选取对响应值(产率、凝胶强度)有显著影响的3个因素进行响应面试验。采用SAS 8.0数据统计分析软件对试验数据进行回归分析。每个自变量的低、中、高水平分别以-1、0、1进行编码,采用最小二乘法拟合二次多项方程[13-14]。
1.2.3 凝胶强度测定方法
按照参考文献[15]的方法略作改动。称取卡拉胶粉1.0 g(折算为干物质)于100 mL烧杯中,加入去离子水100 g,记录总质量为m,水浴加热搅拌使其充分溶解,再用去离子水补充使其总质量为m,搅拌均匀,保鲜膜封口过夜、至20 ℃下静置15 h,在质构仪平台上用1 cm2的探头测定其凝胶强度。
1.2.4 产率测定
$$ X=\frac{M_1 \times 1-W_1}{M_2 \times 1-W_2} $$ 式中X为产率(%);M1为产品质量(g);M2为原料质量(g);W1为产品水分质量分数(%);W2为原料水分质量分数(%)。
1.2.5 数据处理
试验数据用Design-Expert 8.05及Excel软件处理。
2. 结果与分析
2.1 单因素试验2.1.1碱液质量浓度对产率和凝胶强度的影响
分别配制质量浓度为120 g · L-1、160 g · L-1、200 g · L-1、240 g · L-1、280 g · L-1和320 g · L-1的NaOH溶液,按m (麒麟菜) ∶ V (碱液)=1∶ 12(g ·mL-1)的比例投入浸泡,固定浸泡温度25 ℃、浸泡时间3 d,考察碱液质量浓度对卡拉胶产率和凝胶强度的影响,每个质量浓度作3个平行试验。随着碱液质量浓度的升高,卡拉胶的凝胶强度逐渐增大,ρ (碱液)为240 g · L-1时卡拉胶GS为623 g ·cm-2,ρ (碱液)为280 g · L-1时GS达到最大值(694 g ·cm-2),此后趋于平衡;而卡拉胶的产率随着碱液质量浓度的升高缓慢下降,ρ (碱液)为240 g · L-1时产率为27.4%,ρ (碱液)为280 g · L-1时产率为25.7%(图 1)。原因在于碱处理可去除卡拉胶结构中的硫酸基,提高卡拉胶的凝胶强度,还可使无凝胶能力的μ-、ν-等卡拉胶转化为具有凝胶能力的κ-和ι-卡拉胶[12]。在一定碱质量浓度范围内碱质量浓度越高这种转化越彻底,因此,凝胶强度随碱液质量浓度的提高而增大,但是碱液质量浓度过高,溶液粘稠不利于碱液向麒麟菜组织的扩散,或是卡拉胶的化学结构转化已达到平衡,卡拉胶凝胶强度最终趋于平衡。而卡拉胶产率随着碱质量浓度的提高而降低,这可能是因为高质量浓度的碱液对藻体细胞有所损害,导致藻体胶质流入碱液,造成胶体流失[10, 16],也可能是由于碱处理使麒麟菜藻体变硬,提胶时难以出胶,使提胶率下降。
2.1.2 浸泡时间对产率和凝胶强度的影响
在固定碱液质量浓度240 g · L-1、浸泡温度25 ℃条件下考察浸泡时间对卡拉胶产率和凝胶强度的影响,每个浸泡时间作3个平行试验。随浸泡时间的延长,卡拉胶凝胶强度不断增大,最后趋于平衡(图 2)。浸泡前3 d内凝胶强度呈近似直线上升趋势,第1天GS为453 g · cm-2,第3天GS为551 g· cm-2,到第5天GS为584 g · cm-2;而卡拉胶产率随着浸泡时间的延长呈下降趋势,第1天卡拉胶产率为31.4%,第3天为30.7%,此后产率基本呈直线下降趋势,第5天产率为25.1%,到第9天仅为17.1%。这可能是由于麒麟菜浸泡时间越长,硫酸基去除越彻底,无凝胶性的μ-、ν-等卡拉胶转化为凝胶性的κ-和ι-卡拉胶的量也越多,因此,卡拉胶GS随碱处理时间的延长而增强;而在麒麟菜浸泡过程中藻体会发生部分胶溶现象[11],导致胶体流入碱液造成损失,并且在试验中也发现,浸泡时间越长,胶体自溶、流失也越严重,卡拉胶的产率越低。
2.1.3 浸泡温度对产率和凝胶强度的影响
在固定碱液浓度240 g · L-1、浸泡时间3 d条件下考察浸泡温度对卡拉胶产率和凝胶强度的影响,每个浸泡温度作3个平行试验。随着浸泡温度的升高,卡拉胶的凝胶强度和产率均呈先上升后下降的趋势,在浸泡温度为25 ℃时凝胶强度和产率达最大值,分别为673 g · cm-2和31.9%;浸泡温度超过25 ℃后GS和产率不断下降,当浸泡温度为35 ℃时GS和产率均达最低值,分别为571 g · cm-2和17.8%(图 3)。这可能是由于麒麟菜卡拉胶发生化学结构转化时有一定的最适温度范围,在此范围内随温度的升高,化学结构转化和硫酸基去除趋于彻底,因此,凝胶强度不断增强,但当温度超过该范围,加之长时间处理,卡拉胶发生了降解,造成卡拉胶GS降低[17],同时浸泡温度越高,卡拉胶越易发生溶胶,卡拉胶流失也越严重,产率不断降低。
2.2 响应面优化试验
2.2.1 试验安排与结果
根据单因素试验结果,确定各影响因素的编码水平。X1、X2、X3分别代表碱液质量浓度(g · L-1)、浸泡时间(d)、浸泡温度(℃)3个响应变量,Y1、Y2分别代表卡拉胶凝胶强度(g · cm-2)、产率(%)2个响应值,因素水平见表 1,试验安排与结果见表 2[18-20]。每个试验作3个平行。
表 1 响应面因素水平表Table 1. Factors and levels in response surface design因素
factors水平levels -1 0 1 X1碱液质量浓度/g·L-1
concetration200 240 280 X2浸泡时间/d
soaking time3 5 7 X3浸泡温度/℃
soaking temperature20 25 30 表 2 响应面试验设计及结果(X±SD)Table 2. Design and results of response surface试验号
test No.X1 X2 X3 Y1/g·cm-2 Y2/% 1 1 1 0 567±14.07 28.48±2.01 2 1 0 1 641±10.10 30.71±1.41 3 0 1 -1 558±11.42 24.11±1.78 4 1 0 -1 478±12.40 21.19±1.82 5 1 -1 0 516±17.26 25.81±1.43 6 0 0 0 704±10.79 31.94±1.54 7 0 -1 1 517±9.82 26.32±1.63 8 -1 0 -1 474±13.16 23.69±1.12 9 0 -1 -1 423±8.81 21.24±1.26 10 -1 0 1 569±15.27 26.21±2.10 11 0 1 1 649±11.77 30.34±1.33 12 0 0 0 709±16.04 33.83±1.38 13 0 0 0 685±13.61 34.13±1.80 14 -1 -1 0 433±10.01 22.51±0.81 15 -1 1 0 592±16.16 28.49±1.22 2.2.2 模型建立与方差分析
对凝胶强度和产率的模型建立与方差分析。采用Design-Expert统计软件,利用最小二乘法预测各个响应变量的二次多项式模型的回归系数(表 3)。
表 3 响应变量二次多项式模型的回归系数分析Table 3. Regression coefficients of predicated second-order polynomial model for response variables项目
item系数coefficient 标准误差standard error t P Y1 Y2 Y1 Y2 Y1 Y2 Y1 Y2 常量constant 699.33 33.30 9.78 0.48 52.78 69.12 0.00 0.00 X1 16.75 0.66 5.99 0.30 2.06 2.24 0.09 0.08 X2 59.63 1.94 5.99 0.30 7.81 6.58 0.00 0.00 X3 55.38 2.92 5.99 0.30 6.36 9.89 0.00 0.00 X1X2 -27.00 -0.83 8.47 0.42 -2.35 -1.98 0.07 0.10 X1X3 17.00 1.75 8.47 0.42 1.48 4.19 0.19 0.06 X2X3 -0.75 0.29 8.47 0.42 -0.72 0.69 0.50 0.52 X12 -84.29 -3.51 8.81 0.43 -7.37 -8.09 0.00 0.00 X22 -88.04 -3.46 8.81 0.43 -7.97 -50.25 0.00 0.00 X32 -74.54 -4.34 8.81 0.43 -5.93 -9.98 0.00 0.00 根据表 3回归系数可得X1、X2、X3对Y1、Y2的二次多项式回归模型:
Y1=699.33+16.75X1+59.63X2+55.38X3-27.00X1X2+17.00X1X3-0.75X2X3-84.29X12-88.04X22-74.54X32
Y2=33.30+0.66X1+1.94X2+2.92X3-0.83X1X2+1.75X1X3-0.29X2X3-3.51X12-3.46X22-4.34X32
由表 3中t-检验可见,对于响应值凝胶强度Y1和产率Y2,X2、X3的一次项和X1、X2、X3的二次项呈极显著(P<0.01),X1一次项和X1、X2、X3项的交互项均不显著(P>0.05)。这表明,试验中的各因素与响应值间并不是简单的线性关系。对回归模型进一步进行方差和误差分析,结果见表 4。
表 4 回归模型方差和误差分析Table 4. Variance and error analysis of regression model for GS来源
source自由度
df平方和
square sum均方和
mean squareF P R2 Y1 Y2 Y1 Y2 Y1 Y2 Y1 Y2 Y1 Y2 Y1 Y2 模型model 9 9 124 700.00 255.58 1 385.44 28.40 48.33 40.79 0.00 0.00 0.99 0.98 X1 1 1 2 244.50 3.50 2 244.50 3.50 7.83 5.02 0.04 0.03 X2 1 1 28 441.13 30.19 28 441.13 30.19 99.21 43.36 0.00 0.00 X3 1 1 24 531.13 68.15 24 531.13 68.15 85.57 97.89 0.00 0.00 X1X2 1 1 2 916.00 2.74 2 916.00 2.74 10.17 3.93 0.02 0.10 X1X3 1 1 1 156.00 12.25 1 156.00 12.25 4.03 17.59 0.10 0.01 X2X3 1 1 2.25 0.33 2.25 0.33 0.007 85 0.47 0.93 0.52 X12 1 1 26 234.16 45.62 26 234.16 45.62 91.51 65.52 0.00 0.00 X22 1 1 28 620.31 44.27 28 620.31 44.27 99.83 63.58 0.00 0.00 X32 1 1 20 516.16 69.39 20 516.16 69.39 71.56 99.66 0.00 0.00 残差residual 5 5 1 433.42 3.48 286.68 0.70 失拟lack of fit 3 3 1 112.75 0.66 370.92 0.22 2.31 0.16 0.32 0.92 纯误差pure error 2 2 320.67 2.82 160.33 1.41 合计total 14 14 126 100.00 259.06 Y1和Y2回归模型呈极显著(P < 0.01),并且其失拟性都不显著(P>0.05),其决定系数(R2)分别为0.99和0.98,说明2个回归模型能够较好地预测相应的响应变量。一次项和平方项呈极显著性(P < 0.01),说明响应值Y1和Y2主要是由回归模型的一次项和平方项所决定。
2.2.3 响应曲面分析
由拟合的回归模型作响应面和等高线图,分别固定变量碱液质量浓度(X1)、浸泡时间(X2)和浸泡温度(X3)在原点,分别得到另外2个因素的响应面和等高线图(图 4~图 9)。
![]() 图 4 Y1=(X1, X2)的响应面与等高线Figure 4. Response surface and contour plots under Y1=(X1, X2)
图 4 Y1=(X1, X2)的响应面与等高线Figure 4. Response surface and contour plots under Y1=(X1, X2)![]() 图 5 Y1=(X1, X3)的响应面与等高线Figure 5. Response surface and contour plots under Y1=(X1, X3)
图 5 Y1=(X1, X3)的响应面与等高线Figure 5. Response surface and contour plots under Y1=(X1, X3)![]() 图 6 Y1=(X2, X3)的响应面与等高线Figure 6. Response surface and contour plots under Y1 = (X2, X3)
图 6 Y1=(X2, X3)的响应面与等高线Figure 6. Response surface and contour plots under Y1 = (X2, X3)![]() 图 7 Y2=(X1, X2)的响应面与等高线Figure 7. Response surface and contour plots under Y2=(X1, X2)
图 7 Y2=(X1, X2)的响应面与等高线Figure 7. Response surface and contour plots under Y2=(X1, X2)![]() 图 8 Y2=(X1, X3)的响应面与等高线Figure 8. Response surface and contour plots under Y2=(X1, X3)
图 8 Y2=(X1, X3)的响应面与等高线Figure 8. Response surface and contour plots under Y2=(X1, X3)![]() 图 9 Y2=(X2, X3)的响应面与等高线Figure 9. Response surface and contour plots under Y2= (X2, X3)
图 9 Y2=(X2, X3)的响应面与等高线Figure 9. Response surface and contour plots under Y2= (X2, X3)等高线图近似圆形表明交互项的影响不显著,这与表 3、表 4中的回归系数和模型方差显著性的检验结果一致。响应面呈规则的凸起形表明在试验因素水平范围内存在极大值,即响应面的最高点[21]。
2.2.4 最优工艺参数确定
为了进一步得到各因素的最佳条件组合,使得卡拉胶凝胶强度和产率达到最优值,采用Design-Expert软件对各因素和两响应值的数据进行优化分析。在凝胶强度方面,通过分析得到X1、X2、X3的编码值分别为0.138、0.240、0.316,换算得到相应的实际碱液浓度X1=250 g · L-1、浸泡时间X2=3.48 d、浸泡温度X3=26.58 ℃,优化的卡拉胶凝胶强度理论值为719.13 g · cm-2。在产率方面,通过分析得到X1、X2、X3的编码值分别为0.201、0.315、0.415,换算得相应的实际值碱液浓度X1=250 g · L-1、浸泡时间X2=3.63 d、浸泡温度X3=27.08 ℃,优化的卡拉胶产率理论值为34.15%。从实际操作便利方面考虑,将碱处理最佳条件取为碱液浓度250 g ·L-1、浸泡时间3.5 d、浸泡温度25 ℃。在此优化条件下进行3次重复验证试验,结果显示卡拉胶平均凝胶强度和产率分别为721 g · cm-2和35.17%。结果与理论预测值基本吻合,表明响应面分析优化结果与实际值相一致,对卡拉胶提取的碱处理参数具有一定的实际预测价值。
3. 结论
通过Box-Benhnken的中心组合设计响应面法建立了3个影响因素(碱液质量浓度、浸泡时间及浸泡温度)和2个响应值(凝胶强度及产率)之间相互作用的数学模型,得出碱处理时的最佳工艺参数为碱液质量浓度250 g · L-1,浸泡时间3.5 d,浸泡温度25 ℃。按此优化工艺条件对麒麟菜进行碱处理,得到卡拉胶的凝胶强度和产率的最大值分别为721 g · cm-2(10 g · L-1)和35.17%,有效提高了麒麟菜卡拉胶的产率和产品质量。建立的数学模型能较好地预测各试验因素和考查指标之间的关系。
-
![]()
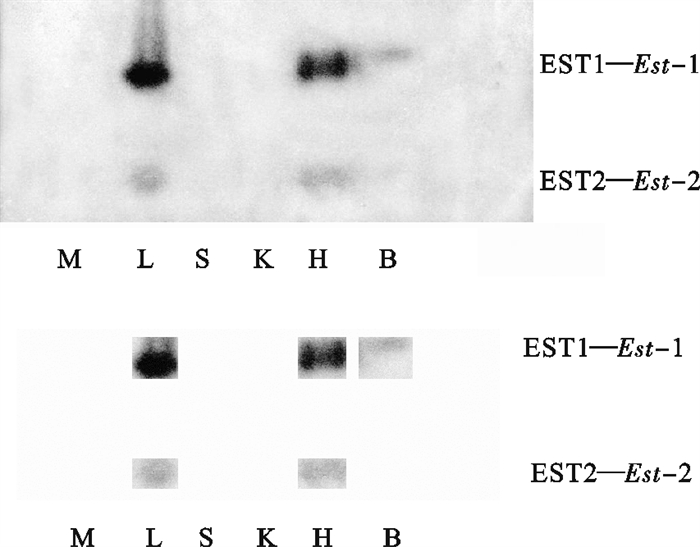
图 1 波纹唇鱼不同组织的EST电泳图谱
M. 肌肉;L.肝脏;S. 脾脏;K. 肾脏;H. 心脏;B. 脑
Figure 1. Electrophoretogram of EST in tissues of C.undulatus
M. muscle; L.liver; S. spleen; K. kidney; H. heart; B. brain
![]()
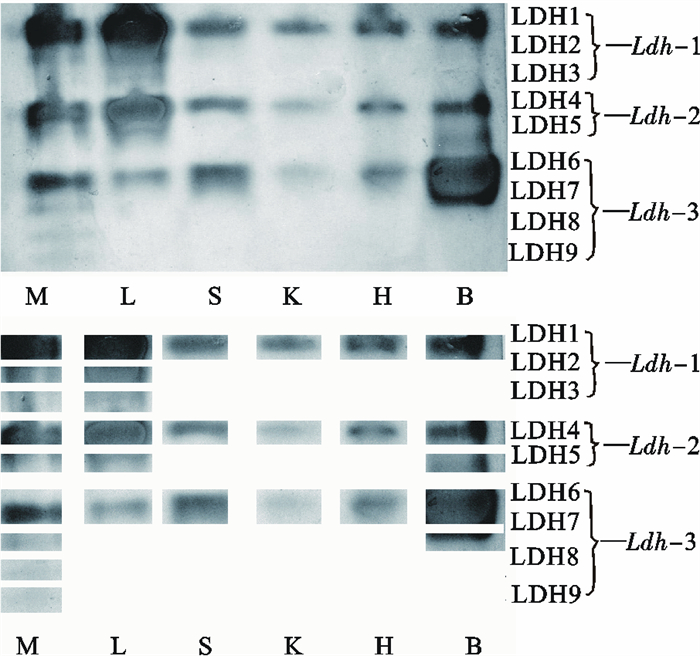
图 2 波纹唇鱼不同组织的LDH电泳图谱
M. 肌肉;L.肝脏;S. 脾脏;K. 肾脏;H. 心脏;B. 脑
Figure 2. Electrophoretogram of LDH in tissues of C.undulatus
M. muscle; L.liver; S. spleen; K. kidney; H. heart; B. brain
![]()
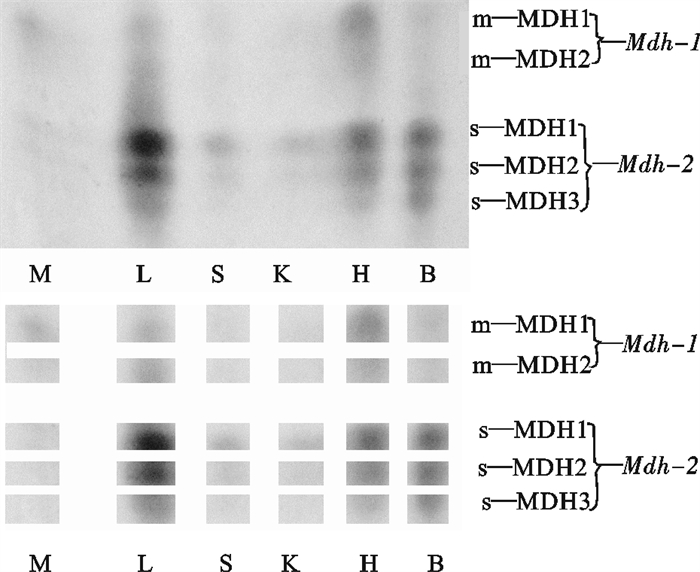
图 3 波纹唇鱼不同组织的MDH电泳图谱
M. 肌肉;L.肝脏;S. 脾脏;K. 肾脏;H. 心脏;B. 脑
Figure 3. Electrophoretogram of MDH in tissues of C.undulatus
M. muscle; L.liver; S. spleen; K. kidney; H. heart; B. brain
![]()
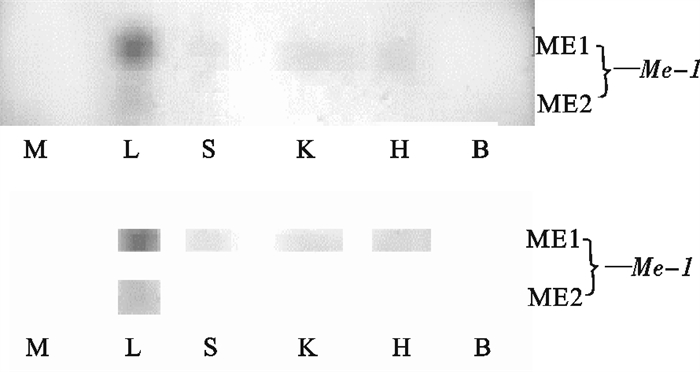
图 4 波纹唇鱼不同组织的ME电泳图谱
M. 肌肉;L. 肝脏;S. 脾脏;K. 肾脏;H. 心脏;B. 脑
Figure 4. Electrophoretogram of ME in tissues of C.undulatus
M. muscle; L. liver; S. spleen; K. kidney; H. heart; B. brain
表 1 用于分析波纹唇鱼同工酶名称、编号、缓冲系统和凝胶浓度
Table 1 The name, serial number, buffer system and gel concentration for analyzing the isozyme of C.undulatus
酶的名称name of enzyme 编号serial number 缓冲系统buffer system 梯度胶/% gel concentration 酯酶EST 3.1.1.1 TVB4~28 4~28 乳酸脱氢酶LDH 1.1.1.27 TC 4~15 苹果酸脱氢酶MDH 1.1.1.37 TC 4~7.5 苹果酸酶ME 1.1.1.40 TC 4~7.5 天门冬氨酸氨基转移酶AST 2.6.1.1 TC 4~15 注:TC. Tris-柠檬酸(pH 7.0);TVB. Tris-硼酸-EDTA(pH 8.4)
Note:TC. Tris-citric acid(pH 7.0);TVB. Tris-boric acid-EDTA(pH 8.4) 下载: 导出CSV
下载: 导出CSV
-
[1] 胡能书, 万贤国. 同工酶技术及其应用[M]. 长沙: 湖南科技出版社, 1985: 86-12. https://xueshu.baidu.com/usercenter/paper/show?paperid=a2879165a1e8dde93ff1b62cfb459820 [2] 何忠效, 张树政. 电泳[M]. 2版. 北京: 科学出版社, 1999: 284-308. https://xueshu.baidu.com/usercenter/paper/show?paperid=6b6975e12431a514f1430f1bb9b12628 [3] 郭尧君. 蛋白质电泳实验技术[M]. 北京: 科学出版社, 1999: 70-80. [4] SHALK LEE J B, ALLINDORF F W, MORIZOT D C, et al. Genetic nomenclature for protein-coding loci in fish[J]. Trans Amer Fish Soc, 1990, 119(1): 2-15. doi: 10.1577/1548-8659(1990)119<0002:GNFPLI>2.3.CO;2
[5] 邓思平, 刘楚吾. 赤点石斑鱼4种同工酶的组织分布及基因位点分析[J]. 浙江海洋学院学报: 自然科学版, 2004, 23(2): 103-106. doi: 10.3969/j.issn.1008-830X.2004.02.002 [6] 姜建国, 熊全沫, 姚汝华. 草鱼同工酶的组织分布及遗传结构分析[J]. 华南理工大学学报: 自然科学版, 1997, 25(12): 105-110. doi: 10.3321/j.issn:1000-565X.1997.12.020 [7] 刘文彬, 陈合格, 张轩杰. 黄颡鱼不同组织中同工酶的表达模式[J]. 激光生物学报, 2003, 12(14): 274-278. doi: 10.3969/j.issn.1007-7146.2003.04.007 [8] 尹绍武, 廖经球, 黄海, 等. 褐点石斑鱼不同组织4种同工酶的研究[J]. 海洋通报, 2007, 26(1): 472-474. doi: 10.3969/j.issn.1001-6392.2007.01.006 [9] 尹绍武, 雷从改, 黄海, 等. 点带石斑鱼不同组织同工酶的研究[J]. 水产科学, 2006, 25(9): 471-474. doi: 10.3969/j.issn.1003-1111.2006.09.010 [10] 丁少雄, 王军, 郭丰, 等. 台湾海区斜带石斑鱼群体遗传学的等位酶研究[J]. 厦门大学学报: 自然科学版, 2003, 42(5): 645-651. https://www.doc88.com/p-5079933552133.html [11] 金春华, 钟爱华, 张海琪, 等. 大弹涂鱼不同组织器官的同工酶研究[J]. 海洋科学, 2004, 28(3): 35-40. doi: 10.3969/j.issn.1000-3096.2004.03.009 [12] 姜建国, 熊全沫, 姚汝华. 鳙鱼不同组织的同工酶研究[J]. 华南理工大学学报, 1998, 26(3): 68-72. doi: 10.3321/j.issn:1000-565X.1998.03.014 [13] 徐成, 王可玲, 尤锋, 等. 鲈鱼群体生化遗传学研究Ⅰ. 同工酶的生化遗传分析[J]. 海洋与湖沼, 2001, 32(1): 42-49. https://lib.cqvip.com/Qikan/Article/Detail?id=4918138&from=Qikan_Article_Detail [14] 冯秀妮, 张文兵, 麦康森, 等. 饲料中维生素B6对皱纹盘鲍幼鲍蛋白质代谢的影响[J]. 海洋科学, 2006, 30(10): 51-53. doi: 10.3969/j.issn.1000-3096.2006.10.012 -
期刊类型引用(8)
1. 余为,刘赫威,陈新军. 气候变化对西南大西洋渔业生态系统的影响研究进展. 上海海洋大学学报. 2024(01): 211-229 .  百度学术
百度学术
2. 崔国辰,玄文丹,魏裙倚,陶迎新,苏施,余谦超,朱文斌. 基于SST和Chl-a的西南大西洋阿根廷滑柔鱼渔场时空变动分析. 浙江海洋大学学报(自然科学版). 2023(01): 10-19 . 百度学术
3. 章贤成,汪金涛,陈新军. 基于BP神经网络的阿根廷滑柔鱼资源CPUE标准化研究. 渔业科学进展. 2022(02): 11-20 . 百度学术
4. 刘赫威,余为,陈新军,汪金涛,张忠. 南极海冰变动对西南大西洋阿根廷滑柔鱼资源丰度及空间分布的影响. 水产学报. 2021(02): 187-199 . 百度学术
5. 刘赫威,余为,陈新军. 西南大西洋阿根廷滑柔鱼资源及其对环境响应的研究进展. 中国水产科学. 2020(10): 1254-1265 . 百度学术
6. 宣思鹏,陈新军,林东明,陈子末. 西南大西洋阿根廷滑柔鱼雄性个体的有效繁殖力特性研究. 水生生物学报. 2018(04): 800-810 . 百度学术
7. 孔啸兰,晏磊,张鹏,陈作志. 南海扁舵鲣生物学特性的初步研究. 南方水产科学. 2015(05): 100-107 . 本站查看
8. 刘连为,陈新军,许强华,傅建军. 阿根廷滑柔鱼2个产卵群体遗传变异的微卫星分析. 海洋渔业. 2013(02): 125-130 . 百度学术
其他类型引用(6)









计量
- 文章访问数: 5167
- HTML全文浏览量: 169
- PDF下载量: 3225
- 被引次数: 14
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
