
Comparative analysis of muscle quality, intestinal morphology and microbial composition in two cultured frogs species
-
摘要:
为分析泰国虎纹蛙 (Hoplobatrachus tigerinus) 和牛蛙 (Lithobates catesbeiana) 的肌肉品质、肠道形态及微生物构成,采用食品安全国家标准测定方法对这两种养殖蛙的肌肉营养成分进行监测,结合肌肉质构和显微结构的分析对两者肌肉品质进行评价,同时利用高通量测序和组织学切片技术对两者肠道微生物群落构成和显微结构进行分析。结果显示,泰国虎纹蛙和牛蛙氨基酸总量差异不显著 (P>0.05);牛蛙的不饱和脂肪酸含量显著高于泰国虎纹蛙 (P<0.05)。此外,牛蛙肌肉的硬度、咀嚼性和胶黏性均显著高于泰国虎纹蛙 (P<0.05)。肠道微生物分析显示,泰国虎纹蛙的Shannon、Ace和Chao1指数均显著高于牛蛙 (P<0.05);门水平上,牛蛙的厚壁菌门丰度显著高于泰国虎纹蛙 (P<0.05),而放线菌门丰度显著低于后者 (P<0.05);属水平上,泰国虎纹蛙的乳球菌属 (Lactococcus) 和埃希杆菌-志贺菌属 (Escherichia-Shigella)丰度显著高于牛蛙 (P<0.05),而乳杆菌属 (Lactobacillus) 和明串珠菌属 (Leuconostoc) 丰度显著低于后者 (P<0.05)。研究表明,泰国虎纹蛙和牛蛙肌肉品质存在差异,但营养价值均较高;高通量测序分析显示两者肠道微生物群落组成存在差异。
Abstract:In order to investigate the muscle quality, intestinal morphology and microbial composition of Hoplobatrachus tigerinus and Lithobates catesbeiana, we monitored the muscle nutritional components of two cultured frogs species by national food safety standard measurement method, and evaluated their muscle quality in addition with analysis of muscle texture and microstructure. Besides, we applied high-throughput sequencing and histological section technology to analyze the microbial community composition and microstructure of their gut. The results show that there was no significant difference in the total amino acids between H. tigerinus and L. catesbeiana (P>0.05). The unsaturated fatty acids content of L. catesbeiana were significantly higher than those of H. tigerinus (P<0.05). In addition, the muscle hardness, chewiness and gumminess of L. catesbeiana were significantly higher than those of H. tigerinus (P<0.05). The analyses of intestinal microbial communities show that the Shannon, Ace and Chao1 indexes of H. tigerinus were significantly higher than those of L. catesbeiana (P<0.05). At phylum level, the abundance of Firmicutes in L. catesbeiana was significantly higher than that in H. tigerinus (P<0.05), while the abundance of Actinobacteriota was significantly lower (P<0.05); At genus level, the abundance of Lactococcus and Escherichia-Shigella in H. tigerinus was significantly higher than that in L. catesbeiana (P<0.05), while Lactobacillus and Leuconostoc were significantly lower (P<0.05). The results show that there are differences in muscle quality between H. tigerinus and L. catesbeiana, but both have high nutritional value; high-throughput sequencing analyses show that there are differences in the composition of intestinal microbial communities between the two species.
-
牛蛙 (Lithobates catesbeiana) 原产于北美洲,具有繁殖快、适应性强、生长迅速等优点,已成为我国主要养殖蛙类[1];泰国虎纹蛙 (Hoplobatrachus tigerinus) 是广东省近年来引进的优良水产养殖蛙类,其养殖投入低、养殖周期短且生长快,现已初步形成产业规模[2]。由于近几年养殖蛙市场需求量大,养殖户大多采用小池高密度养殖模式,且不同养殖品种均使用同一种 (同品牌) 商业人工配合饲料。同种人工饲料是否适宜不同养殖品种的生长和消化,相应的蛙产品肌肉品质是否能满足消费者需求以及如何科学开发饲料配方是养殖蛙产业研究的热点问题。肌肉品质是进入消费市场的重要评价指标,而肠道菌群一定程度上可反映养殖蛙的消化特性,这些基础数据对蛙类养殖和配合饲料的可持续开发至关重要。
肌肉品质包括营养价值和质构特性等[3],肌肉营养价值通常由蛋白质和氨基酸、脂肪和多不饱和脂肪酸的组成来作为评估指标[4],肌肉质构特性则由肌肉的硬度、黏性、弹性、咀嚼性、胶黏性和内聚性等质构参数来体现[3]。有关养殖蛙的肌肉营养价值,有学者针对黑斑蛙 (Pelophylax nigromaculatus)[5]、棘胸蛙 (Quasipaa spinosa)[6]、泰国虎纹蛙[7]、东北林蛙 (Rana dybowskii) 和牛蛙[8]肌肉营养组成开展了部分研究。然而,国内外有关养殖蛙类肌肉质构特性的研究报道较少,而肌肉质构是产品进入消费市场的重要评价指标。
肠道菌群与宿主的消化、营养吸收、代谢和生理功能密切相关[9]。目前,已报道肠道菌群的养殖蛙类有棘胸蛙[10]、黑斑蛙[11]、虎纹蛙 (H. chinensis)[12]等,但对泰国虎纹蛙和牛蛙肠道菌群的研究较少。开展人工养殖条件下泰国虎纹蛙和牛蛙肠道显微结构和微生物群落结构研究,有助于分析其对饲料的消化吸收及生长特性。本研究通过测定泰国虎纹蛙和牛蛙的肌肉营养组成和肌肉质构特性对其肌肉品质进行全面评价,通过高通量测序技术和组织切片技术分析两种养殖蛙的肠道微生物群落构成和显微结构,旨在全面了解泰国虎纹蛙和牛蛙的肌肉品质和肠道菌群差异,为养殖蛙类的开发利用及配合饲料的研制提供基础资料。
1. 材料与方法
1.1 样品采集
鲜活的成年雄性牛蛙和泰国虎纹蛙 (投喂同种膨化配合饲料) 取自广东省星蛙农业科技有限公司花都养殖基地。采用小池高密度养殖模式,养殖密度为140 只·m−2,养殖水位为5~10 cm,水温为 (28±2) ℃。配合饲料营养成分组成 (w):水分9.34%、粗蛋白质41.90%、粗脂肪5.78%、粗灰分9.48%。同一种蛙随机选择大小均一、形态相近的个体各6只,泰国虎纹蛙体质量为 (125.28±2.56) g,牛蛙体质量为 (151.87±3.15) g。毁髓处死后,每只蛙取1 cm大小的腿部肌肉组织,并于前肠相同肠道部位切取肠道组织各1 cm,用4% (φ) 多聚甲醛溶液固定,用于组织切片制作;将其余含有内容物的肠道组织迅速取出,液氮速冻后于−80 ℃保存,用于肠道菌群分析;沿着肌纤维剪取腿部肌肉组织,切成1 cm×1 cm×2 cm的长方体组织块,用于质构特性检测;腿部其余肌肉用于营养成分测定。每组做6个重复 (n=6)。
1.2 肌肉营养成分测定
按照鲜质量计算各营养成分的含量。参照GB 5009.5—2016 (凯氏定氮法)、GB 5009.6—2016 (索氏抽提法)、GB 5009.3—2016 (直接干燥法) 和GB 5009.4—2016 (第一法) 分别检测两种蛙肌肉的粗蛋白、粗脂肪、水分和灰分含量。使用全自动氨基酸分析仪参照GB 5009.124—2016测定氨基酸组成;由于采用酸水解处理样品,色氨酸被酸解,未测定。使用气相色谱仪参照GB 5009.168—2016分析脂肪酸种类和含量。根据联合国粮农组织/世界卫生组织 (FAO/WHO) 提出和1991年中国预防医学科学院提出的氨基酸评分模式,计算氨基酸评分 (Amino acid score, AAS)、化学评分 (Chemical score, CS) 和必需氨基酸指数 (Essential amino acid index, EAAI)[13]。
1.3 肌肉质构测定
将肌肉组织块放在CT3质构仪 (Brookfield公司,美国) 上,探头为P35,采用TPA模式进行测定,测前速度为2 mm·s−1,测试速度为1 mm·s−1,测后速度为5 mm·s−1,2次压缩的时间间隔为2 s,压缩比为25%,对肌肉质构特性进行测定。
1.4 组织切片观察
组织切片:对肌肉和肠道组织按照固定、脱水、透蜡、包埋、切片和染色的顺序进行操作,最终制作成组织切片。按照S=πr2计算肌纤维直径 (S为肌纤维横切面面积;r为肌纤维横切面半径;π为圆周率) ;并测量统计不同区域内肌纤维的数量和面积,计算肌纤维密度。每个样本对300条肌纤维进行统计。
1.5 肠道菌群高通量测序
按照EZNA® Soil DNA Kit试剂盒 (Omega Bio-tek) 说明书提取肠道菌群总DNA。DNA浓度和纯度使用NanoDrop2000检测。以正向引物338F (5'-ACTCCTACGGGAGGCAGCA-3') 和反向引物806R (5'-GGACTACHVGGGTWTCTAAT-3') 扩增16S rRNA基因V3—V4区。20 μL PCR反应体系为:10 μL 2×Pro Taq,正向引物和反向引物 (5 mmol·L−1) 各0.8 μL,10 ng DNA模板,ddH2O补足至20 μL。PCR过程:95 ℃ 3 min;95 ℃ 30 s,60 ℃ 30 s,72 ℃ 45 s,29个循环;最后72 ℃延伸10 min。利用AxyPrep DNA Gel Extraction Kit进行PCR产物纯化,使用Illumina测序专用的TruSeqTM DNA Sample Prep Kit试剂盒建立基因文库。构建好的文库通过Illumina Miseq PE300平台进行测序,由上海美吉生物医药科技有限公司完成。对获得的原始测序序列进行过滤、双端拼接,得到优化序列 (Tags);将优化序列进行聚类,划分操作分类单元 (Operational taxonomic units, OTU),并根据OTU的序列组成得到其物种分类。基于OTU分析结果,在美吉生物云平台 (https://cloud.majorbio.com) 上利用R语言 (version 3.3.1) 软件进行Alpha多样性指数分析、显著物种差异分析及功能差异预测分析。具体测定和分析方法参照Xia等[14]。
1.6 统计学分析
数据以“平均值±标准差 (
$\bar x $ ±s)”表示,采用SPSS 19.0软件在单因素方差分析 (One-way ANOVA) 基础上通过Duncan's多重比较法检验组间差异,显著性水平为P<0.05。2. 结果
2.1 常规营养成分
如表1所示,牛蛙的粗脂肪含量显著高于泰国虎纹蛙,而泰国虎纹蛙的灰分含量显著高于牛蛙 (P<0.05)。泰国虎纹蛙和牛蛙的水分含量和粗蛋白含量均无显著性差异 (P>0.05)。
表 1 泰国虎纹蛙和牛蛙每100 g肌肉常规营养成分比较Table 1. Comparison of conventional nutrients per 100 g of muscle between H. tigerinus and L. catesbeian g营养成分
Nutrientional composition泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeiana水分 Moisture 74.53±0.12 73.03±0.12 灰分 Crude ash 1.13±0.06b 1.03±0.06a 粗蛋白 Crude protein 23.63±0.15 24.00±0.36 粗脂肪 Crude lipid 0.70±0.00a 0.83±0.56b 注:同行不同字母表示组间差异显著 (P<0.05),下表同此。 Note: Values with different letters within the same row indicate significant differences (P<0.05), the same in the following tables. 2.2 氨基酸含量
在两种养殖蛙肌肉中均检测到16种氨基酸,其中必需氨基酸7种,非必需氨基酸9种 (表2)。两种蛙的甘氨酸含量均最高,其次是天门冬氨酸和异亮氨酸。除泰国虎纹蛙的苏氨酸含量显著高于牛蛙外 (P<0.05),其余氨基酸之间无显著性差异 (P>0.05)。两种蛙的必需氨基酸 (Essential amino acids, EAA)、非必需氨基酸 (Non-essential amino acids, NEAA)、呈味氨基酸 (Flavor amino acids, FAA)、支链氨基酸 (Branched chain amino acids, BCAA)、芳香氨基酸 (Aromatic amino acids, AAA) 与氨基酸总量 (Total amino acids, TAA) 均无显著性差异 (P>0.05)。
表 2 泰国虎纹蛙和牛蛙每100 g肌肉氨基酸比较Table 2. Comparison of amino acids per 100 g of muscle of H. tigerinus and L. catesbeiang 氨基酸
Amino acid泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeiana氨基酸
Amino acid泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeiana天门冬氨酸 Asp 2.57±0.06 2.55±0.08 亮氨酸 Leu 0.62±0.01 0.55±0.03 谷氨酸 Glu 1.18±0.02 1.17±0.44 赖氨酸 Lys 1.64±0.04 1.62±0.05 丝氨酸 Ser 1.16±0.02 1.18±0.04 脯氨酸 Pro 1.14±0.06 1.02±0.01 甘氨酸 Gly 3.98±0.12 3.98±0.13 氨基酸总量 TAA 24.50±0.52 24.10±0.79 苏氨酸 Thr 1.20±0.08b 1.03±0.01a 必需氨基酸 EAA 11.10±0.22 10.80±0.35 组氨酸 His 1.47±0.04 1.42±0.05 非必需氨基酸 NEAA 13.42±0.29 13.31±0.40 丙氨酸 Ala 1.23±0.02 1.23±0.38 呈味氨基酸 FAA 10.72±0.24 10.75±0.35 精氨酸 Arg 0.73±0.01 0.77±0.03 支链氨基酸 BCAA 5.02±0.12 4.94±0.20 酪氨酸 Tyr 1.21±0.04 1.19±0.04 芳香氨基酸 AAA 2.28±0.55 2.26±0.73 缬氨酸 Val 2.08±0.05 2.07±0.09 EAA/NEAA 82.69% 81.13% 蛋氨酸 Met 0.96±0.02 0.95±0.03 EAA/TAA 45.26% 44.75% 苯丙氨酸 Phe 1.07±0.02 1.07±0.04 FAA/TAA 43.71% 44.61% 异亮氨酸 Ile 2.33±0.07 2.31±0.09 BCAA/AAA 2.20 2.19 2.3 氨基酸营养品质评价
对泰国虎纹蛙和牛蛙肌肉必需氨基酸进行氨基酸评分(表3),两者除异亮氨酸的AAS (0.58) 外,其他均小于0.50;两者异亮氨酸的化学评分最高。
表 3 泰国虎纹蛙和牛蛙肌肉氨基酸营养价值评价Table 3. Evaluation of amino acids traits in muscles of H. tigerinus and L. catesbeiana项目
Item必需氨基酸
Essential amino acid泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeiana氨基酸评分
AAS异亮氨酸 Ile 0.58 0.58 亮氨酸 Leu 0.09 0.08 赖氨酸 Lys 0.30 0.30 苏氨酸 Thr 0.30 0.26 缬氨酸 Val 0.41 0.41 蛋氨酸+半胱氨酸
Met+Cys0.27 0.27 苯丙氨酸+酪氨酸
Phe+Tyr0.38 0.37 化学评分
CS异亮氨酸 Ile 0.43 0.43 亮氨酸 Leu 0.07 0.06 赖氨酸 Lys 0.23 0.23 苏氨酸 Thr 0.25 0.22 缬氨酸 Val 0.42 0.42 蛋氨酸+半胱氨酸
Met+Cys0.17 0.17 苯丙氨酸+酪氨酸
Phe+Tyr0.24 0.24 必需氨基酸
指数
EAAI22.76 21.81 2.4 脂肪酸含量
两种蛙的肌肉中均含有22种脂肪酸 (表4),包括饱和脂肪酸酸 (Saturated fatty acids, SFA) 8种,不饱和脂肪酸 (Unsaturated fatty acids, UFA) 14种;单不饱和脂肪酸 (Monounsaturated fatty acids, MUFA) 6种,多不饱和脂肪酸酸 (Polyunsaturated fatty acids, PUFA) 8种。从总体脂肪酸含量来看,泰国虎纹蛙为多不饱和脂肪酸>饱和脂肪酸>单不饱和脂肪酸,牛蛙为多不饱和脂肪酸>单不饱和脂肪酸>饱和脂肪酸,其中泰国虎纹蛙肌肉中饱和脂肪酸含量显著高于牛蛙 (P<0.05);牛蛙肌肉中单不饱和脂肪酸含量显著高于泰国虎纹蛙 (P<0.05),而多不饱和脂肪酸含量显著低于泰国虎纹蛙 (P<0.05)。
表 4 泰国虎纹蛙和牛蛙每100 g肌肉脂肪酸比较Table 4. Comparison of fatty acids in per 100 g muscles of H. tigerinus and L. catesbeianag 脂肪酸
Fatty acid泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeiana脂肪酸
Fatty acid泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeiana月桂 C12:0 0.03±0.00b 0.02±0.00a 二十碳三烯酸 C20:3 2.64±0.05 2.76±0.10 豆蔻 C14:0 0.38±0.03a 0.57±0.01b 二十碳四烯酸 C20:4 8.75±0.21b 5.27±0.05a 十五烷酸 C15:0 0.18±0.01a 0.22±0.01b 芥酸 C22:1 0.07±0.00a 0.10±0.00b 棕榈酸 C16:0 22.23±0.06b 19.57±0.15a 二十碳五烯酸 C20:5 1.72±0.05a 3.08±0.10b 十七烷酸 C17:0 0.29±0.00a 0.33±0.00b 二十二碳四烯酸 C22:4 0.55±0.02 0.64±0.06 硬脂酸 C18:0 7.42±0.36b 6.25±0.22a 二十四碳一烯酸 C24:1 0.85±0.03b 0.30±0.02a 花生酸 C20:0 0.09±0.00 0.09±0.01 二十二碳五烯酸 C24:1 2.30±0.06a 2.59±0.05b 二十三烷酸 C23:0 0.34±0.01a 0.57±0.01b 二十二碳六烯酸 (DHA) C22:6 6.53±0.11a 9.28±0.23b 棕桐油酸 C16:1 2.84±0.24a 5.15±0.15b 必需脂肪酸 EFA 25.71±0.36b 19.07±0.18a 油酸 C18:1 14.73±0.31a 22.20±0.17b 饱和脂肪酸 SFA 30.94±0.31b 27.61±0.35a 亚油酸 C18:2 24.40±0.30b 17.93±0.15a 不饱和脂肪酸 UFA 68.73±0.31a 72.30±0.23b 亚麻酸 C18:3 1.31±0.08b 1.14±0.03a 单不饱和脂肪酸 MUFA 21.38±0.38a 31.17±0.20b 二十碳一烯酸 C20:1 0.61±0.02a 0.94±0.06b 多不饱和脂肪酸 PUFA 47.6±0.14b 41.1±0.41a 二十碳二烯酸 C20:2 1.70±0.12b 1.02±0.02a PUFA/SFA (P/S) 1.53±0.12 1.49±0.33 泰国虎纹蛙的必需脂肪酸 (Essential fatty acids, EFA) 含量显著高于牛蛙 (P<0.05),其中,亚油酸和亚麻酸显著高于牛蛙 (P<0.05)。SFA中,除C20:0含量在两种蛙间不显著外 (P>0.05),其余各脂肪酸含量在两者之间均有显著性差异 (P<0.05)。UFA中,除C20:3和C22:4在两种蛙间不显著外 (P>0.05),其余各脂肪酸含量在两者间均有显著性差异 (P<0.05),此外,P/S介于1.49~1.53,符合WHO建议值 (P/S≥0.4)。
2.5 肌肉质构特性
如表5所示,牛蛙肌肉的硬度、咀嚼性、胶黏性均显著高于泰国虎纹蛙 (P<0.05),而黏性显著低于泰国虎纹蛙 (P<0.05)。此外,两者肌肉的弹性和内聚性无显著性差异 (P>0.05)。
表 5 泰国虎纹蛙和牛蛙肌肉质构特性的比较Table 5. Comparison of texture properties in muscle of H. tigerinus and L. catesbeiana指标
Index泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeiana硬度 Hardness/N 58.67±9.05a 90.37±15.76b 黏性Stickiness 4.91±1.17b 3.27±1.23a 弹性 Springiness/mm 0.65±0.08 0.66±0.12 咀嚼性 Chewiness/mJ 25.99±0.08a 43.24±12.87b 胶黏性 Gumminess/N 37.49±5.70a 66.43±15.12b 内聚性 Cohesiveness/% 0.73±0.46 0.76±0.07 2.6 肌肉和肠道形态结构比较
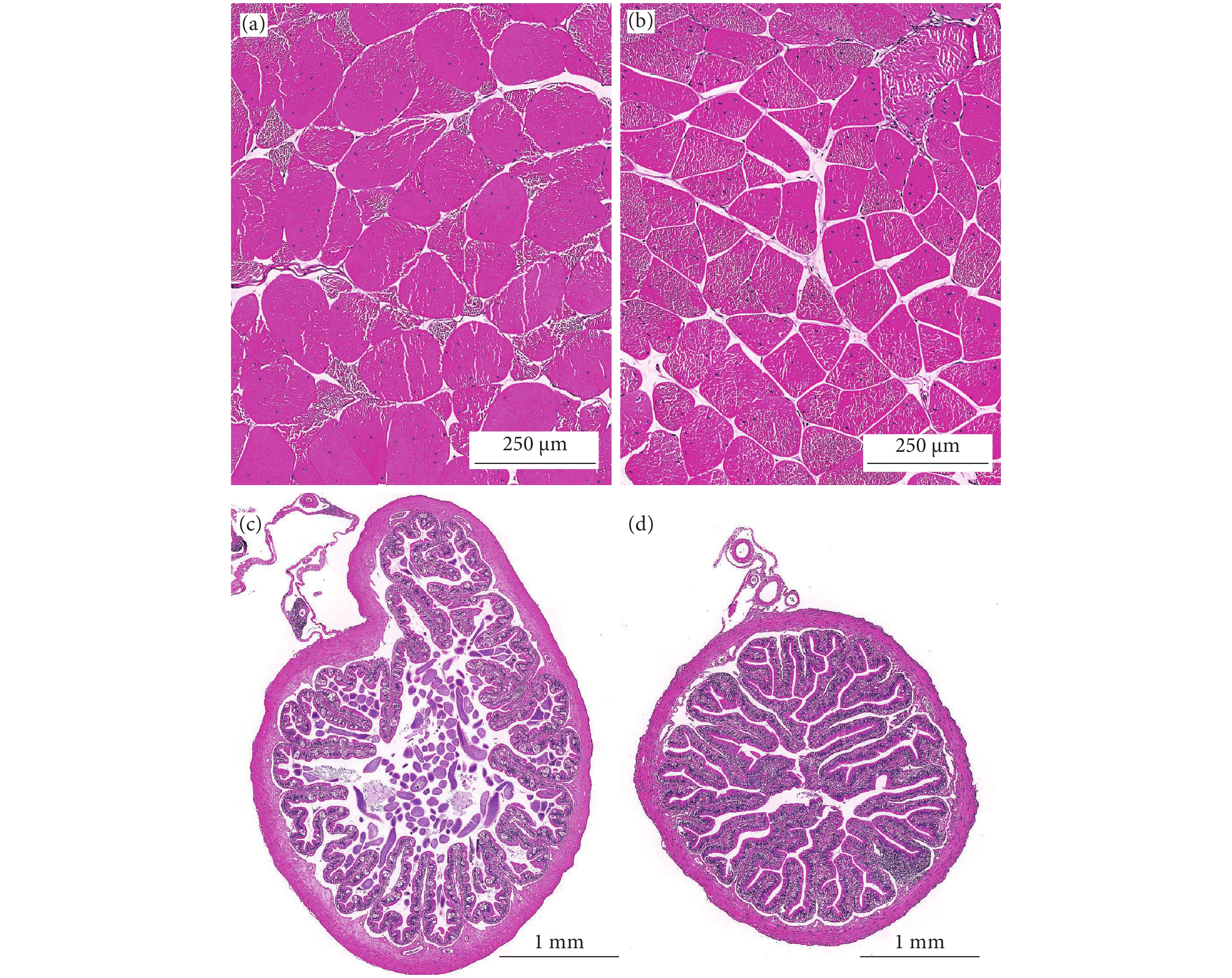
泰国虎纹蛙和牛蛙的肌肉和肠道形态结构如图1所示。两者肌纤维完整,排列规则,纤维大小与间隙均匀一致。牛蛙肌纤维直径显著大于泰国虎纹蛙 (P<0.05),而两者的肌纤维密度无显著性差异 (P>0.05,表6)。
![]() 图 1 泰国虎纹蛙和牛蛙肠道和肌肉的组织结构a. 泰国虎纹蛙肌肉切片;b. 牛蛙肌肉切片;c. 泰国虎纹蛙肠道切片;d. 牛蛙肠道切片。Figure 1. Tissue structures of intestine and muscle of two frogs speciesa. Muscle of H. tigerinus; b. Muscle of L. catesbeiana; c. Intestine of H. tigerinus; d. Intestine of L. catesbeiana.表 6 泰国虎纹蛙和牛蛙肌纤维和肠道组织结构比较Table 6. Comparison of muscle fibe and intestine morphology of H. tigerinus and L. catesbeiana
图 1 泰国虎纹蛙和牛蛙肠道和肌肉的组织结构a. 泰国虎纹蛙肌肉切片;b. 牛蛙肌肉切片;c. 泰国虎纹蛙肠道切片;d. 牛蛙肠道切片。Figure 1. Tissue structures of intestine and muscle of two frogs speciesa. Muscle of H. tigerinus; b. Muscle of L. catesbeiana; c. Intestine of H. tigerinus; d. Intestine of L. catesbeiana.表 6 泰国虎纹蛙和牛蛙肌纤维和肠道组织结构比较Table 6. Comparison of muscle fibe and intestine morphology of H. tigerinus and L. catesbeiana项目
Item泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeiana肌纤维密度
Density of muscle fibers/(根·mm−2)124.80±10.89 122.24±7.87 肌纤维直径
Diameter of muscle fibers/μm87.72±27.97a 94.10±18.02b 肠绒毛高度
Intestinal villus height/μm636.46±93.56a 676.54±111.15b 肠绒毛宽度
Intestinal villus width/μm188.69±30.86a 231.69±51.77b 肠肌层厚度
Intestinal muscular thickness/μm140.85±34.82b 124.01±15.16a 泰国虎纹蛙和牛蛙的肠道组织结构正常,肠壁较厚,结构完整性较好,肠绒毛密且粗壮,末端无破损,在两者的肠绒毛上皮细胞间均可观察到大量的杯状细胞。牛蛙的肠绒毛高度和宽度显著高于泰国虎纹蛙 (P<0.05),而肠肌层厚度则显著低于后者 (P<0.05)。
2.7 肠道微生物群落结构
2.7.1 Alpha多样性指数
高通量测序筛选后共获得945 594个有效序列,序列长度为201~451 bp,平均长度为426 bp。按照97%的序列相似性可聚类为618个 OTU。Alpha多样性分析显示,两种养殖蛙Good_coverage均高于0.99,说明样本中肠道微生物被充分检出,测序质量良好 (表7)。微生物群落多样性由Shannon和Simpson指数来反映,微生物群落丰富度由Ace和Chao1指数来反映。泰国虎纹蛙Shannon指数显著高于牛蛙 (P<0.01),表明泰国虎纹蛙肠道微生物群落多样性高于牛蛙。泰国虎纹蛙肠道的Ace和Chao1指数显著高于牛蛙 (P<0.05),表明泰国虎纹蛙肠道微生物的物种丰富度高于牛蛙。
表 7 Alpha多样性指数Table 7. Alpha diversity indices of six samples多样性指数
Diversity index泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeianaGood_coverage 0.999 5 0.999 7 Shannon 2.39±0.49b 1.13±0.20a Simpson 0.17±0.05a 0.50±0.10b Ace 219.42±82.25b 70.66±19.79a Chao1 213.39±80.07b 69.73±21.97a 2.7.2 OTU分布
两种养殖蛙之间有98个 (15.86%) 相同OTU,泰国虎纹蛙有487个 (78.80%) 独有OTU,牛蛙有33个 (5.34%) 独有OTU (图2)。泰国虎纹蛙的OTU数量显著高于牛蛙 (P<0.05),这与Shannon指数的结果一致。
![]() 图 2 OTU水平下两种养殖蛙肠道微生物的Veen图Figure 2. Veen diagrams of intestinal microbiota in H. tigerinus and L. catesbeiana at OTU level
图 2 OTU水平下两种养殖蛙肠道微生物的Veen图Figure 2. Veen diagrams of intestinal microbiota in H. tigerinus and L. catesbeiana at OTU level2.7.3 基于门水平肠道菌群分析
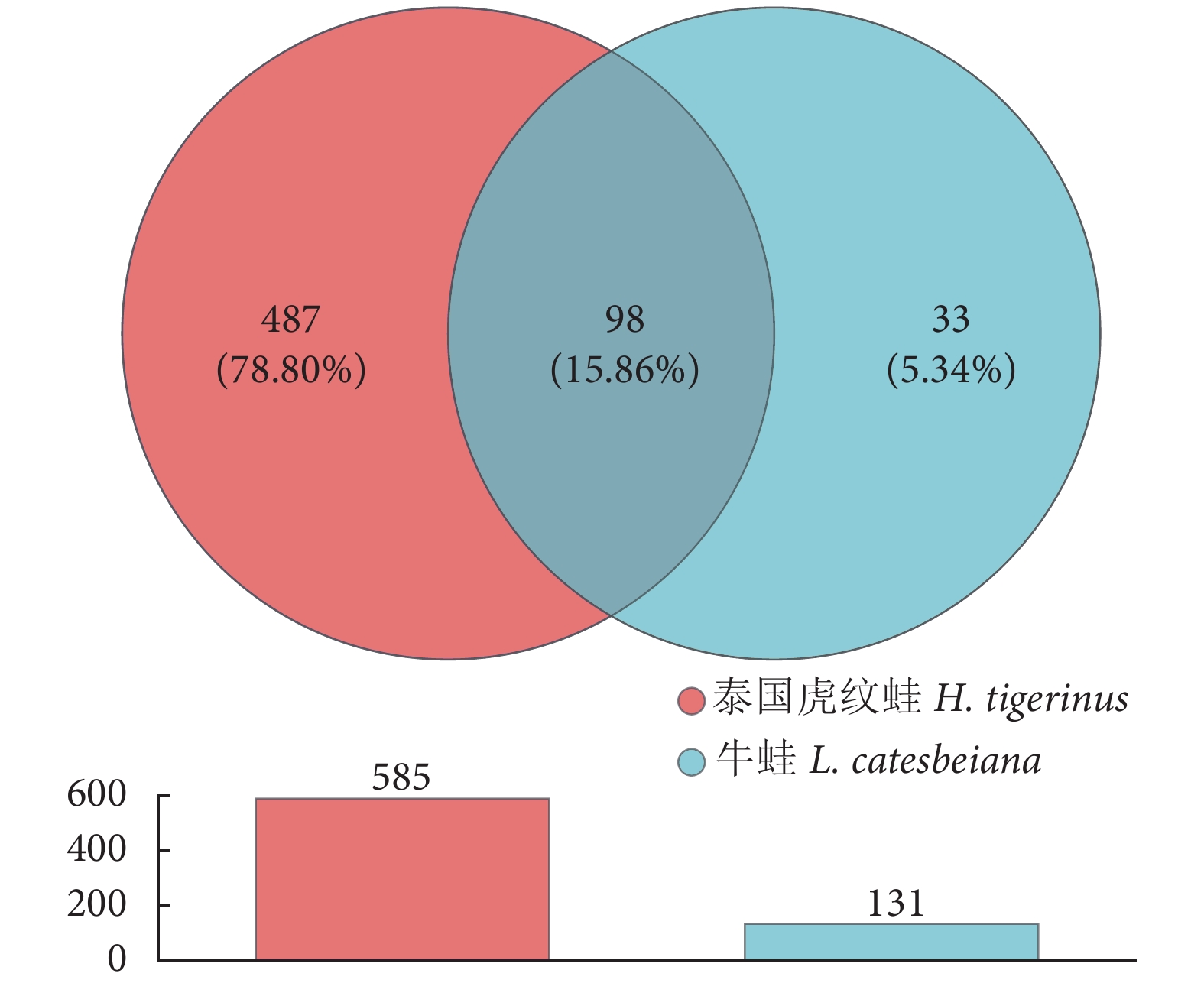
如图3所示,牛蛙肠道优势菌门是厚壁菌门 (92.26%) 和变形菌门 (7.36%)。泰国虎纹蛙优势菌门是厚壁菌门 (63.41%)、变形菌门 (19.43%)、梭杆菌门 (8.70%) 和拟杆菌门 (5.76%)。单因素方差分析显示,牛蛙肠道的厚壁菌门丰度显著高于泰国虎纹蛙 (P<0.05),而放线菌门丰度显著低于后者 (P<0.05)。
![]()
2.7.4 基于属水平肠道菌群分析
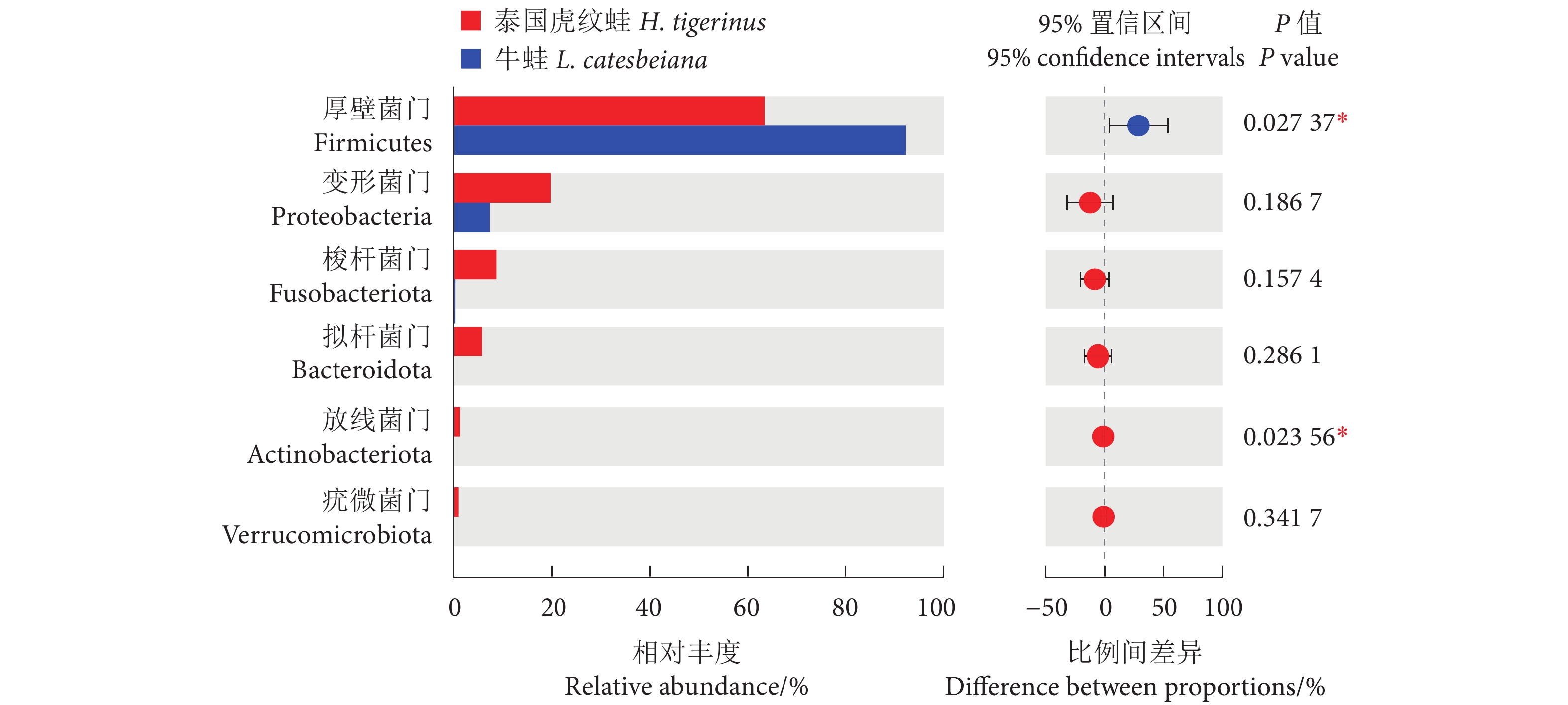
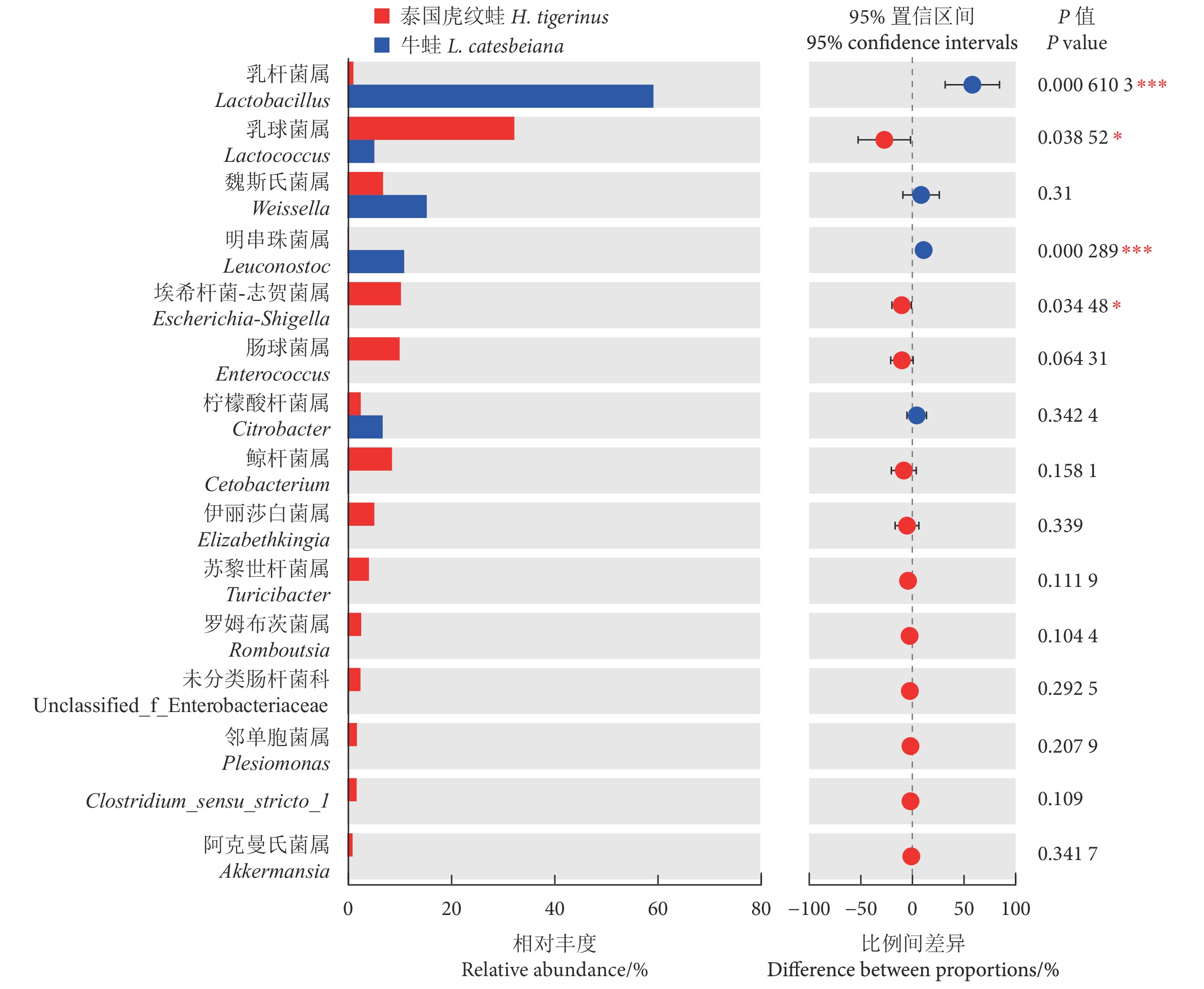
如图4所示,泰国虎纹蛙肠道菌群中丰度排名前5的菌属为乳球菌属 (Lactococcus, 32.34%)、埃希杆菌-志贺菌属 (Escherichia-Shigella, 10.36%)、肠球菌属 (Enterococcus, 10.11%)、鲸杆菌属 (Cetobacterium, 8.62%) 和魏斯氏菌属 (Weissella, 6.90%);牛蛙肠道优势菌属为乳杆菌属 (Lactobacillus, 59.30%)、魏斯氏菌属 (15.37%)、明串珠菌属 (Leuconostoc, 10.98%)、柠檬酸杆菌属 (Citrobacter, 6.82%) 和乳球菌属 (5.19%)。单因素方差分析显示,泰国虎纹蛙的乳球菌属和埃希杆菌-志贺菌属丰度显著高于牛蛙 (P<0.05),而乳杆菌属和明串珠菌属丰度显著低于后者 (P<0.05)。
2.7.5 肠道微生物功能预测
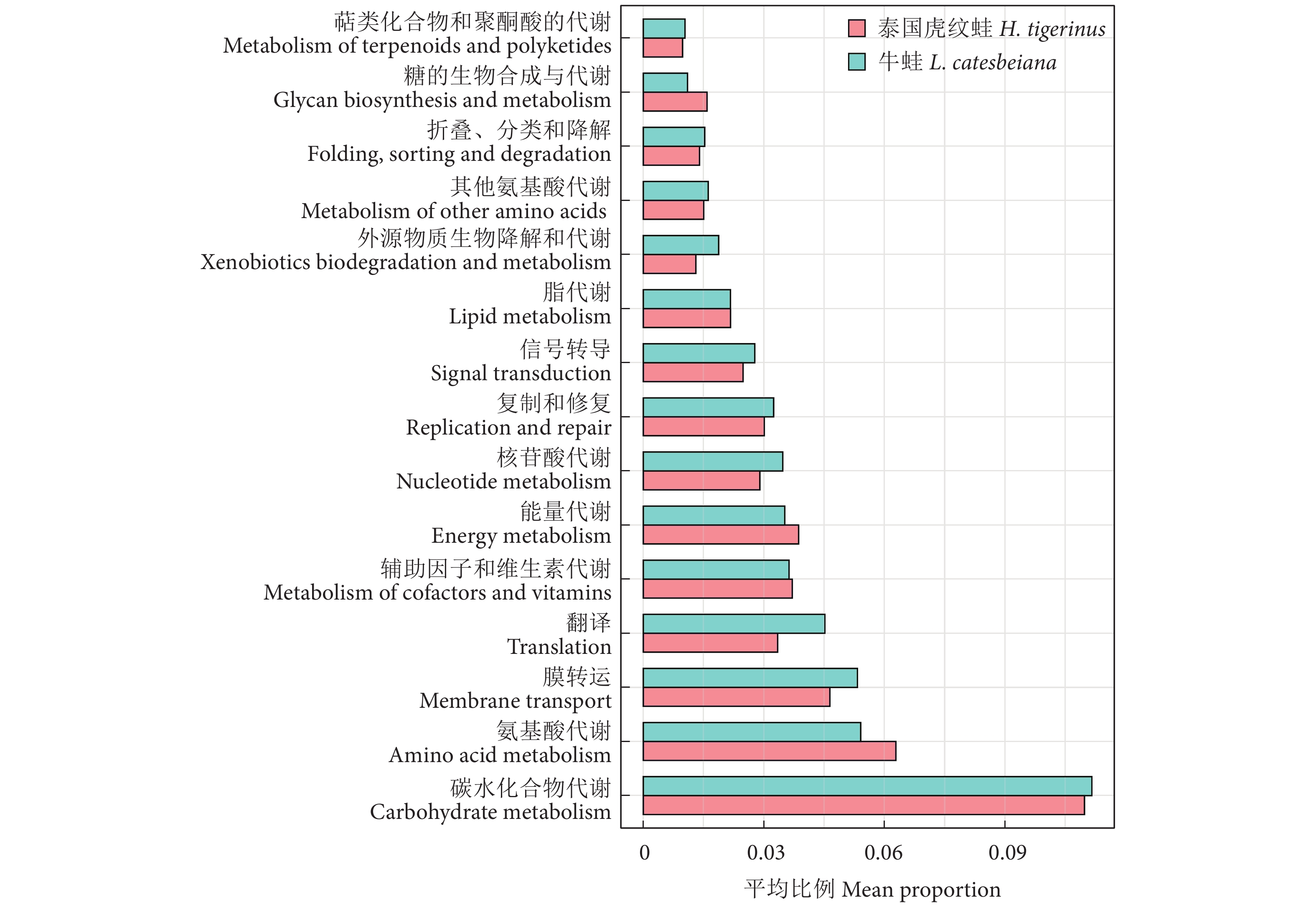
泰国虎纹蛙和牛蛙肠道菌群功能基因中占比较高的均为与新陈代谢相关的功能基因 (图5),包括碳水化合物代谢、氨基酸代谢、辅助因子和维生素代谢、能量代谢和脂代谢等新陈代谢功能基因,此外,膜转运和信号转导等功能基因也在两者肠道菌群中占有优势。功能预测显示泰国虎纹蛙和牛蛙肠道菌群可能主要参与宿主营养物质转运及代谢等相关进程。
![]() 图 5 泰国虎纹蛙和牛蛙肠道微生物功能预测Figure 5. Functional prediction of intestinal microbiota in H. tigerinus and L. catesbeiana
图 5 泰国虎纹蛙和牛蛙肠道微生物功能预测Figure 5. Functional prediction of intestinal microbiota in H. tigerinus and L. catesbeiana3. 讨论
3.1 肌肉常规营养成分
水分是肌肉含量最多的成分,一定程度上来说,肌肉水分充足则肉质更鲜嫩[8],这也是两种养殖蛙类口感鲜嫩多汁的主要原因。灰分是肌肉经高温处理后残留的无机物质,结果显示泰国虎纹蛙的灰分含量显著高于牛蛙。粗蛋白、粗脂肪水平是评价肌肉品质的重要指标,本研究中两种蛙的粗脂肪含量远低于15种常见的经济淡水鱼类[15],泰国虎纹蛙相比于牛蛙脂肪含量偏低,这可能是由于两者的规格不同。此外,与棘胸蛙[6]、黑斑蛙[10]和双团棘胸蛙 (Paa yunnanensis)[16]等养殖蛙类相比,泰国虎纹蛙和牛蛙的粗蛋白含量依然占据优势。泰国虎纹蛙和牛蛙肌肉营养总体呈现高蛋白低脂肪的特点,符合现代人们对高品质膳食的营养需求。
3.2 肌肉氨基酸组成
泰国虎纹蛙和牛蛙氨基酸总量中必需氨基酸的占比分别为45.26%和44.75%,必需氨基酸与非必需氨基酸的比值分别为82.69%和81.13%,优于FAO/WHO建议的理想模式40%和60%,因此在营养学上,泰国虎纹蛙和牛蛙肌肉均属于优质蛋白质来源。此外,泰国虎纹蛙和牛蛙的第一限制性氨基酸是亮氨酸,第二限制性氨基酸为蛋氨酸。
泰国虎纹蛙和牛蛙肌肉中必需氨基酸含量最丰富的是异亮氨酸,其属于人体必需氨基酸。异亮氨酸具有增强肠道黏膜免疫功能的作用[17],此外其通过调节机体对葡萄糖的摄取来调节肠道发育[18]。适量食用泰国虎纹蛙和牛蛙能够大量补充异亮氨酸,具有一定的保健功效。泰国虎纹蛙中苏氨酸含量显著高于牛蛙,苏氨酸是一种重要的营养强化剂,有缓解人体疲劳、促进生长发育的效果[19]。支链氨基酸具有促进蛋白质的合成、减缓衰老和防治肝肾功能衰竭的作用[20]。高支链氨基酸、低芳香氨基酸混合物对人体肝脏具有保护作用[21]。泰国虎纹蛙和牛蛙肌肉支链氨基酸与芳香氨基酸的比值基本一致,分别为2.20和2.19,均属于高支链氨基酸、低芳香氨基酸的食品。因此,食用泰国虎纹蛙和牛蛙肌肉可能有助于维持机体肝脏健康。肉类产品鲜美程度一般受呈味氨基酸含量和构成比例的影响,其中天冬氨酸、谷氨酸呈鲜味特性,甘氨酸、丙氨酸则呈甘味特性,且以谷氨酸的鲜味最强[22]。本研究中泰国虎纹蛙和牛蛙肌肉中含量最高的呈味氨基酸均为甘氨酸。泰国虎纹蛙和牛蛙肌肉中呈味氨基酸与总氨基酸的比值 (FAA/TAA) 较高,分别为43.71%和44.61%,这也是其味道鲜美的主要原因。
3.3 肌肉脂肪酸组成
泰国虎纹蛙和牛蛙肌肉中饱和脂肪酸以C16:0居多,单不饱和脂肪酸以C18:1居多,多不饱和脂肪酸以C18:2居多。两者饱和脂肪酸中含量最高的是C16:0,其次是C18:0;前者能使血清胆固醇降低,起到改善胰岛素抵抗的作用[23],后者可以减少肠道胆固醇吸收进而使肝脏胆固醇含量降低[24]。不饱和脂肪酸具有降血脂、预防心血管疾病和促进生长发育的功能[25],此外,不饱和脂肪酸含量对于肉质风味的形成具有重要作用[26],牛蛙的不饱和脂肪酸含量显著高于泰国虎纹蛙,说明其具有更好的风味和保健效果。单不饱和脂肪酸的适量摄入可以使低密度脂蛋白水平和冠心病发生率显著降低[27]。C18:1可降低血液总胆固醇含量,减少动脉粥样硬化与心脏病的患病风险[28]。牛蛙的单不饱和脂肪酸和C18:1含量显著高于泰国虎纹蛙,拥有更好的心脏保健效果。多不饱和脂肪酸含量与肉质加热过程中产生的香气成正比[29],一定程度上体现了肌肉的多汁性;多不饱和脂肪酸具有控制血压、血脂和抗肿瘤的作用[30],还具有延缓衰老等保健功效。泰国虎纹蛙的多不饱和脂肪酸含量显著高于牛蛙,其肌肉口感更具多汁性。C22:6 (DHA) 是人体必需脂肪酸,具有保护心肌及血管、治疗癌症、增强视力和健脑的功能[22]。牛蛙DHA含量比泰国虎纹蛙更高,因此从DHA含量角度来看,食用牛蛙的保健效果更佳。P/S是衡量脂肪酸营养价值的重要指标,WHO建议P/S应高于0.4[31]。泰国虎纹蛙和牛蛙的P/S介于1.49~1.53,超过一般的经济鱼类 (0.35~1.25)[32],说明泰国虎纹蛙和牛蛙是很好的脂肪酸营养食物源。可见,泰国虎纹蛙和牛蛙具有较高的脂肪酸营养价值,可对其营养和保健功效进行开发利用。
3.4 肌肉形态结构与质构特性
肌纤维是肌肉的基本组成单位,肌纤维直径会影响肌肉的质地和结构参数,如硬度、弹性和咀嚼性等[33],且肌纤维直径越大,肌肉硬度越高[34],而高弹性、高硬度的肌肉口感更佳[33]。肌肉形态学显示牛蛙肌纤维直径大于泰国虎纹蛙,这与肌肉质构特性的结果相一致。因此,从肌肉质构特性的角度看,牛蛙肌肉的食用口感更佳。
3.5 肠道形态结构和微生物多样性
肠绒毛高度、宽度和肌层厚度常用于评价肠道形态健康[35]。肠道绒毛的形态结构对其消化吸收营养物质的能力有着直接影响[36],肠绒毛的高度和宽度影响营养物质消化的速度;而食糜主要通过肠道肌层的收缩进行机械消化,因而肠肌层厚度的增加有利于机体对食糜的充分消化[37]。牛蛙肠绒毛高度和宽度均大于泰国虎纹蛙,表明牛蛙肠道消化吸收能力更优;而泰国虎纹蛙肠肌层厚度显著高于牛蛙,说明其对食糜的机械消化能力更胜一筹。
肠道微生物群落参与宿主的消化、代谢调节、免疫系统调控等过程。刘文舒等[12]发现虎纹蛙肠道优势菌门为厚壁菌门、变形菌门、拟杆菌门和梭杆菌门,与本研究结果相似。邵闯等[38]研究发现牛蛙肠道优势菌门为解糖微小寄生菌门、放线菌门及变形菌门,与本研究结果存在差异。以全豆粕蛋白为饲料来源的牛蛙,其肠道优势菌门为变形菌门、梭杆菌门和厚壁菌门[39],与本研究结果相似。因此,不同饲料可能会影响牛蛙肠道的优势菌。江昀等[40]推测蛙类肠道固有菌群为厚壁菌门、拟杆菌门和变形菌门,基因、年龄、性别以及养殖环境等多种因素会影响其组成比例。泰国虎纹蛙和牛蛙的厚壁菌门和放线菌门丰度差异显著,可能是由于两者基因不同对肠道优势菌产生了影响。
益生菌对维持肠道健康有重要作用,作为添加剂可以提高水产动物的生长率,增强其免疫力和抗病性,但是非宿主来源的益生菌在使用过程中存在潜在风险,宿主来源的益生菌更值得关注和挖掘[41]。乳球菌属[42]、乳杆菌属[43]、肠球菌属[44]、魏斯氏菌属[45]和明串珠菌属[46]是泰国虎纹蛙和牛蛙肠道菌群的优势菌属,均被证实是水产动物的潜在益生菌,对营养吸收、生长发育和抑制有害菌有积极作用;泰国虎纹蛙的乳球菌属丰度显著高于牛蛙,而乳杆菌属和明串珠菌属丰度则显著低于牛蛙,表明两者潜在的益生菌比例构成存在种间差异。鲸杆菌属能提高氨基酸转运和代谢率[47],泰国虎纹蛙中该菌属相对丰度最高,这与功能预测中氨基酸代谢的功能基因丰度的结果相一致。泰国虎纹蛙和牛蛙肠道菌群功能基因中占比较高的均为与新陈代谢相关的功能基因,这有利于提高宿主的生长发育、营养物质转运及代谢性能。
4. 结论
泰国虎纹蛙和牛蛙肉质总体呈现高蛋白、低脂肪的特点,具有高质量的必需氨基酸组成以及丰富的不饱和脂肪酸和必需脂肪酸;牛蛙的FAA/TAA和肌肉质构特性优于泰国虎纹蛙,具有更优质的风味和口感。高通量测序显示,泰国虎纹蛙的肠道菌群Alpha多样性更高;乳球菌属在泰国虎纹蛙肠道中显著富集,而乳杆菌属和明串珠菌属在牛蛙肠道中显著富集;功能预测显示两者肠道菌群可能主要参与宿主营养物质转运及代谢等相关进程。泰国虎纹蛙和牛蛙是平衡膳食营养结构的优质来源,具有良好的开发利用与人工养殖前景。
-
![]()
图 1 泰国虎纹蛙和牛蛙肠道和肌肉的组织结构
a. 泰国虎纹蛙肌肉切片;b. 牛蛙肌肉切片;c. 泰国虎纹蛙肠道切片;d. 牛蛙肠道切片。
Figure 1. Tissue structures of intestine and muscle of two frogs species
a. Muscle of H. tigerinus; b. Muscle of L. catesbeiana; c. Intestine of H. tigerinus; d. Intestine of L. catesbeiana.
![]()
图 2 OTU水平下两种养殖蛙肠道微生物的Veen图
Figure 2. Veen diagrams of intestinal microbiota in H. tigerinus and L. catesbeiana at OTU level
![]()
![]()
图 5 泰国虎纹蛙和牛蛙肠道微生物功能预测
Figure 5. Functional prediction of intestinal microbiota in H. tigerinus and L. catesbeiana
表 1 泰国虎纹蛙和牛蛙每100 g肌肉常规营养成分比较
Table 1 Comparison of conventional nutrients per 100 g of muscle between H. tigerinus and L. catesbeian g
营养成分
Nutrientional composition泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeiana水分 Moisture 74.53±0.12 73.03±0.12 灰分 Crude ash 1.13±0.06b 1.03±0.06a 粗蛋白 Crude protein 23.63±0.15 24.00±0.36 粗脂肪 Crude lipid 0.70±0.00a 0.83±0.56b 注:同行不同字母表示组间差异显著 (P<0.05),下表同此。 Note: Values with different letters within the same row indicate significant differences (P<0.05), the same in the following tables.  下载: 导出CSV
下载: 导出CSV
表 2 泰国虎纹蛙和牛蛙每100 g肌肉氨基酸比较
Table 2 Comparison of amino acids per 100 g of muscle of H. tigerinus and L. catesbeian
g 氨基酸
Amino acid泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeiana氨基酸
Amino acid泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeiana天门冬氨酸 Asp 2.57±0.06 2.55±0.08 亮氨酸 Leu 0.62±0.01 0.55±0.03 谷氨酸 Glu 1.18±0.02 1.17±0.44 赖氨酸 Lys 1.64±0.04 1.62±0.05 丝氨酸 Ser 1.16±0.02 1.18±0.04 脯氨酸 Pro 1.14±0.06 1.02±0.01 甘氨酸 Gly 3.98±0.12 3.98±0.13 氨基酸总量 TAA 24.50±0.52 24.10±0.79 苏氨酸 Thr 1.20±0.08b 1.03±0.01a 必需氨基酸 EAA 11.10±0.22 10.80±0.35 组氨酸 His 1.47±0.04 1.42±0.05 非必需氨基酸 NEAA 13.42±0.29 13.31±0.40 丙氨酸 Ala 1.23±0.02 1.23±0.38 呈味氨基酸 FAA 10.72±0.24 10.75±0.35 精氨酸 Arg 0.73±0.01 0.77±0.03 支链氨基酸 BCAA 5.02±0.12 4.94±0.20 酪氨酸 Tyr 1.21±0.04 1.19±0.04 芳香氨基酸 AAA 2.28±0.55 2.26±0.73 缬氨酸 Val 2.08±0.05 2.07±0.09 EAA/NEAA 82.69% 81.13% 蛋氨酸 Met 0.96±0.02 0.95±0.03 EAA/TAA 45.26% 44.75% 苯丙氨酸 Phe 1.07±0.02 1.07±0.04 FAA/TAA 43.71% 44.61% 异亮氨酸 Ile 2.33±0.07 2.31±0.09 BCAA/AAA 2.20 2.19
下载: 导出CSV
表 3 泰国虎纹蛙和牛蛙肌肉氨基酸营养价值评价
Table 3 Evaluation of amino acids traits in muscles of H. tigerinus and L. catesbeiana
项目
Item必需氨基酸
Essential amino acid泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeiana氨基酸评分
AAS异亮氨酸 Ile 0.58 0.58 亮氨酸 Leu 0.09 0.08 赖氨酸 Lys 0.30 0.30 苏氨酸 Thr 0.30 0.26 缬氨酸 Val 0.41 0.41 蛋氨酸+半胱氨酸
Met+Cys0.27 0.27 苯丙氨酸+酪氨酸
Phe+Tyr0.38 0.37 化学评分
CS异亮氨酸 Ile 0.43 0.43 亮氨酸 Leu 0.07 0.06 赖氨酸 Lys 0.23 0.23 苏氨酸 Thr 0.25 0.22 缬氨酸 Val 0.42 0.42 蛋氨酸+半胱氨酸
Met+Cys0.17 0.17 苯丙氨酸+酪氨酸
Phe+Tyr0.24 0.24 必需氨基酸
指数
EAAI22.76 21.81
下载: 导出CSV
表 4 泰国虎纹蛙和牛蛙每100 g肌肉脂肪酸比较
Table 4 Comparison of fatty acids in per 100 g muscles of H. tigerinus and L. catesbeiana
g 脂肪酸
Fatty acid泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeiana脂肪酸
Fatty acid泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeiana月桂 C12:0 0.03±0.00b 0.02±0.00a 二十碳三烯酸 C20:3 2.64±0.05 2.76±0.10 豆蔻 C14:0 0.38±0.03a 0.57±0.01b 二十碳四烯酸 C20:4 8.75±0.21b 5.27±0.05a 十五烷酸 C15:0 0.18±0.01a 0.22±0.01b 芥酸 C22:1 0.07±0.00a 0.10±0.00b 棕榈酸 C16:0 22.23±0.06b 19.57±0.15a 二十碳五烯酸 C20:5 1.72±0.05a 3.08±0.10b 十七烷酸 C17:0 0.29±0.00a 0.33±0.00b 二十二碳四烯酸 C22:4 0.55±0.02 0.64±0.06 硬脂酸 C18:0 7.42±0.36b 6.25±0.22a 二十四碳一烯酸 C24:1 0.85±0.03b 0.30±0.02a 花生酸 C20:0 0.09±0.00 0.09±0.01 二十二碳五烯酸 C24:1 2.30±0.06a 2.59±0.05b 二十三烷酸 C23:0 0.34±0.01a 0.57±0.01b 二十二碳六烯酸 (DHA) C22:6 6.53±0.11a 9.28±0.23b 棕桐油酸 C16:1 2.84±0.24a 5.15±0.15b 必需脂肪酸 EFA 25.71±0.36b 19.07±0.18a 油酸 C18:1 14.73±0.31a 22.20±0.17b 饱和脂肪酸 SFA 30.94±0.31b 27.61±0.35a 亚油酸 C18:2 24.40±0.30b 17.93±0.15a 不饱和脂肪酸 UFA 68.73±0.31a 72.30±0.23b 亚麻酸 C18:3 1.31±0.08b 1.14±0.03a 单不饱和脂肪酸 MUFA 21.38±0.38a 31.17±0.20b 二十碳一烯酸 C20:1 0.61±0.02a 0.94±0.06b 多不饱和脂肪酸 PUFA 47.6±0.14b 41.1±0.41a 二十碳二烯酸 C20:2 1.70±0.12b 1.02±0.02a PUFA/SFA (P/S) 1.53±0.12 1.49±0.33
下载: 导出CSV
表 5 泰国虎纹蛙和牛蛙肌肉质构特性的比较
Table 5 Comparison of texture properties in muscle of H. tigerinus and L. catesbeiana
指标
Index泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeiana硬度 Hardness/N 58.67±9.05a 90.37±15.76b 黏性Stickiness 4.91±1.17b 3.27±1.23a 弹性 Springiness/mm 0.65±0.08 0.66±0.12 咀嚼性 Chewiness/mJ 25.99±0.08a 43.24±12.87b 胶黏性 Gumminess/N 37.49±5.70a 66.43±15.12b 内聚性 Cohesiveness/% 0.73±0.46 0.76±0.07
下载: 导出CSV
表 6 泰国虎纹蛙和牛蛙肌纤维和肠道组织结构比较
Table 6 Comparison of muscle fibe and intestine morphology of H. tigerinus and L. catesbeiana
项目
Item泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeiana肌纤维密度
Density of muscle fibers/(根·mm−2)124.80±10.89 122.24±7.87 肌纤维直径
Diameter of muscle fibers/μm87.72±27.97a 94.10±18.02b 肠绒毛高度
Intestinal villus height/μm636.46±93.56a 676.54±111.15b 肠绒毛宽度
Intestinal villus width/μm188.69±30.86a 231.69±51.77b 肠肌层厚度
Intestinal muscular thickness/μm140.85±34.82b 124.01±15.16a
下载: 导出CSV
表 7 Alpha多样性指数
Table 7 Alpha diversity indices of six samples
多样性指数
Diversity index泰国虎纹蛙
H. tigerinus牛蛙
L. catesbeianaGood_coverage 0.999 5 0.999 7 Shannon 2.39±0.49b 1.13±0.20a Simpson 0.17±0.05a 0.50±0.10b Ace 219.42±82.25b 70.66±19.79a Chao1 213.39±80.07b 69.73±21.97a
下载: 导出CSV
-
[1] 方卫东, 鲁康乐, 张春晓, 等. 豆粕替代鱼粉对牛蛙生长、体组成、消化酶活力及肝脏生化指标的影响[J]. 水产学报, 2016, 40(11): 1742-1752. [2] 刘世伟. 稻蛙共作对稻田土壤生态及水稻生长、产量与品质性状的影响[D]. 广州: 华南农业大学, 2019: 8. [3] 牛树辉, 李红燕, 潘厚军, 等. 鲜活饵料和人工配合饲料对鳜肌肉营养成分和质构特性的影响[J]. 水生生物学报, 2023, 47(1): 37-44. [4] JIANG W D, WU P, TANG R J, et al. Nutritive values, flavor amino acids, healthcare fatty acids and flesh quality improved by manganese referring to up-regulating the antioxidant capacity and signaling molecules TOR and Nrf2 in the muscle of fish[J]. Food Res Int, 2016, 89: 670-678. doi: 10.1016/j.foodres.2016.09.020
[5] 何志刚, 王冬武, 徐永福, 等. 黑斑蛙肌肉营养成分分析及评价[J]. 中国饲料, 2018(17): 74-77. [6] 刘芳彬, 王伟, 黄永春, 等. 棘胸蛙肌肉营养成分季节变化分析与评价[J]. 食品工业科技, 2022, 43(9): 365-371. [7] 刘丽, 刘楚吾, 林东年, 等. 泰国虎纹蛙与中国虎纹蛙肌肉的营养成分比较[J]. 水利渔业, 2008(3): 64-66. [8] 蓝蔚青, 杜金涛, 刘大勇, 等. 3种养殖蛙类肌肉基本营养成分分析[J]. 广东海洋大学学报, 2022, 42(2): 142-147. [9] WANG A R, RAN C, RINGO E, et al. Progress in fish gastrointestinal microbiota research[J]. Rev Aquac, 2018, 10(3): 626-640. doi: 10.1111/raq.12191
[10] 何志刚, 伍远安, 徐永福, 等. 野生与养殖黑斑蛙肌肉营养品质的比较分析[J]. 水产科学, 2019, 38(4): 506-513. [11] 刘文舒, 郭小泽, 陈彦良, 等. 养殖黑斑蛙肠道微生物结构与功能分析[J]. 经济动物学报, 2021, 25(2): 68-76. [12] 刘文舒, 陈彦良, 郭小泽, 等. 虎纹蛙肠道微生物结构与功能分析[J]. 野生动物学报, 2020, 41(1): 171-181. [13] 刘亚秋, 刘明典, 李新辉, 等. 3个广东鲂地理种群肌肉营养成分及能量密度比较研究[J]. 南方水产科学, 2022, 18(4): 163-169. [14] XIA Y, YU E M, LU M X, et al. Effects of probiotic supplementation on gut microbiota as well as metabolite profiles within Nile tilapia, Oreochromis niloticus[J]. Aquaculture, 2020, 527: 735428. doi: 10.1016/j.aquaculture.2020.735428
[15] 韩迎雪, 林婉玲, 杨少玲, 等. 15种淡水鱼肌肉脂肪含量及脂肪酸组成分析[J]. 食品工业科技, 2018, 39(20): 217-222. [16] 陈明勇, 彭剑林, 李灿鹏. 双团棘胸蛙(Paa yunnanensis)肌肉营养成分分析与评价[J]. 云南大学学报(自然科学版), 2010, 32(1): 114-117. [17] 李晋南, 王良, 王连生. 水产动物异亮氨酸营养研究进展[J]. 动物营养学报, 2022, 34(3): 1374-1387. doi: 10.3969/j.issn.1006-267x.2022.03.002 [18] ZHANG S H, YANG Q, REN M, et al. Effects of isoleucine on glucose uptake through the enhancement of muscular membrane concentrations of GLUT1 and GLUT4 and intestinal membrane concentrations of Na+/glucose co-transporter 1 (SGLT-1) and GLUT2[J]. Br J Nutr, 2016, 116(4): 593-602. doi: 10.1017/S0007114516002439
[19] 魏佳, 王壮壮, 于海波, 等. 产L-苏氨酸重组大肠杆菌的构建和发酵性能[J]. 微生物学通报, 2019, 46(4): 695-706. [20] 王莉苹, 陈飞, 韩雨哲, 等. 支链氨基酸(BCAA)及水产动物对其需求量的研究进展[J]. 饲料工业, 2015, 36(14): 35-40. [21] 赵亭亭, 陈超, 邵彦翔. 雌雄条纹锯鮨肌肉营养成分的比较与评价[J]. 渔业科学进展, 2019, 40(3): 151-159. [22] 卢文麒, 许琪娅, 蔡国鹤, 等. 不同规格花鳗鲡肌肉营养成分比较分析[J]. 饲料工业, 2023, 44(14): 80-86. doi: 10.13302/j.cnki.fi.2023.14.013 [23] 张茜, 王瑞, 于智超, 等. 棕榈酸诱导胰岛β细胞氧化应激和内质网应激的研究进展[J]. 激光生物学报, 2023, 32(2): 118-125. [24] 徐庆, 张新胜, 张永, 等. 辛酸、癸酸和硬脂酸对ApoE~(-/-)小鼠肠道中外源性胆固醇吸收的影响[J]. 卫生研究, 2018, 47(1): 119-122, 127. [25] SIRI-TARINO P W, CHIU S, BERGERON N, et al. Saturated fats versus polyunsaturated fats versus carbohydrates for cardiovascular disease prevention and treatment[J]. Annu Rev Nutr, 2015, 35(1): 517-543. doi: 10.1146/annurev-nutr-071714-034449
[26] 张瑞, 白云鹏, 贾莉, 等. 牛至精油对平凉红牛半腱肌肉品质、脂肪酸及挥发性风味物质的影响[J]. 动物营养学报, 2022, 34(7): 4452-4463. [27] 梁琍, 桂庆平, 冉辉, 等. 野生与养殖黄颡鱼鱼卵的营养成分比较[J]. 水产科学, 2016, 35(5): 522-527. [28] 胡磊, 谢庆超, 潘迎捷, 等. 蒸制和煮制对中华绒螯蟹中胆固醇和脂肪酸含量的影响[J]. 上海海洋大学学报, 2023, 32(3): 649-659. [29] 吴娜. 基于脂质热氧化降解解析中华绒螯蟹关键香气物质的形成机制[D]. 上海: 上海海洋大学, 2017: 44-73. [30] 周礼敬, 沈东霞, 詹会祥. 鱼类肌肉营养成分与人体健康研究[J]. 畜牧与饲料科学, 2013, 34(5): 69-71. [31] 郭俊涛, 谢遇春, 苏馨, 等. 大足黑山羊不同部位肌肉脂肪酸比较研究[J]. 肉类研究, 2020, 34(9): 6-11. [32] 杜宁宁, 黄晓丽, 郝其睿, 等. 八种鲑科鱼类肌肉营养成分分析与营养价值评价[J]. 水产学杂志, 2022, 35(4): 9-18, 37. [33] 董立学, 喻亚丽, 毛涛, 等. 池塘内循环流水养殖斑点叉尾鮰肌肉品质的分析[J]. 中国水产科学, 2021, 28(7): 914-924. [34] CHENG J H, SUN D W, HAN Z, et al. Texture and structure measurements and analyses for evaluation of fish and fillet freshness quality: a review[J]. Comp Rev Food Sci Food Saf, 2014, 13(1): 52-61. doi: 10.1111/1541-4337.12043
[35] KUEBUTORNYE F K A, WANG Z, LU Y, et al. Effects of three host-associated Bacillus species on mucosal immunity and gut health of Nile tilapia, Oreochromis niloticus and its resistance against Aeromonas hydrophila infection[J]. Fish Shellfish Immunol, 2020, 97: 83-95. doi: 10.1016/j.fsi.2019.12.046
[36] KHOSRAVI S, RAHIMNEJAD S, HERAULT M, et al. Effects of protein hydrolysates supplementation in low fish meal diets on growth performance, innate immunity and disease resistance of red sea bream Pagrus major[J]. Fish Shellfish Immunol, 2015, 45(2): 858-868. doi: 10.1016/j.fsi.2015.05.039
[37] 林宜锦, 杨欢, 林鹏志, 等. 饵料中添加肉桂醛对日本鳗鲡肠道结构的影响[J]. 饲料研究, 2022, 45(11): 52-55. [38] 邵闯, 秦守旺, 朱波, 等. 投喂策略对牛蛙蝌蚪生长发育及肝肠功能的影响[J]. 水产学报, 2023, 47(10): 72-83. [39] 王健, 王玲, 鲁康乐, 等. 摄食全豆粕蛋白饲料的牛蛙肠道微生物组成[J]. 集美大学学报(自然科学版), 2018, 23(4): 241-248. [40] 江昀, 胡亚洲, 向建国, 等. 不同健康状态的棘胸蛙肠道菌群结构分析[J]. 水生生物学报, 2022, 46(9): 1332-1340. [41] LIU Z, LIU W S, RAN C, et al. Abrupt suspension of probiotics administration may increase host pathogen susceptibility by inducing gut dysbiosis[J]. Sci Rep, 2016, 6(1): 23214. doi: 10.1038/srep23214
[42] 张美玲, 单承杰, 杜震宇. 益生菌与鱼类肠道健康研究进展[J]. 水产学报, 2021, 45(1): 147-157. [43] 包瑞璇, 周皓隆, 方再郗, 等. 大菱鲆肠道乳杆菌Lactobacillus sp. N5的筛选鉴定及其抑制水产病原菌的研究[J]. 水产学杂志, 2023, 36(3): 38-45. [44] 雷喜玲, 叶金云, 陈丽仙, 等. 屎肠球菌的作用机制及其在水产动物中的应用[J]. 饲料工业, 2022, 43(22): 9-15. [45] 王腾腾. 一株许氏平鲉肠道乳酸菌的分离鉴定、特性分析及其应用研究[D]. 上海: 上海海洋大学, 2017: 31-37. [46] 韩琳. 一例日本对虾暴发性死亡的病原分析及益生菌联用对对虾免疫增强作用[D]. 上海: 上海海洋大学, 2017: 21-36. [47] WANG L, WANG J, LU K L, et al. Total replacement of fish meal with soybean meal in diets for bullfrog (Lithobates catesbeianus): effects on growth performance and gut microbial composition[J]. Aquaculture, 2020, 524: 735236. doi: 10.1016/j.aquaculture.2020.735236
-
期刊类型引用(1)
1. 高祥,崔清明,刘莹莹,任慧波,邓缘,朱吉,李志强,杨仕柳,陈晨,彭英林. 乡草花猪种质特性研究. 中国畜牧杂志. 2024(08): 199-204 .  百度学术
百度学术
其他类型引用(0)
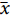
计量
- 文章访问数: 695
- HTML全文浏览量: 45
- PDF下载量: 52
- 被引次数: 1
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
