
Effects and potential mechanism of oyster peptide on paroxetine-induced sexual dysfunction in male mice
-
摘要:
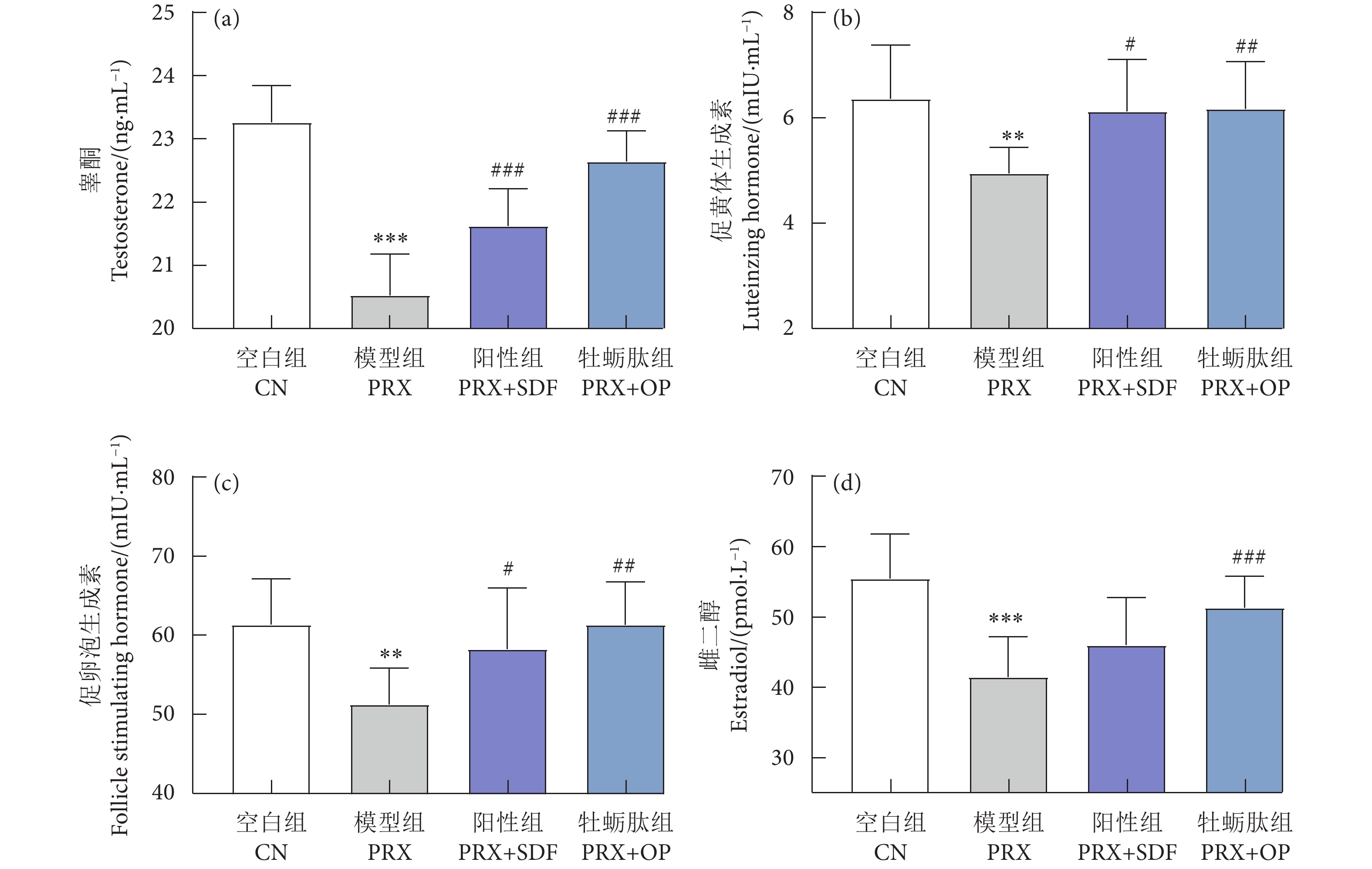
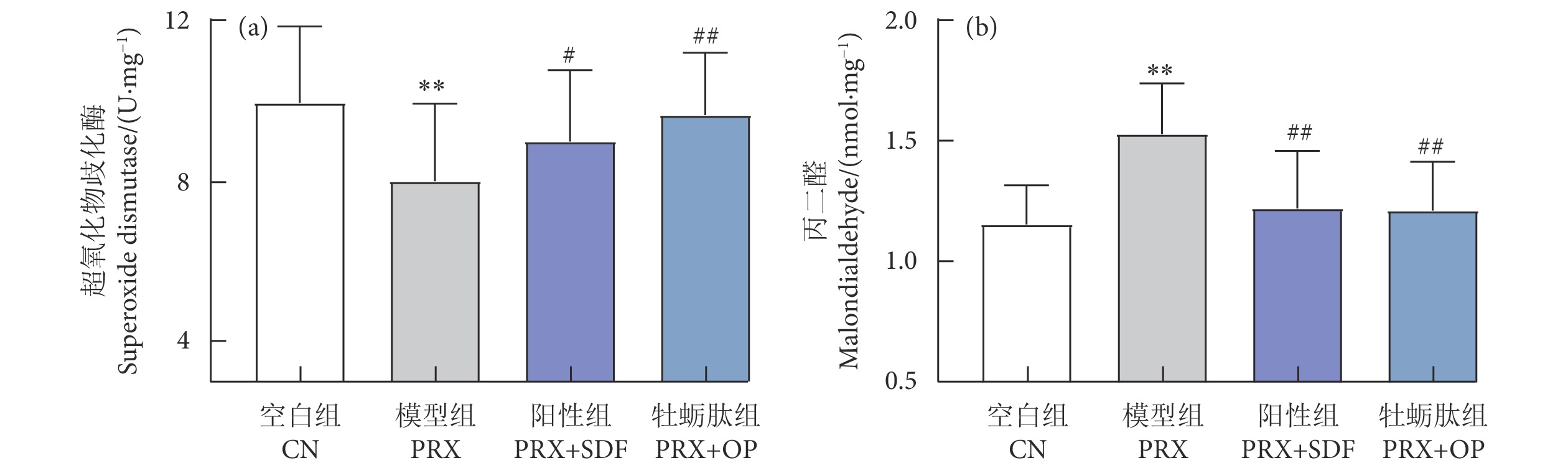
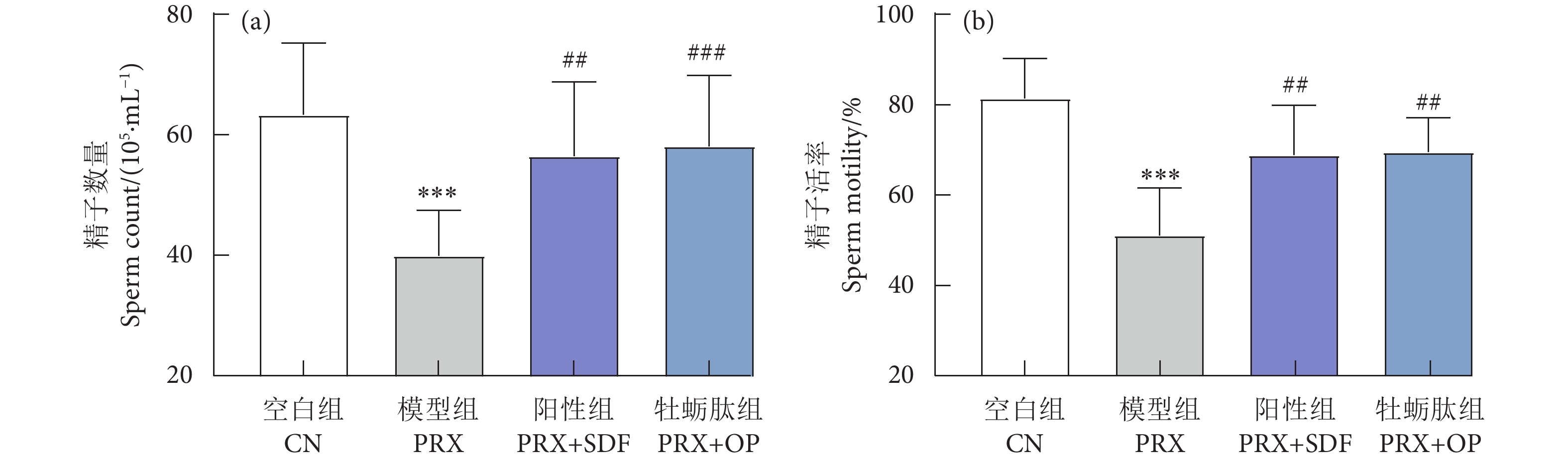
牡蛎肽 (Oyster peptide, OP) 具有多种生物活性,然而,其对男性性功能障碍的作用效果仍知之甚少。以牡蛎肽为研究对象,探讨其对男性性功能障碍的作用效果及其潜在机制。每天灌胃帕罗西汀 (Paroxetine, PRX) 构建雄性小鼠性功能障碍模型,同时灌胃牡蛎肽 (500 mg·kg−1),持续28 d。结果表明:与模型 (PRX) 组小鼠相比,牡蛎肽可显著提高雄性小鼠的性能力 (P<0.05),恢复血清性激素水平 (P<0.01),提高阴茎组织一氧化氮 (NO) 含量 (P<0.01)、环磷酸鸟苷 (cGMP) 含量 (P<0.05), 和一氧化氮合酶 (NOS) 活性 (P<0.05),并降低磷酸二酯酶-5 (PDE-5) 活性 (P<0.01);同时,牡蛎肽可增强睾丸标志性酶活性 (P<0.05) 和抗氧化能力 (P<0.01),改善精子质量。此外,HE染色结果显示:牡蛎肽可恢复小鼠睾丸生精小管内生精细胞的数量与形态,减少生精小管空泡化现象。综上所述,牡蛎肽可有效减缓PRX导致的雄性小鼠性功能障碍,推测其对男性性功能障碍具有潜在的保护作用。
Abstract:Oyster peptide (OP) has various biological activities. However, its effects on male sexual dysfunction is still poorly understood. In this study, we explored its effects and potential mechanism on male sexual dysfunction. Besides, we established a paroxetine (PRX)-induced sexual dysfunction model by gavaging OP (500 mg·kg− 1) in mice for 28 d. The results show that compared with the model (PRX) group, OP could improve the sexual performance of male mice (P<0.05), restored serum sex hormone levels (P<0.01), increased penile tissue nitric oxide (NO) content (P<0.01), cyclic guanosine monophosphate (cGMP) content (P<0.05) and nitric oxide synthase (NOS) activity (P<0.05), and decreased phosphodiesterase-5 (PDE-5) activity (P<0.01). Meanwhile, OP enhanced testicular marker enzymes activities (P<0.05) and antioxidant capacity (P<0.01), and improved sperm quality. In addition, HE staining results show that OP could restore the number and morphology of spermatogenic cells in seminiferous tubules of mice, and reduced the vacuolization of seminiferous tubules. In conclusion, OP can alleviate PRX-induced sexual dysfunction effectively in male mice and has a potential protective effect on male sexual dysfunction.
-
Keywords:
- Oyster peptide /
- Paroxetine /
- Sexual dysfunction /
- Erectile dysfunction /
- Sexual behavior
-
三疣梭子蟹 (Portunus trituberculatus),俗称梭子蟹,属于甲壳纲、十足目、梭子蟹科、梭子蟹属,其肉质鲜美、营养丰富,广受消费者青睐,是中国沿海重要的经济蟹类[1]。梭子蟹生长速度快、养殖成本低、经济效益高,已成为中国沿海地区的重要养殖品种。由于雌、雄梭子蟹的营养成分不同,在市场销售前需要对其性别进行分类。目前,梭子蟹的性别分类主要靠人眼识别和手工挑选,效率低下。因此,开发一种计算机辅助自动分类系统来区分梭子蟹的性别非常必要。
早期的分类任务主要是手工设计提取特征,包括传统机器学习[2-4]和利用上下文信息[5-6],但是该类方法的表达能力较弱,所以分类模型的泛化能力不强。例如,Lecun等[7]提出的一种多层人工神经网络LeNet是为手写数字分类而设计的卷积神经网络 (Convolutional neural network, CNN)。2006年,深度学习 (Deep learning) [8]的概念被提出,在深度学习技术的推动下,图像分类及识别的相关研究迅速发展[9-13]。
基于深度学习的图像分类中,Krizhevsky等[9]构建了一种较深层数的网络AlexNet,并首次引入了Relu激活函数,同时在全连接层中使用Dropout,解决了模型的过拟合问题。牛津大学的几何视觉组 (Visual geometry group[10])设计的VGG模型在网络结构上并没有太大的创新,但是通过实验对比发现,增加网络的层数确实能够在一定程度上提高网络的训练效果。此外,VGG模型始终使用的是非常小的卷积核,通过串联很多小的卷积核后,其感受野和一个大卷积核相同,因此能很大程度地减少模型训练所需的参数。He等[12]开发出一种具有深度残差结构的卷积神经网络ResNet,很好地解决了网络深度达到一定层数时,模型的性能逐渐会趋向于饱和的问题,以及在网络达到某一深层时会使模型的性能急剧下降的问题。
在过去的几年里,图像的成熟分类技术主要得益于两个关键因素:一个是卷积神经网络,另一个是大量的可用图像数据集,如CIFAR数据集[14]和ImageNet 数据集[15]。当前,一些研究者已经将图像分类方法应用到实际场景中。例如,在医学图像分类领域中,杜丽君等[16]提出了一种基于注意力机制和多任务学习的阿尔兹海默症分类方法。该方法是一种不需要人工提取特征的网络,通过引入注意力机制,能够在不丢失重要特征信息的前提下,将分类任务所需要的关注重点放在目标区域中;随后通过不同的全连接层来实现多任务学习,从而提高了网络的泛化能力。实验结果表明,该方法能够对阿尔兹海默症进行准确分类。在遥感图像分类领域中,王宁等[17]在随机森林、支持向量机和BP神经网络的基础上设计出一种集成分类模型,并将其应用于水产养殖的水体资源遥感动态监测任务中。该模型很好地弥补了单个分类模型分类精度较低以及鲁棒性较差等缺点,在很大程度上避免了山体与建筑阴影等外在因素对水体特征提取的干扰。最终实验结果显示该分类模型拥有较高的分类精度,表明该模型能够成为水体资源遥感动态监测的分析工具之一。此外,朱明等[18]基于轻量级神经网络构建出一种鲈鱼捕食状态分类网络,满足了现代智能水产养殖的自动投喂需求。
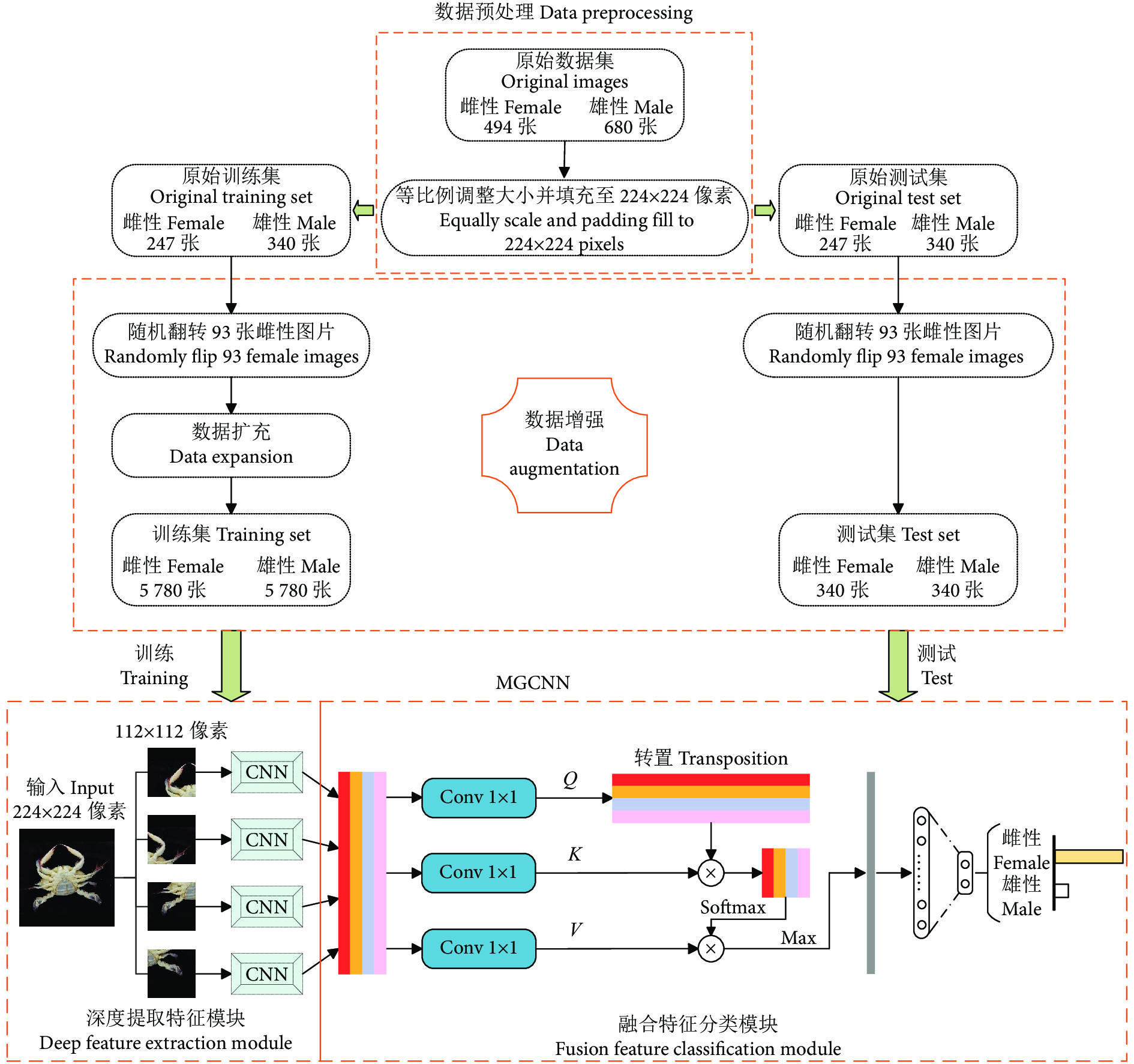
然而,尚未见图像分类的相关技术应用于梭子蟹性别识别的研究报道。为此,本文提出一种应用于梭子蟹性别识别任务的分类方法:首先构建一个用于梭子蟹性别分类的数据集PGCD;然后搭建一种多组卷积神经网络 (Multi-group convolutional neural network, MGCNN),并使用注意力机制更专注地找出输入数据的有用信息;最后通过一系列的调整参数工作提高MGCNN的分类精度。
1. 材料与方法
图1展示了本文所提方法的整体框架,包括数据预处理、数据增强以及MGCNN。 其中,MGCNN包含了深度提取特征模块和融合特征分类模块。数据预处理主要是将采集到的梭子蟹图像进行降低像素处理,从而解决后续处理速度过慢的问题;数据增强主要是对经过预处理后的梭子蟹图像进行数据扩充,解决了少量样本可能给网络带来的过度拟合问题,或者样本不均衡导致模型预测时对多数类的侧重问题;深度提取特征模块使用几组成熟的CNN 来提取图像的视觉特征,通过实验对比,最终的CNN本文选取ResNet50;融合特征分类模块主要是先将CNN提取到的特征进行信息融合,然后利用注意力机制代替传统池化层,在降低融合特征图维度的同时,去除了特征图的冗余信息,保留特征图的重要信息,最后经过输出分类层获取分类结果。
1.1 数据预处理
在公开数据库中,并没有梭子蟹性别分类的数据集,因此首先需要构建相关的数据集。本文的原始梭子蟹数据集主要由课题组在舟山水产品加工公司采集,采集所用设备的配置为10 800万像素的主摄+1 300万像素的超广角镜头+500万像素的长焦微距镜头。采集到的原始数据集共有1 174 张图像,其中雌、雄性图像各494和680 张,图像像素约为3 024×4 032。图2为部分采集的梭子蟹样品。
由于所采集的图像像素较大,导致后续操作中处理速度较慢,因此需要对图像进行像素降低处理,调整后的图像大小统一为224×224像素。传统的降低像素操作通常是利用opencv库的resize( ), 然而这种操作对于尺寸非N×N大小转化为N×N大小的图像来说,会改变图像原有的特征,影响网络的最终分类精度[19-21]。因此,本文采用一种等比例调整图像大小的方法,即在不改变宽高比的情况下进行图片调整,并填充至实验所需要的224×224像素的图像。resize ( ) 和等比例调整大小并填充的图像对比如图3所示。
1.2 数据增强
经过预处理后,将数据集随机分成原始训练集和原始测试集,均包含587 张梭子蟹图像 (雌性 247 张,雄性 340 张)。为了解决少量样本可能给网络带来的过度拟合问题,或者样本不均衡导致模型预测时对多数类的侧重问题,需要对原始数据集进行数据增强,以扩充出更多数据来提高数据集的数量和质量,提高网络模型的学习效果[22-24]。数据增强的过程如下所述:
(I) 随机翻转:为了解决样本不均衡问题,本文随机从原始训练集和原始测试集中各选取93 张雌性图像,进行随机翻转,从而使得原始训练集和原始测试集中雌性和雄性图像均有340 张。随机翻转是指从3种翻转方式中 (水平翻转、垂直翻转和水平-垂直翻转) 随机选取一种进行翻转。
(II) 随机旋转:对 (I) 中的原始训练集图像进行随机角度旋转,旋转角度在0°~90°、90°~180°、180°~270°和270°~360° 4个范围内进行随机选取。
(III) 随机平移:对 (I) 中的原始训练集图像进行随机像素平移,像素平移在−20至20之间进行随机选取,包括8个方向。
(IV) 随机明亮度:对 (I) 中的原始训练集图像进行随机明亮度调整,调整系数在0.8~1.0和1.0~1.2两个范围内随机选取。
(V) 随机噪声:对 (I) 中的原始训练集图像随机添加高斯噪声和椒盐噪声,添加噪声的系数均介于0.009~0.011随机选取。
值得注意的是,除了(I) 对训练集和测试集进行数据增强,(II)—(V) 均仅对训练集进行数据增强。其中,(IV) 和 (V) 主要是针对模型识别过程中图像出现的明亮度变化和噪声影响。通过 (IV) 和 (V) 两种数据增强技术可以让模型在现实环境中具有更好的应对能力。
5种不同类型的数据增强技术示例如图4所示。因此,经过数据增强后最终的数据集共有12 312 张梭子蟹图像,将其命名为PGCD。其中,训练集和测试集分别各有11 560 张 (雌性 5 780 张,雄性 5 780 张) 和680张 (雌性 340 张,雄性 340 张) 图像。
![]() 图 4 5 种不同类型的数据增强技术的示例Figure 4. Examples of five different types of data enhancement technologies
图 4 5 种不同类型的数据增强技术的示例Figure 4. Examples of five different types of data enhancement technologies1.3 深度提取特征模块
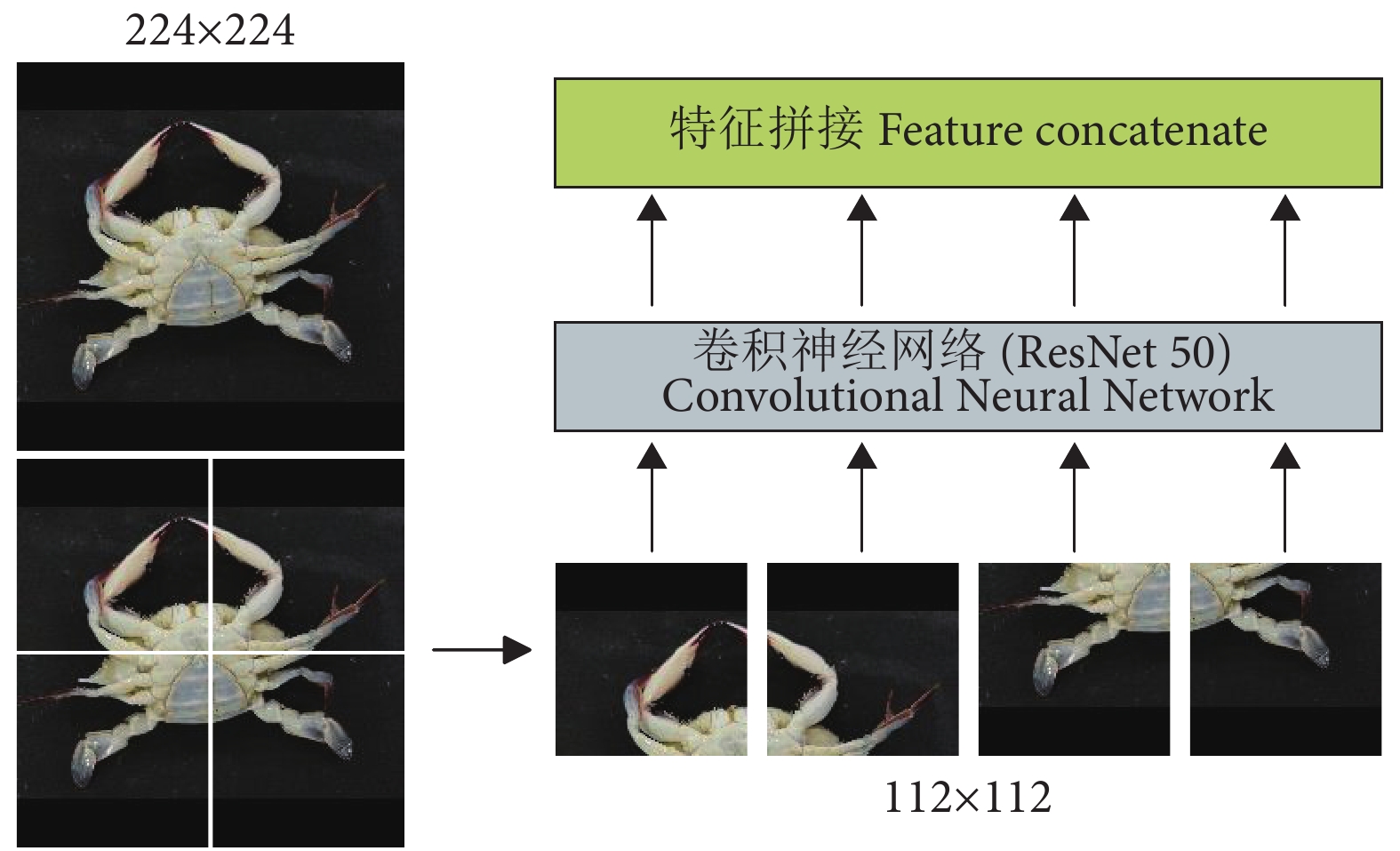
虽然Transformer[25]已经成为自然语言处理任务上的重要体系结构之一,但它在计算机视觉 (Computer Vision, CV) 领域的应用却非常有限。为了解决该问题,Dosovitskiy等[26]开发出一种Vision Transformer (ViT) 模型,可以直接应用于图像块序列 (Sequences of image patches),并且能够很好地执行图像分类任务。受这项工作的启发,本研究在提取特征之前设置了图像块来渲染每张图 (图5)。
具体操作是将输入图像切分成多个大小一致的图像块。在此假设输入图像的尺寸大小为N×N,需要切分的图像块数为n2,那么图像块的尺寸大小NP×NP用公式可表示为:
$$ {N_P} \times {N_P} = \frac{N}{n} \times \frac{N}{n} $$ (1) 式中:n为正整数。本文所提的MGCNN的输入图像尺寸为224×224 像素,图像块的个数n2=4,所以图像块的尺寸为112×112 像素。
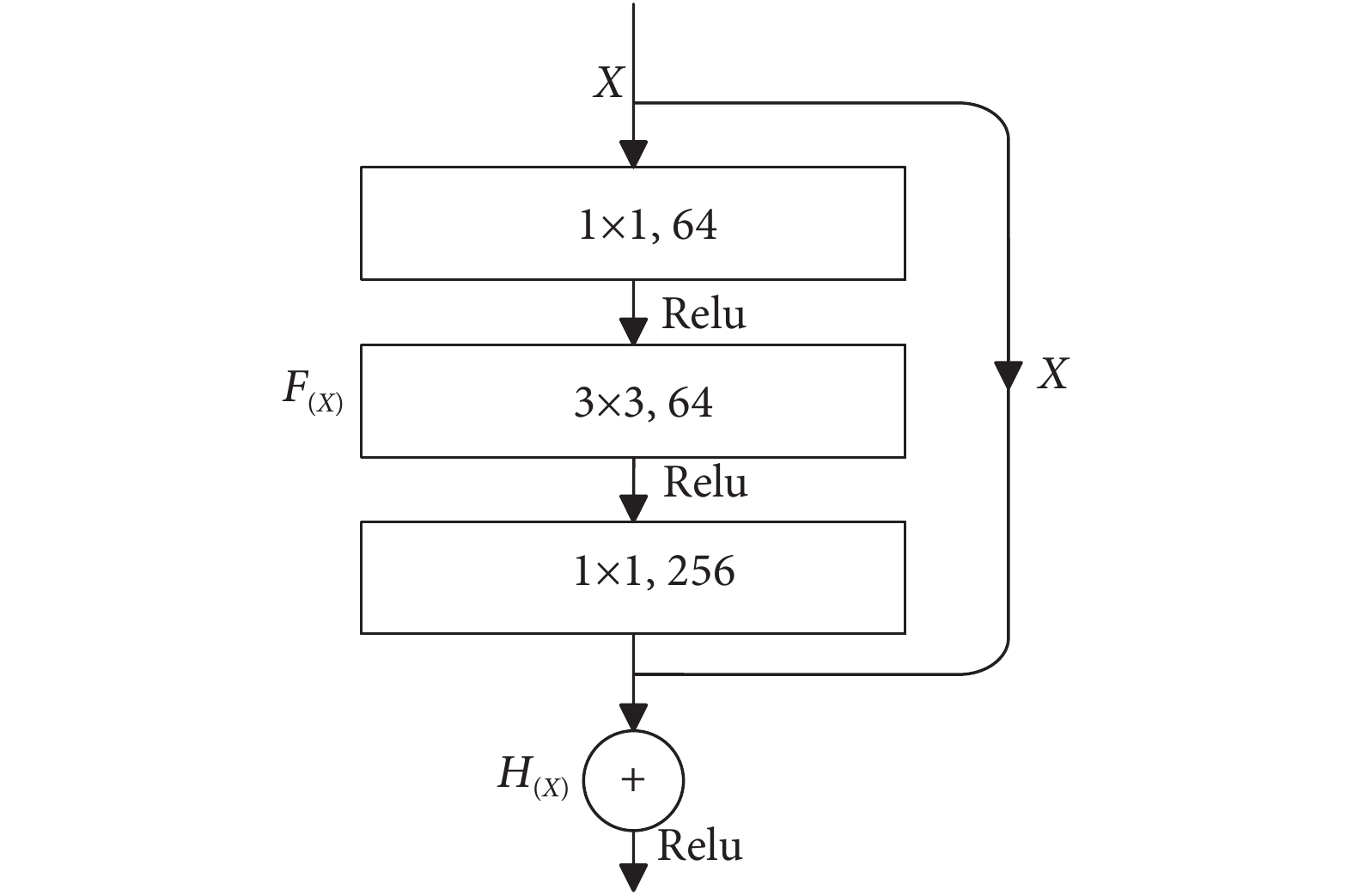
在4组图像块渲染图像之后,需要通过CNN来对每个图像块进行特征学习。为了减少特征提取过程的信息丢失,使得特征提取更有力,本文使用具有残差块的ResNet50作为深度提取特征模块中的主干CNN。ResNet50的残差块示意图见图6。
可以看出,残差块具有两条路径,一条是进行跳跃连接的输入特征X,另一条是经过3次卷积操作后得到的映射函数F(X),然后将这两条路径连接后就能得到残差模块的输出H(X),其过程可用公式 (2) 来表示:
$$ {H_{(X)}} = X + {F_{(X)}} $$ (2) 1.4 融合特征分类模块
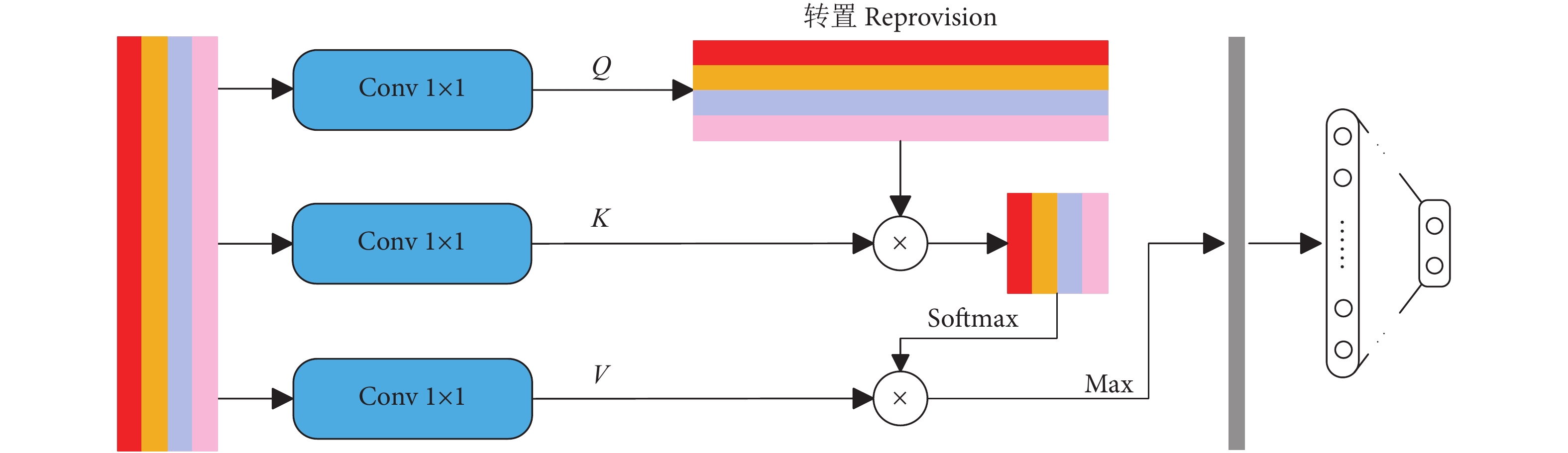
融合特征分类模块如图7所示。通过CNN学习不同图像块的视觉特征后,需要一个独特的全局描述符来表示图像。本文将不同图像块的视觉特征拼接 (Concatenate) 成一个全局特征图。最近一些研究表明,适当增加一些注意力机制可以更专注地找出输入数据的有用信息[27-29]。 为此,在分类之前本研究引入了一个注意力机制,来强调全局特征图中的细节重要性。首先,通过3个1×1的卷积层从全局特征图中获得Q、K、V 3个特征层;其次,将Q转置后与K做乘积运算形成一个新的特征图,再经过softmax函数后形成注意力权重;然后,将V与注意力权重做积运算,并使用最大池化对其结果进行降低维度处理;最后,通过全连接层获取待分类类别的概率分布。
2. 实验
2.1 实验设置
本实验均在同一环境配置的计算机上完成,训练集和测试集均来自于构建的PGCD数据集。用于实验的计算机配置为Windows10、NVidia GeForce GTX 1080 Ti GPU、16 GB 内存,实验基于Pytorch的方法[30]来实现,损失函数使用交叉熵损失 (Cross entropy loss) ,最大epoch值设置为100。通过实验对比,本研究提出方法的backbone model为ResNet50,学习率值设置为 0.000 1,batch size设置为32。此外,为了优化整体模型架构,本文使用 Adam[31]作为训练阶段优化器。
2.2 评估指标
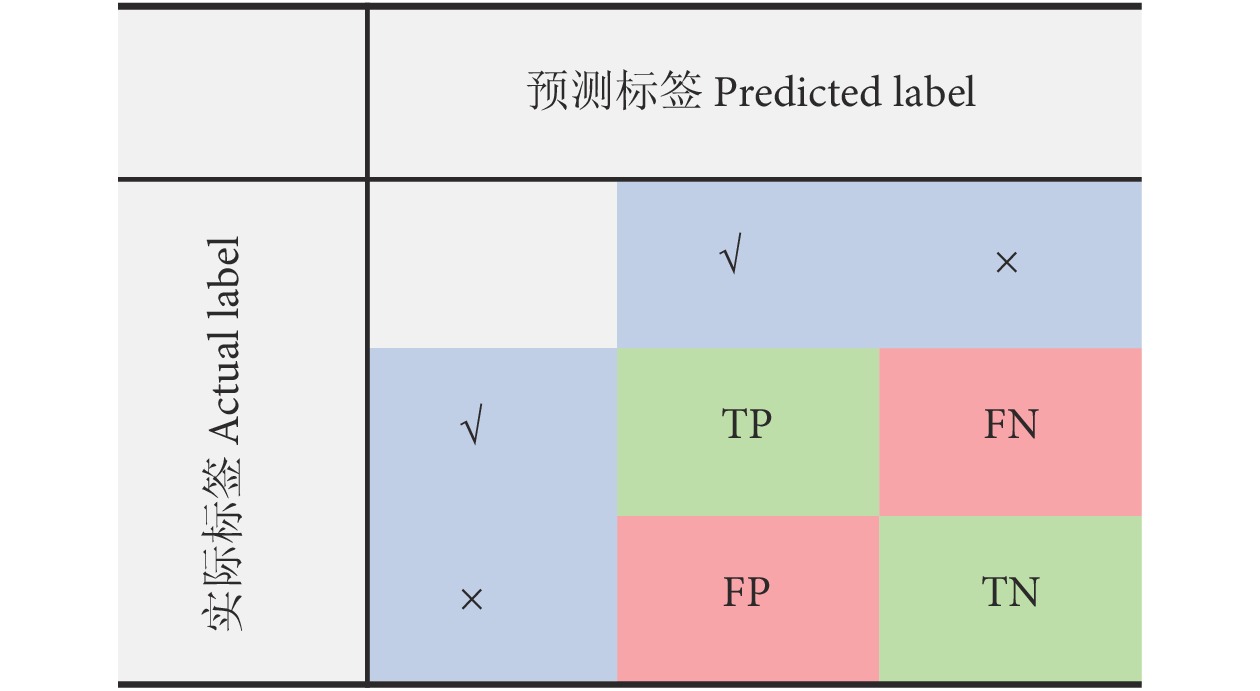
衡量模型分类性能的评估指标主要包括分类准确率 (又称分类精度)、召回率与查准率等[32-35]。本文主要采用这3个指标对所提方法进行评价。图8为混淆矩阵图。
![]() 图 8 混淆矩阵注:TP. 真阳性;FN. 假阴性;FP. 假阳性;TN. 真阴性。Figure 8. Confusion matrixNote: TP. True positive; FN. False negative; FP. False positive; TN. True negative.
图 8 混淆矩阵注:TP. 真阳性;FN. 假阴性;FP. 假阳性;TN. 真阴性。Figure 8. Confusion matrixNote: TP. True positive; FN. False negative; FP. False positive; TN. True negative.为此,分类准确率可用公式 (3) 进行表征:
$$ A_{\rm{cc}} = \frac{{{{\rm{TP}}} + {{\rm{TN}}}}}{{{{\rm{TP}}} + {{\rm{FP}}} + {{\rm{TN}}} + {{\rm{FN}}}}} $$ (3) 召回率可用公式 (4) 进行表征:
$$ R = \frac{{{{\rm{TP}}}}}{{{{\rm{TP}}} + {{\rm{FN}}}}} $$ (4) 查准率可用公式 (5) 进行表征:
$$ P = \frac{{{{\rm{TP}}}}}{{{{\rm{TP}}} + {{\rm{FP}}}}} $$ (5) 式中:Acc代表分类准确率 (Accuracy);R代表召回率 (Recall);P代表查准率 (Precision)。
2.3 实验分析
2.3.1 不同骨干模型对MGCNN性能的影响
成熟的卷积神经网络能够很好地进行特征学习和特征提取[36-37]。为此,本文主要选取VGG模型[10]和ResNet模型[12]作为MGCNN的深度提取特征模块的主干模型。不考虑DenseNet模型[23]最主要的原因是该模型属于较大的网络,需要训练更多的参数,并且可能由于过拟合导致分类精度提高不明显。而VGG和ResNet系列属于较小的网络,可以节省额外的计算开销以及训练时间。为了公平比较,在这里统一将学习率值设置为 0.001,批大小设置为32;此外,本文选取了 SGD[38]作为不同骨干模型的对比实验训练阶段的优化器。结果如表1所示,可以看出,在相同的学习率、批大小以及相同优化器的情况下,ResNet 系列普遍优于VGG系列。值得注意的是,ResNet152的精度未超过90%,再次验证了较大网络的过拟合现象会影响分类精度。由于ResNet50以92.79%的分类精度位居第一,因此本研究选取ResNet50作为MGCNN的骨干模型。
表 1 不同骨干模型对 MGCNN 性能的影响Table 1. Effects of different backbone models on MGCNN performance骨干模型
Backbone model准确率
Accuracy/%VGG VGG11 76.88 VGG13 82.59 VGG16 89.76 VGG19 86.09 ResNet ResNet18 90.44 ResNet34 91.15 ResNet50 92.79 ResNet101 92.21 ResNet152 88.24 2.3.2 不同优化器对MGCNN性能的影响
深度学习的过程中会产生一定的损失[39],所以应尽可能地减少损失来优化所提出的MGCNN模型,使其拥有更好的分类性能。事实上,深度神经网络的每一层都有各自的权重参数,这些权重参数决定着神经网络的输出。因此需要利用优化器 (Optimizer) 来降低损失,从而更新模型的可学习权重参数来优化网络模型。本研究以SGD[38]、AdaGrad[40]、RMSprop[41]、Adam[31]、Adamax[42]、ASGD[43] 6种优化器作为探讨优化器选取的对比实验。在这里学习率仍为 0.001,批大小为32,结果如表2所示。可以看出,在相同骨干模型的情况下,上述6种优化器中Adam最为突出,分类精度达到95.29%,均领先于其他优化器。因此,本文选取Adam作为MGCNN的优化器。
表 2 不同优化器对MGCNN性能的影响Table 2. Effects of different optimizers on MGCNN performance骨干模型
Backbone model优化器
Optimizer准确率
Accuracy/%ResNet50 SGD 92.79 AdaGrad 89.56 RMSprop 95.15 Adam 95.29 Adamax 93.82 ASGD 92.65 2.3.3 不同参数对MGCNN性能的影响
通过调整学习率以及批大小可以提高模型的分类性能[28,44-45],在MGCNN的骨干模型和优化器均确定的情况下 (ResNet50+Adam),本研究进一步探讨了不同参数对其分类性能的影响,以实现最佳精度。对于模型训练而言,学习率是控制权重更新的重要参数[46]。一方面,使用过大的学习率可能会忽略最优值的位置,导致模型不收敛;另一方面,使用过小的学习率容易出现过拟合,导致模型收敛缓慢。因此,首先应确定最优学习率。在本实验中,将最开始的学习率设置为0.000 1,批大小设置为32。本次实验遵循从较小的速率开始,并依次增加直到发现最佳学习率。当最佳学习率确定后,需要进行实验来确定最佳批大小,以优化网络训练的收敛速度和稳定性[45]。本研究依次将批大小设置为64、32和16来训练网络,最终的实验结果如表3所示。结果表明,当学习率和批大小分别为0.001 5和32时,MGCNN的分类效果最高,分类精度达到95.59%。因此,本文将最佳学习率设置为0.001 5,最佳批大小设置为32。
表 3 不同参数对 MGCNN 性能的影响Table 3. Effects of different parameters on MGCNN performance学习率
Learning rate批大小
Batch size准确率
Accuracy/%0.000 1 32 92.94 0.000 5 95.00 0.001 0 95.29 0.001 5 95.59 0.002 0 92.65 0.001 5 64 94.56 32 95.59 16 95.15 2.4 实验结果及可视化
为了评估所提方法MGCNN的分类性能,本文在已构建的PGCD数据集上进行了相关的分类任务实验,并将所提方法与其他先进的模型[9-10,12-13,46]进行比较,包括AlexNet、VGG16、ResNet152、InceptionV3和DenseNet121。这些模型之前均在用于图像分类的ImageNet数据集[9]上训练过,但并没有训练过PGCD。为了适应PGCD数据集,实验过程中将上述网络模型的输出分类层替换为具有两个类别 (雌性和雄性) 的输出分类层。
表4给出了MGCNN与一些先进方法的实验对比结果。可以看出,AlexNet显示了最差的性能;ResNet152和DenseNet121网络较深,召回率和查准率相对平衡,但分类精度未超过95%;InceptionV3分类精度达到95%,却因其查准率高导致召回率低;而本文所提的网络相比InceptionV3而言,分类精度有所提升,并且在查准率仅降低0.54%的情况下,将召回率提升1.76%。网络分类错误率最低,仅占4.41%,分类性能均领先于其他方法。
表 4 MGCNN 与先进方法的比较Table 4. Comparison between MGCNN and state-of-the-art methods方法
Method准确率
Accuracy/%召回率
Recall/%查准率
Precision/%错误率
Error/%AlexNet 54.71 59.36 67.93 45.29 VGG16 89.85 88.24 91.19 10.15 ResNet152 94.56 94.71 94.43 5.44 DenseNet121 94.41 94.41 94.41 5.59 InceptionV3 95.00 92.65 97.22 5.00 MGCNN
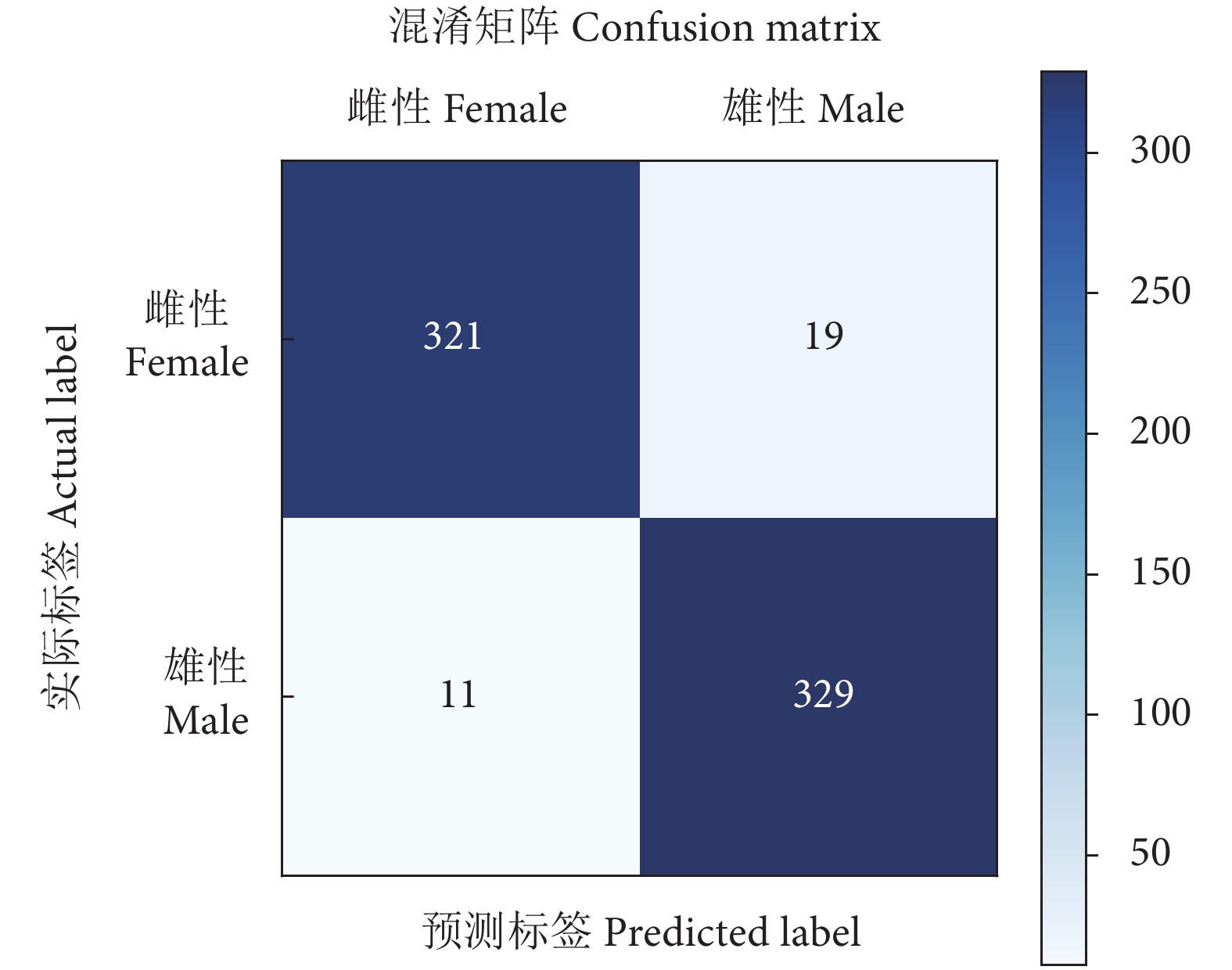
(本研究方法 Our method)95.59 94.41 96.68 4.41 为了更直观地显示所提方法的优越性,本文给出了PGCD测试集上梭子蟹性别分类的混淆矩阵 (Confusion matrix) ,如图9所示。图中主对角线显示的是识别正确的数目,副对角线显示的是识别错误的数目。可以看出,梭子蟹共有680 只,仅30 只被错误归类。其中,雌、雄蟹各有321、329 只被正确归类,仅11只雄蟹错归为雌性,19 只雌蟹错归为雄性。表明所提方法具有很好的分类效果。
![]() 图 9 梭子蟹性别分类的混淆矩阵Figure 9. Confusion matrix of gender classification of P. tritubereulatus
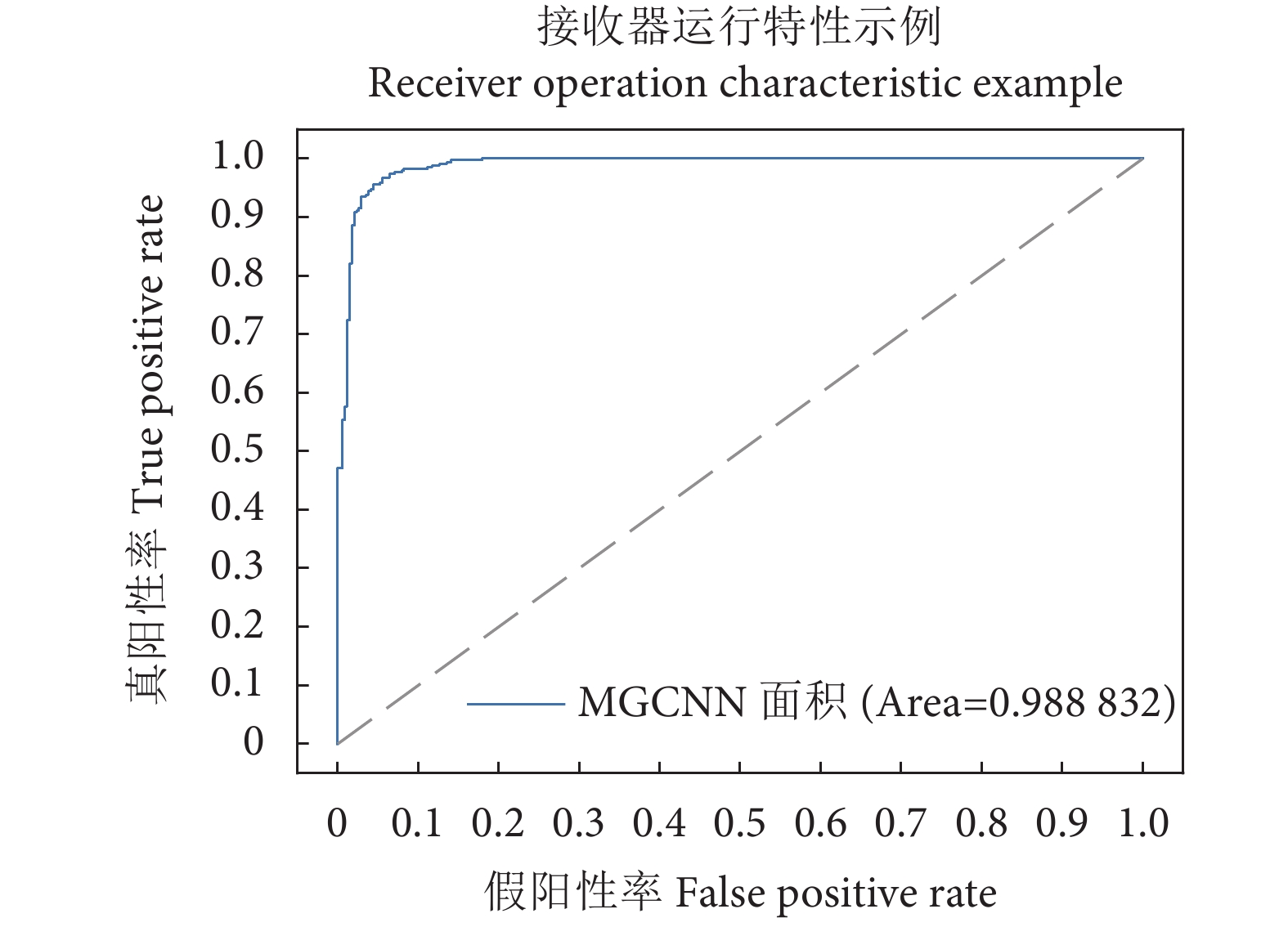
图 9 梭子蟹性别分类的混淆矩阵Figure 9. Confusion matrix of gender classification of P. tritubereulatus此外,受试者工作特征 (Receiver operating characteristic, ROC) 曲线和ROC曲线下的面积 (Area under the ROC curve, AUC) 也可以用于度量分类模型的好坏。MGCNN的ROC曲线和AUC如图10所示。可以看出,所提方法的AUC达到98.88%,在梭子蟹性别分类任务中取得优异的性能。
![]() 图 10 比较不同网络性能的受试者工作特征曲线 (ROC) 和 ROC曲线下的面积Figure 10. Comparison of Receiver Operating Characteristic (ROC) curve and area under ROC curve for subjects with different network performance
图 10 比较不同网络性能的受试者工作特征曲线 (ROC) 和 ROC曲线下的面积Figure 10. Comparison of Receiver Operating Characteristic (ROC) curve and area under ROC curve for subjects with different network performance2.5 单幅图像预测结果
为了验证模型的实际应用效果,采购雌、雄梭子蟹各20只,在实验室对其性别进行自动识别验证。验证图像采用等比例调整大小并填充的预处理方式。图11展示了单幅图像预测的示例,可以看出预测均正确,且预测概率均超过95%。此外,经统计分析,识别一张图像的时间不超过1 s。因此,本研究所提的算法可以很好地应用在梭子蟹性别自动分类及识别系统。
3. 小结
为了实现梭子蟹性别的智能化识别,促进现代渔业分拣装备由半机械化、机械化走向智能化,本文构建了梭子蟹性别分类数据集 (PGCD),提出了一种用于梭子蟹性别分类的多组卷积神经网络框架。该网络首先通过引入ResNet对图像块提取特征,减少特征提取过程的信息丢失,使得特征提取更有力;然后提出一种注意力机制代替传统的池化层,从而更专注地找出输入数据的有用信息;最后进行了一系列的参数调整,使得所提的MGCNN拥有最优分类性能。实验结果表明,所提方法在PGCD数据集上具有分类优越性,分类准确率高达95.59%。未来,将围绕梭子蟹的尺寸、肥瘦、蟹钳完整程度等特征进行分类,进一步完善梭子蟹智能识别系统。
-
![]()
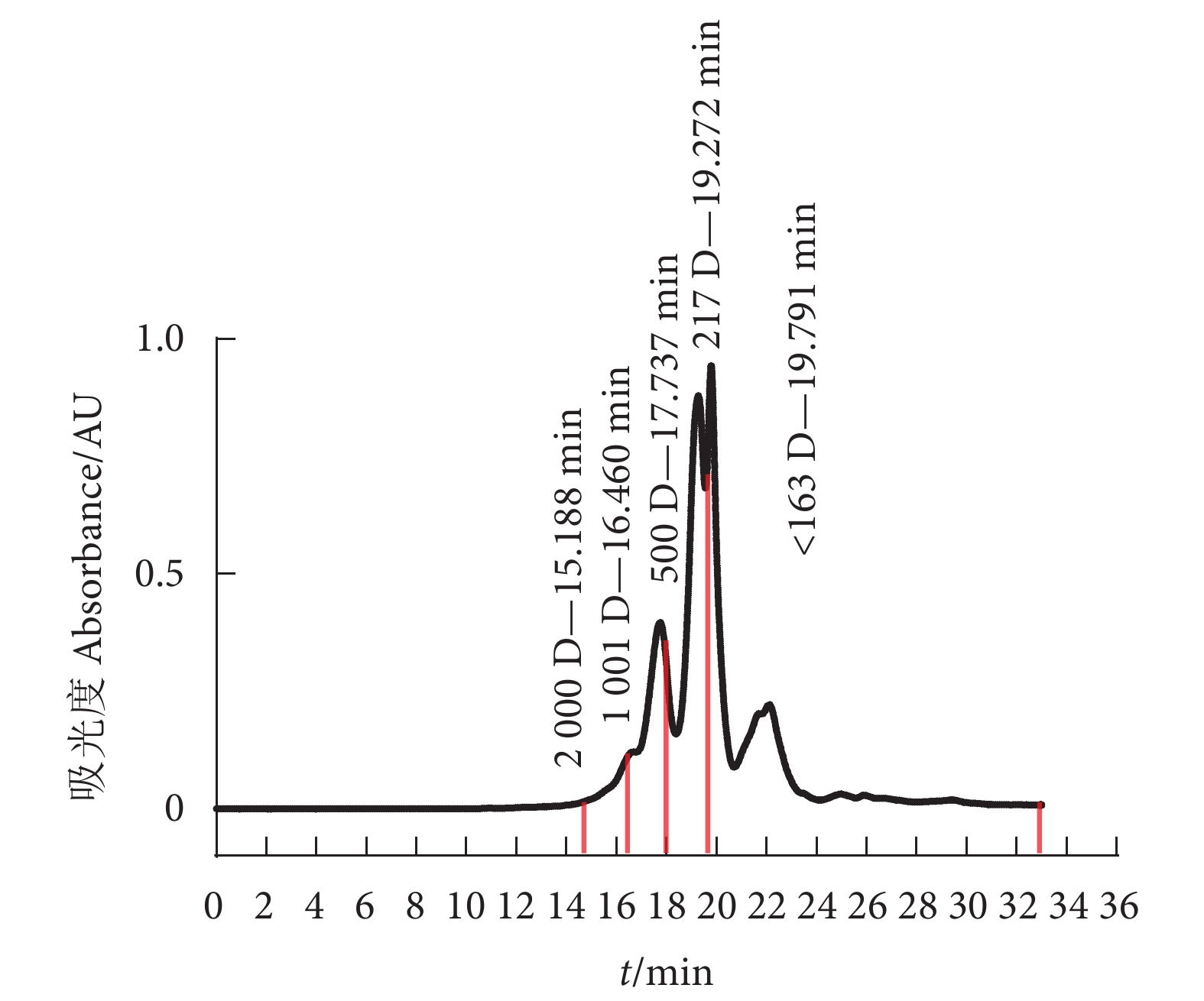
图 1 牡蛎肽相对分子质量分布色谱图
Figure 1. Chromatogram of relative molecular mass distribution of oyster peptides
![]()
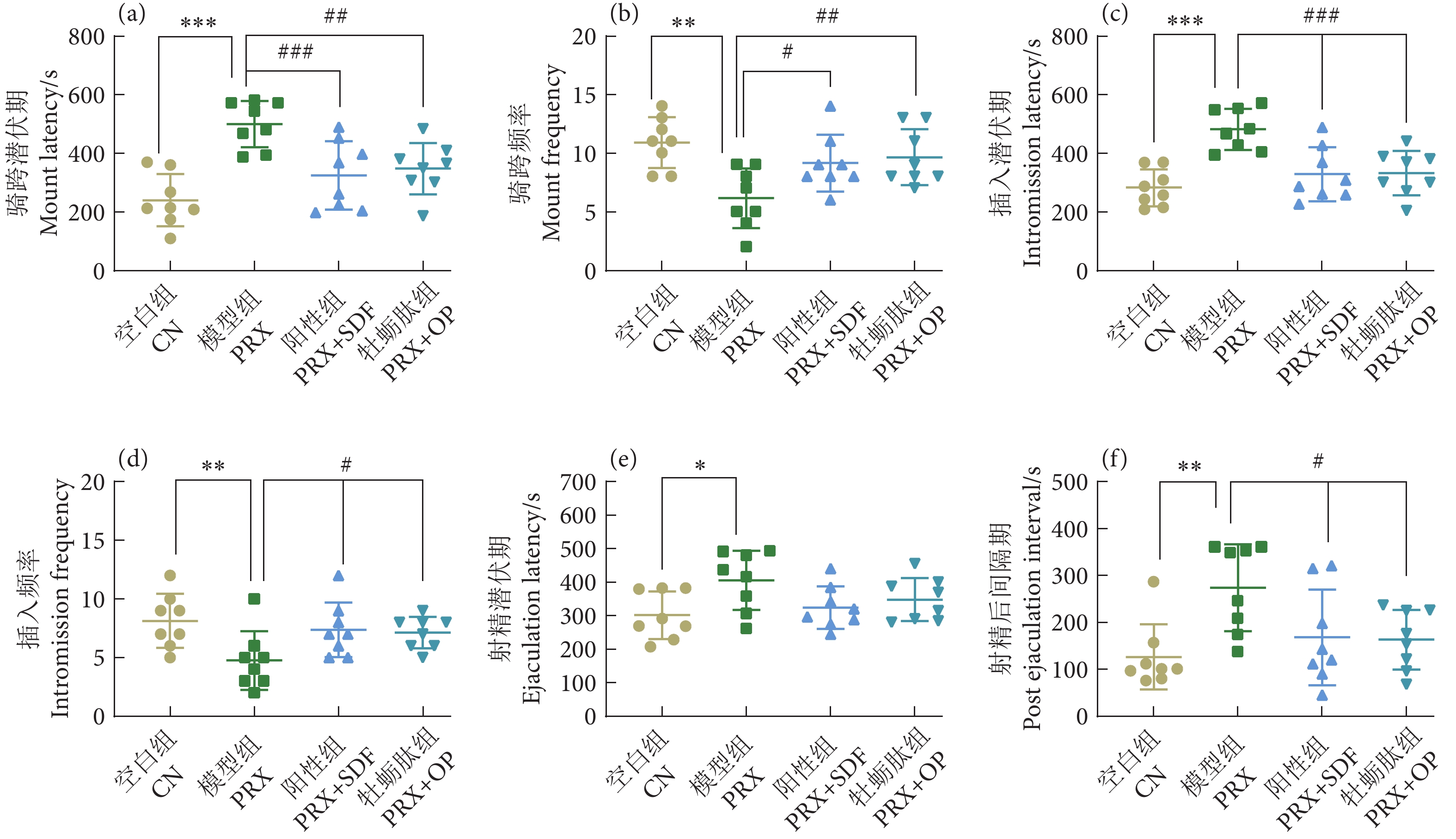
图 2 牡蛎肽对小鼠交配行为的影响
注:与空白组相比,*. 差异显著 (P<0.05),**. 差异非常显著 (P<0.01),***. 差异极显著 (P<0.001);与模型组相比,#. 差异显著 (P<0.05);##. 差异非常显著 (P<0.01),###. 差异极其显著 (P<0.001),下同。
Figure 2. Effect of oyster peptid on sexual behavior in mice
Note: Compared with the blank group, *. Significant difference (P<0.05); **. Very significant difference (P<0.01); ***. Extremely significant difference (P<0.001). Compared with the model group, #. Significant difference (P<0.05); ##. Very significant difference (P<0.01); ###. Extremely significant difference (P<0.001). The same case in the following figures.
![]()
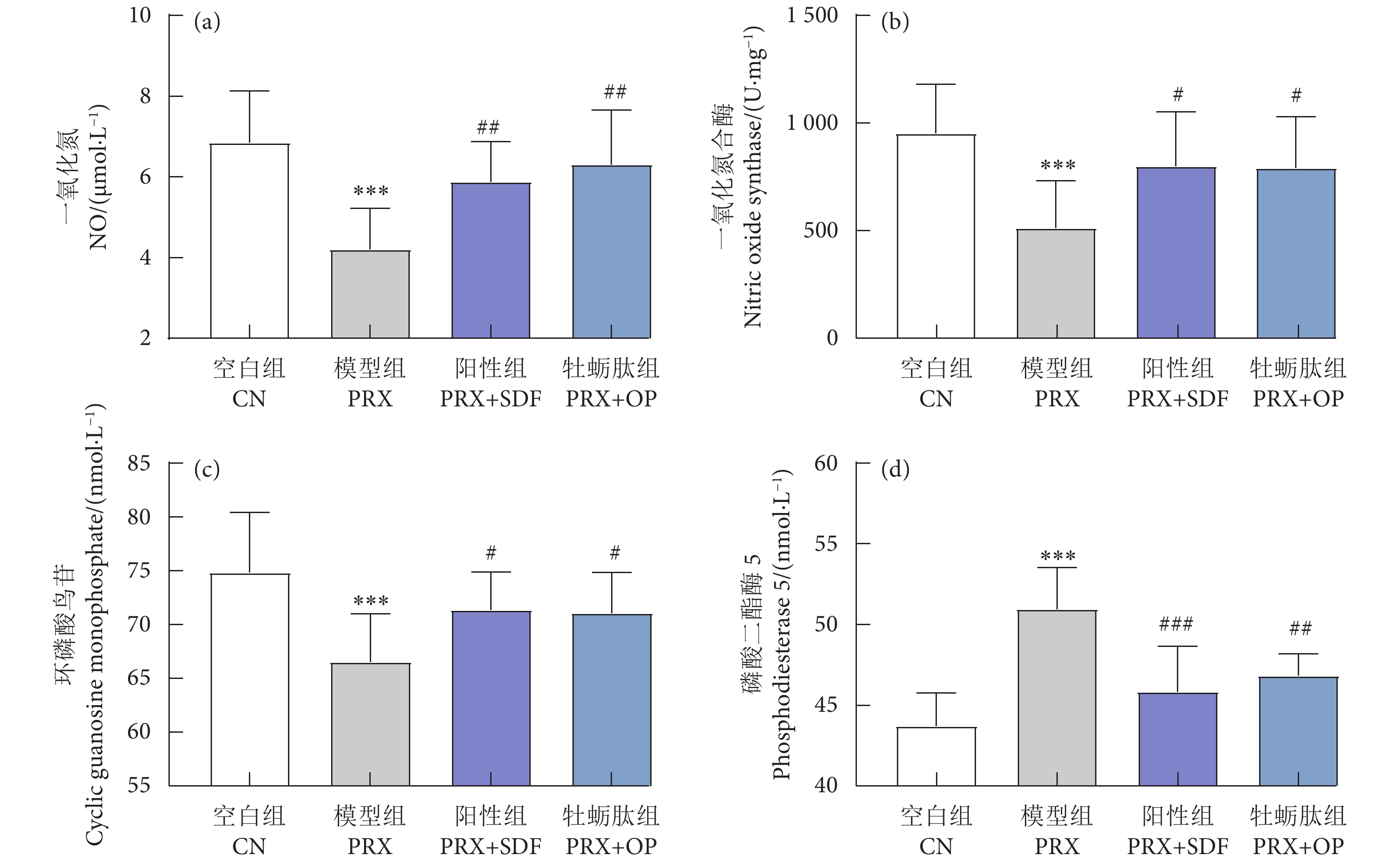
图 4 牡蛎肽对阴茎组织一氧化氮、一氧化氮合酶、环磷酸鸟苷、磷酸二酯酶-5含量或活性的影响
Figure 4. Effect of OP on content or activity of NO, NOS, cGMP and PDE5 in penile tissue
![]()
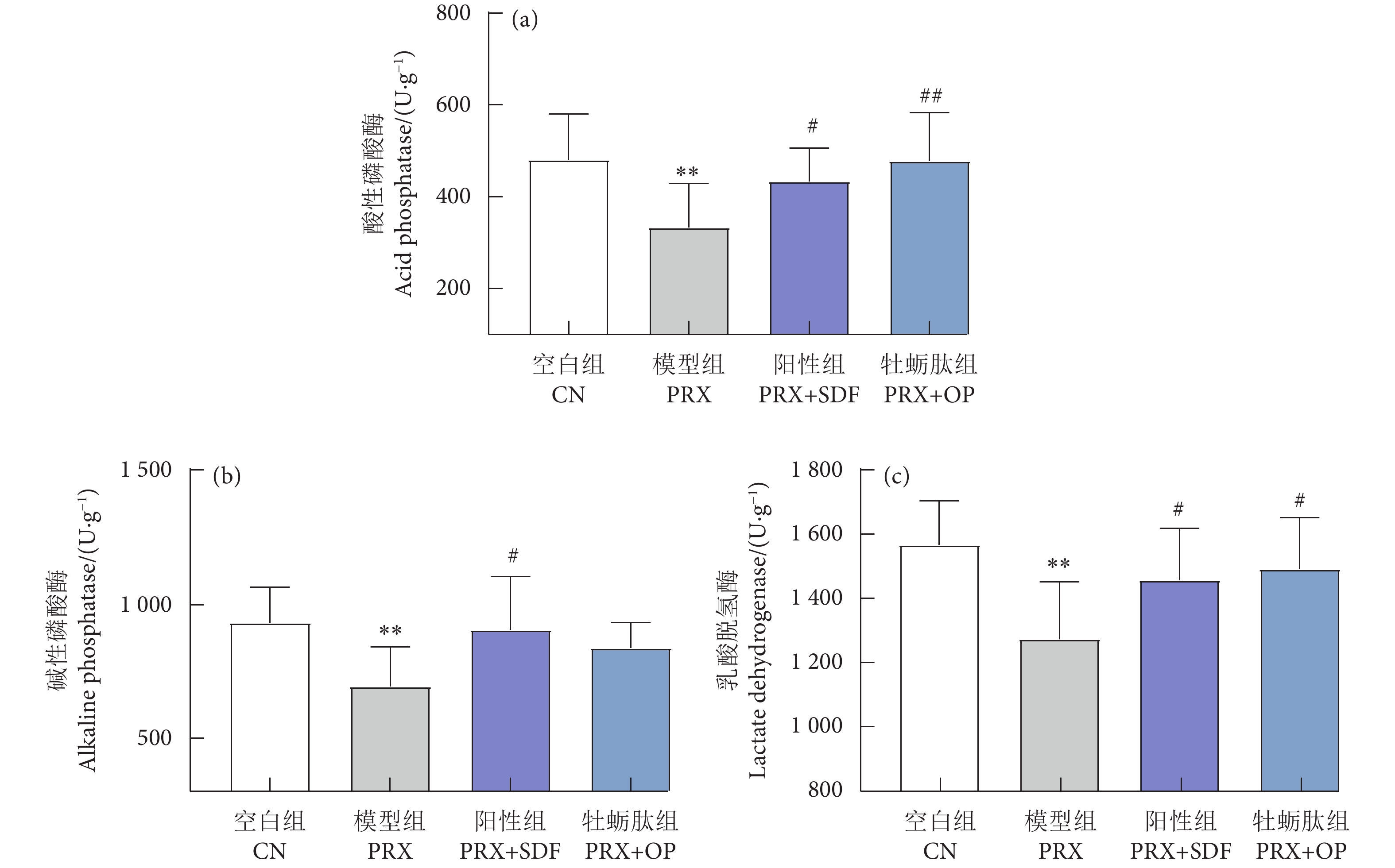
图 5 牡蛎肽对睾丸组织标志性酶的影响
Figure 5. Effect of oyster peptid on marker enzymes of testis tissue
![]()
图 6 牡蛎肽对睾丸组织抗氧化能力的影响
Figure 6. Effect of oyster peptid on antioxidant capacity of testis tissue
![]()
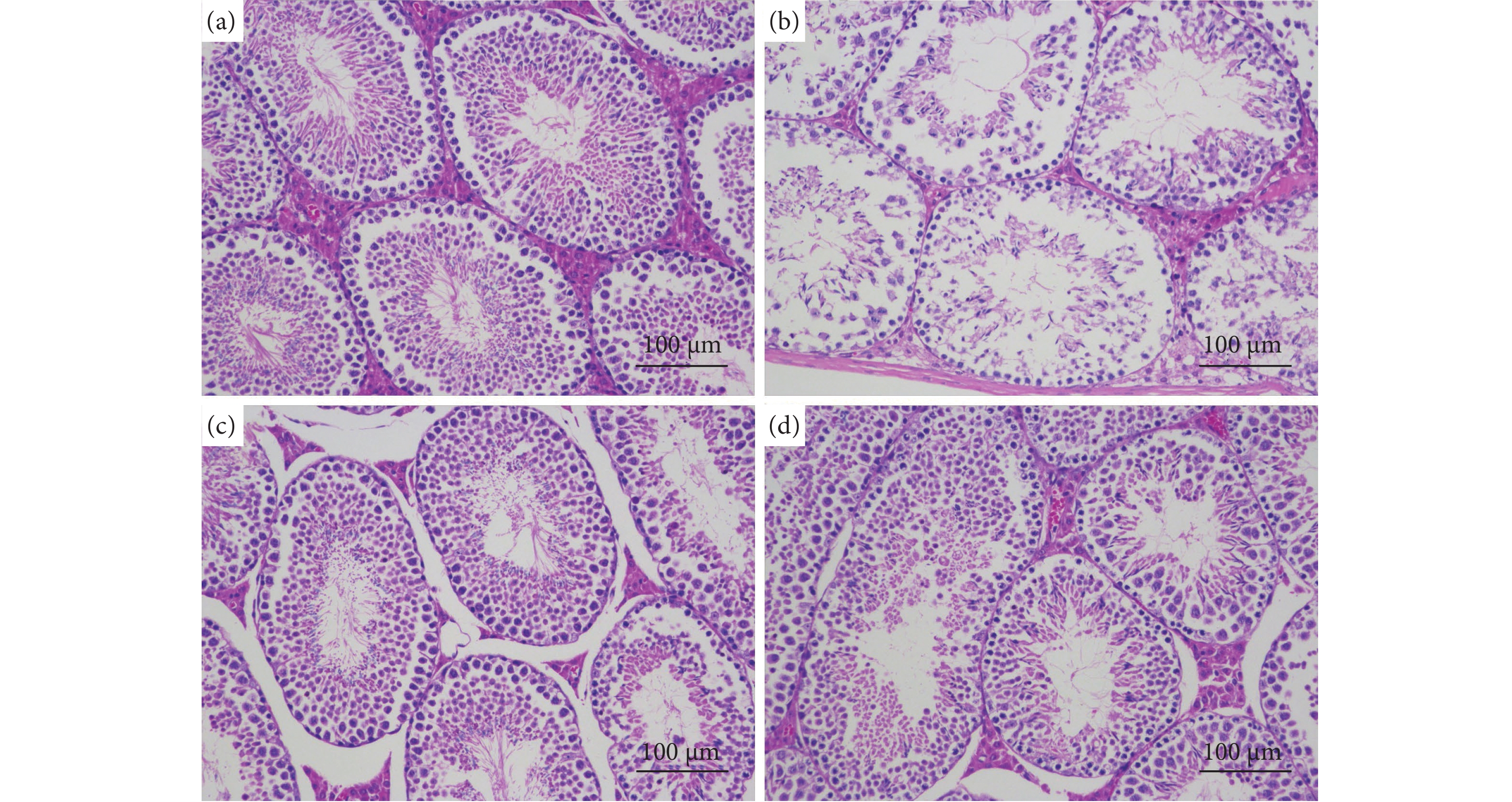
图 7 睾丸组织病理分析
注:a. 空白组;b. 模型组;c. 阳性组;d. 牡蛎肽组。
Figure 7. Pathological analysis of testicular tissue
Note: a. Control group; b. PRX group; c. PRX+SDF group; d. PRX+OP group.
![]()
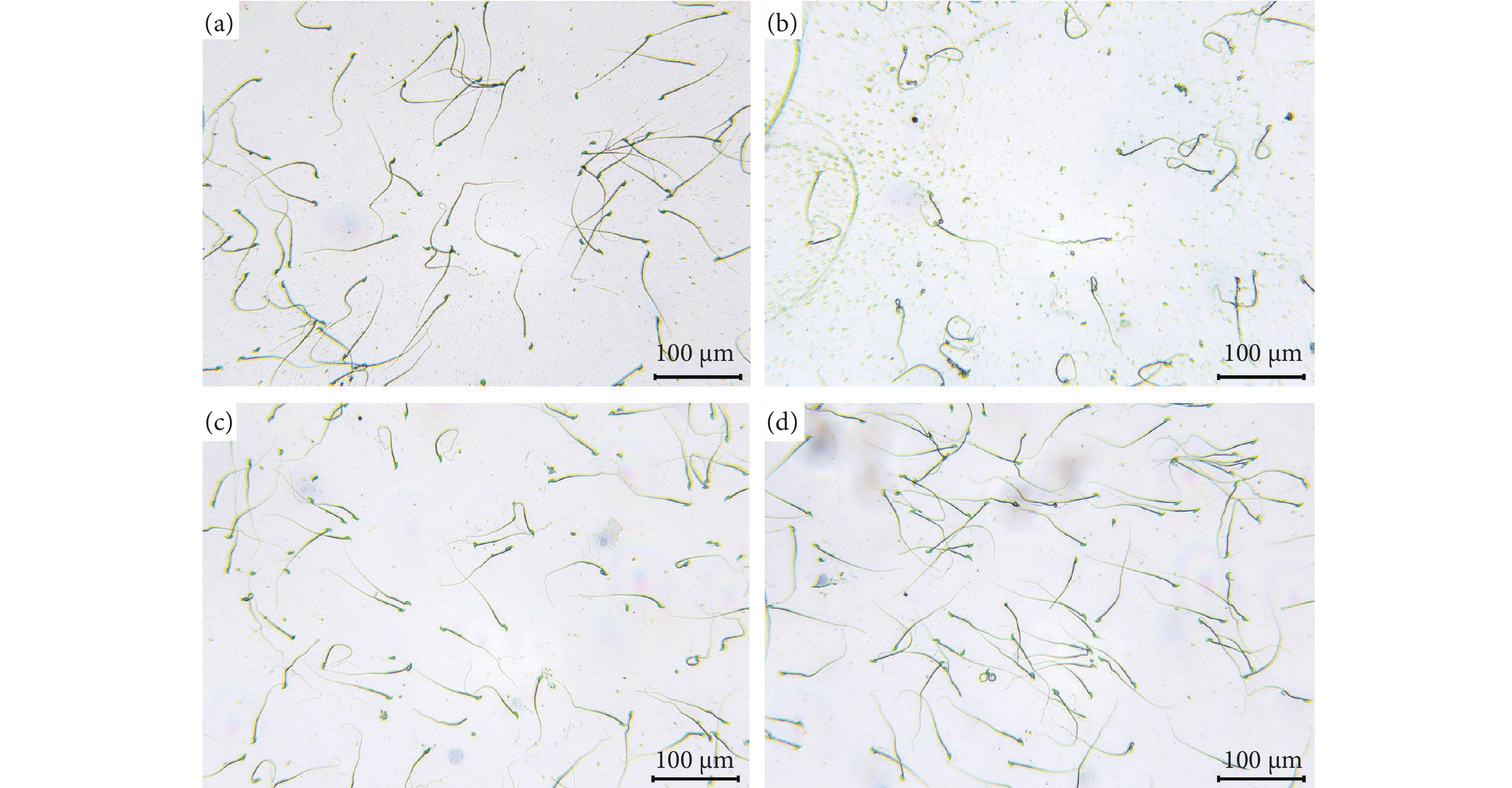
图 9 精子形态分析
注:a. 空白组;b. 模型组;c. 阳性组;d. 牡蛎肽组。
Figure 9. Sperm morphology analysis
Note: a. Control group; b. PRX group; c. PRX+SDF group; d. PRX+OP peptide group.
表 1 牡蛎肽游离氨基酸含量分析
Table 1 Analysis of free amino acid content of oyster peptid
序号
No.游离氨基酸
Free amino acid质量分数或含量
Mass fraction or content/%1 天门冬氨酸 Asp 0.39±0.032 2 苏氨酸 Thr① 0.41±0.031 3 丝氨酸 Ser 0.30±0.021 4 谷氨酸 Glu 0.86±0.052 5 脯氨酸 Pro② 0.07±0.015 6 甘氨酸 Pro 0.31±0.033 7 丙氨酸 Ala② 1.26±0.065 8 胱氨酸 Cys 0.04±0.000 9 缬氨酸 Val①②③ 0.57±0.145 10 蛋氨酸 Met②③ 0.26±0.036 11 异亮氨酸 Ile①②③ 0.76±0.111 12 亮氨酸 Leu①②③ 1.68±0.166 13 酪氨酸 Tyr 0.96±0.030 14 苯丙氨酸 Phe①② 1.13±0.267 15 赖氨酸 Lys① 2.14±0.248 16 组氨酸 His 0.29±0.019 17 精氨酸 Arg 2.68±0.142 氨基酸总量
Total amino acid, TAA14.10 疏水性氨基酸
Hydrophobic amino acid, HAA5.74 必需氨基酸
Essential amino acid, EAA6.95 支链氨基酸
Branched-chain amino acids, BCAA3.01 EAA/TAA (%) 49.29 BCAA/TAA (%) 21.35 HAA/TAA (%) 40.71 注:n=3;① 必需氨基酸;② 疏水性氨基酸;③ 支链氨基酸。 Note: n=3; ① Essential amino acid; ② Hydrophobic amino acid; ③ A branched-chain amino acid.  下载: 导出CSV
下载: 导出CSV
表 2 牡蛎肽对脏器系数的影响
Table 2 Effect of oyster peptid on organ coefficient
% 项目
Item空白组
CN模型组
PRX阳性组
PRX+SDF牡蛎肽组
PRX+OP心脏 Heart 0.599±0.067 0.581±0.072 0.597±0.082 0.622±0.079 胸腺 Thymus 0.11±0.029 0.104±0.021 0.116±0.46 0.113±0.022 脾 Lien 0.328±0.023 0.305±0.026 0.335±0.042 0.309±0.041 肝脏 Liver 5.106±0.586 4.798±0.351 5.004±0.581 5.018±0.306 肾脏 Ren 1.682±0.117 1.629±0.094 1.638±0.151 1.669±0.069 肺 Lung 0.657±0.061 0.637±0.048 0.641±0.034 0.652±0.039 阴茎 Penis 0.120±0.019 0.109±0.026 0.119±0.017 0.119±0.012 睾丸 Testis 0.759±0.083 0.629±0.061*** 0.699±0.069# 0.685±0.037# 精囊腺 Seminal vesicle 0.839±0.108 0.574±0.097*** 0.687±0.084# 0.731±0.061###
下载: 导出CSV
-
[1] CHEN L, SHI G R, HUANG D D, et al. Male sexual dysfunction: a review of literature on its pathological mechanisms, potential risk factors, and herbal drug intervention[J]. Biomed Pharmacother, 2019, 112: 108585. doi: 10.1016/j.biopha.2019.01.046
[2] RAHMAN A U, ALAM F, REHMAN Z U, et al. Effects of Mirabilis jalapa L. root extract and sildenafil on paroxetine-induced sexual dysfunction in male rats and characterization of its phytoconstituents by UPLC-MS[J]. S Afr J Bot, 2023, 152: 240-246. doi: 10.1016/j.sajb.2022.12.004
[3] ERDEMIR F, ATILGAN D, FIRAT F, et al. The effect of sertraline, paroxetine, fluoxetine and escitalopram on testicular tissue and oxidative stress parameters in rats[J]. Int Braz J Urol, 2014, 40: 100-108. doi: 10.1590/S1677-5538.IBJU.2014.01.15
[4] NAJAFABADI B T, FARSINEJAD M, SHOKRAEE K, et al. Possible effects of saffron (Crocus sativus) in the treatment of erectile dysfunction: a randomized, double-blind, placebo-controlled trial[J]. J Herb Med, 2022, 32: 100551. doi: 10.1016/j.hermed.2022.100551
[5] FARNIA V, ALIKHANI M, EBRAHIMI A, et al. Ginseng treatment improves the sexual side effects of methadone maintenance treatment[J]. Psychiat Res, 2019, 276: 142-150. doi: 10.1016/j.psychres.2019.05.004
[6] KOLOKO B L, BUSHRA I, WANKEU-NYA M, et al. In vivo effects of Rauvolfia vomitoria (Apocynaceae) ethanolic extract on sexual performance and reproductive activity in male rats[J]. Andrologia, 2020, 52(1): e13414.
[7] LI L, CHEN B B, AN T, et al. BaZiBuShen alleviates altered testicular morphology and spermatogenesis and modulates Sirt6/P53 and Sirt6/NF-κB pathways in aging mice induced by D-galactose and NaNO2[J]. J Ethnopharmacol, 2021, 271: 113810. doi: 10.1016/j.jep.2021.113810
[8] ALLOUH M Z, DARADKA H M, BARBARAWI M M A, et al. Fresh onion juice enhanced copulatory behavior in male rats with and without paroxetine-induced sexual dysfunction[J]. Exp Biol Med, 2014, 239(2): 177-182. doi: 10.1177/1535370213508360
[9] TEIXEIRA T M, BOEFF D D, de OLIVEIRA CARVALHO L, et al. The traditional use of native Brazilian plants for male sexual dysfunction: evidence from ethnomedicinal applications, animal models, and possible mechanisms of action[J]. Biomed Pharmacother, 2023: 116876.
[10] 郑环宇, 高加龙, 章超桦, 等. 华贵栉孔扇贝肉及其酶解产物对半去势雄性大鼠生殖能力的影响[J]. 南方水产科学, 2021, 17(3): 94-101. [11] QIONG L, JUN L, JUN Y, et al. The effect of Laminaria japonica polysaccharides on the recovery of the male rat reproductive system and mating function damaged by multiple mini-doses of ionizing radiations[J]. Environ Toxicol Phar, 2011, 31(2): 286-294. doi: 10.1016/j.etap.2010.11.006
[12] IBRAHIM N M, IBRAHIM S R, ASHOUR O H, et al. The effect of red seaweed (Chondrus crispus) on the fertility of male albino rats[J]. Saudi J Biol Sci, 2021, 28(7): 3864-3869. doi: 10.1016/j.sjbs.2021.03.059
[13] JE J G, KIM H S, LEE H G, et al. Low-molecular weight peptides isolated from seahorse (Hippocampus abdominalis) improve vasodilation via inhibition of angiotensin-converting enzyme in vivo and in vitro[J]. Process Biochem, 2020, 95: 30-35. doi: 10.1016/j.procbio.2020.04.016
[14] RYU B M, KIM M J, HIMAYA S W A, et al. Statistical optimization of high temperature/pressure and ultra-wave assisted lysis of Urechis unicinctus for the isolation of active peptide which enhance the erectile function in vitro[J]. Process Biochem, 2014, 49(1): 148-153. doi: 10.1016/j.procbio.2013.09.019
[15] 张雪妍, 秦小明, 高加龙, 等. 牡蛎酶解工艺优化及其酶解产物对小鼠睾酮分泌的影响[J]. 广东海洋大学学报, 2019, 39(3): 96-102. [16] 章超桦. 牡蛎营养特性及功能活性研究进展[J]. 大连海洋大学学报, 2022, 37(5): 719-731. [17] 朱国萍, 章超桦, 曹文红, 等. 牡蛎酶解产物对小鼠学习记忆的影响[J]. 广东海洋大学学报, 2021, 41(4): 84-92. [18] ZHANG C, LV J T, QIN X M, et al. Novel antioxidant peptides from crassostrea hongkongensis improve photo-oxidation in UV-induced HaCaT Cells[J]. Mar Drugs, 2022, 20(2): 100. doi: 10.3390/md20020100
[19] ZHANG Z R, SU G W, ZHOU F B, et al. Alcalase-hydrolyzed oyster (Crassostrea rivularis) meat enhances antioxidant and aphrodisiac activities in normal male mice[J]. Food Res Int, 2019, 120: 178-187. doi: 10.1016/j.foodres.2019.02.033
[20] 张婷, 秦小明, 章超桦, 等. 牡蛎酶解产物改善睡眠作用效果研究[J]. 大连海洋大学学报, 2021, 36(3): 430-436. [21] ZHANG W W, WEI Y F, CAO X X, et al. Enzymatic preparation of Crassostrea oyster peptides and their promoting effect on male hormone production[J]. J Ethnopharmacol, 2021, 264: 113382. doi: 10.1016/j.jep.2020.113382
[22] LUO X L, LIU W X, ZHONG H, et al. Synergistic effect of combined oyster peptide and ginseng extracts on anti-exercise-fatigue and promotion of sexual interest activity in male ICR mice[J]. J Funct Foods, 2021, 86: 104700. doi: 10.1016/j.jff.2021.104700
[23] 黄艳球. 牡蛎肉及其酶解产物对半去势雄性大鼠性功能的影响[D]. 湛江: 广东海洋大学, 2019: 12-19. [24] ZHANG X Y, PENG Z L, ZHENG H N, et al. The potential protective effect and possible mechanism of peptides from oyster (Crassostrea hongkongensis) hydrolysate on triptolide-induced testis injury in male mice[J]. Mar Drugs, 2021, 19(10): 566. doi: 10.3390/md19100566
[25] ADEMOSUN A O, ADEBAYO A A, OBOH G. Anogeissus leiocarpus attenuates paroxetine-induced erectile dysfunction in male rats via enhanced sexual behavior, nitric oxide level and antioxidant status[J]. Biomed Pharmacother, 2019, 111: 1029-1035. doi: 10.1016/j.biopha.2019.01.022
[26] 张锴佳, 张雪妍, 秦小明, 等. 香港牡蛎酶解产物对雷公藤甲素诱导雄性小鼠生精障碍的影响[J]. 大连海洋大学学报, 2022, 37(6): 941-948. [27] KHALID M, ALQARNI M H, WAHAB S, et al. Ameliorative sexual behavior and phosphodiesterase-5 inhibitory effects of Spondias mangifera fruit extract in rodents: in silico, in vitro, and in vivo study[J]. J Clin Sleep Med, 2022, 11(13): 3732. doi: 10.3390/jcm11133732
[28] CANPOLAT S, ULKER N, YARDIMCI A, et al. Irisin ameliorates male sexual dysfunction in paroxetine-treated male rats[J]. Psychoneuroendocrino, 2022, 136: 105597. doi: 10.1016/j.psyneuen.2021.105597
[29] 方磊, 张瑞雪, 陈亮, 等. 牡蛎肽对TM3细胞性功能的影响[J]. 中国食品学报, 2021, 21(5): 140-147. [30] HOU Y Q, WU G Y. Nutritionally essential amino acids[J]. Adv Nutr, 2018, 9(6): 849-851. doi: 10.1093/advances/nmy054
[31] 陈义明, 孙瑞坤, 张帅, 等. 圆舵鲣暗色肉酶解物5 ku组分的抗氧化活性与抗疲劳作用[J]. 广东海洋大学学报, 2017, 37(4): 92-97. doi: 10.3969/j.issn.1673-9159.2017.04.014 [32] 葛晓鸣, 顾伟, 徐永健. 海马水解蛋白的氨基酸组成与抗氧化能力的关系[J]. 核农学报, 2019, 33(2): 322-329. doi: 10.11869/j.issn.100-8551.2019.02.0322 [33] 陈悦, 李路, 闫朝阳, 等. 小分子牡蛎多肽对雄性小鼠性功能的影响[J]. 基因组学与应用生物学, 2019, 38(1): 109-116. doi: 10.13417/j.gab.038.000109 [34] SCHULSTER M, BERNIE A M, RAMASAMY R. The role of estradiol in male reproductive function[J]. Asian J Androl, 2016, 18(3): 435. doi: 10.4103/1008-682X.173932
[35] SAIKIA Q, HAZARIKA A, KALITA J C. Isoliquiritigenin ameliorates paroxetine-induced sexual dysfunction in male albino mice[J]. Reprod Toxicol, 2023: 108341.
[36] EL-GAAFARAWI I, HASSAN M, FOUAD G, et al. Toxic effects of paroxetine on sexual and reproductive functions of rats[J]. Egypt J Hosp Med, 2005, 21(1): 16-32. doi: 10.21608/ejhm.2005.18045
[37] YAKUBU M T, JIMOH R O. Carpolobia lutea roots restore sexual arousal and performance in paroxetine-induced sexually impaired male rats[J]. Rev Int Androl, 2014, 12(3): 90-99.
[38] AJIBOYE T O, NURUDEEN Q O, YAKUBU M T. Aphrodisiac effect of aqueous root extract of Lecaniodiscus cupanioides in sexually impaired rats[J]. J Basic Clin Physiol Pharmacol, 2014, 25(2): 241-248.
[39] ZHANG Y F, YANG J Y, MENG X P, et al. L-arginine protects against T-2 toxin-induced male reproductive impairments in mice[J]. Theriogenology, 2019, 126: 249-253. doi: 10.1016/j.theriogenology.2018.12.024
[40] BOLNICK J M, KILBURN B A, BOLNICK A D, et al. Sildenafil stimulates human trophoblast invasion through nitric oxide and guanosine 3', 5'-cyclic monophosphate signaling[J]. Fertil Steril, 2015, 103(6): 1587-1595.e2. doi: 10.1016/j.fertnstert.2015.02.025
[41] DEGIONI A, CAMPOLO F, STEFANINI L, et al. The NO/cGMP/PKG pathway in platelets: the therapeutic potential of PDE5 inhibitors in platelet disorders[J]. J Thromb Haemost, 2022, 20(11): 2465-2474. doi: 10.1111/jth.15844
[42] PACHER P, BECKMAN J S, LIAUDET L. Nitric oxide and peroxynitrite in health and disease[J]. Physiol Rev, 2007, 87(1): 315-424. doi: 10.1152/physrev.00029.2006
[43] YAKUBU M T, JIMOH R O. Aqueous extract of Carpolobia lutea root ameliorates paroxetine-induced anti-androgenic activity in male rats[J]. Middle East Fertil S, 2015, 20(3): 192-197. doi: 10.1016/j.mefs.2014.10.001
[44] OGUNRO O B, YAKUBU M T. Fadogia agrestis (Schweinf. Ex hiern) stem extract restores selected biomolecules of erectile dysfunction in the testicular and penile tissues of paroxetine-treated wistar rats[J]. Reprod Sci, 2023, 30(2): 690-700. doi: 10.1007/s43032-022-01050-6
[45] ADEFEGHA S A, OYELEYE S I, DADA F A, et al. Modulatory effect of quercetin and its glycosylated form on key enzymes and antioxidant status in rats penile tissue of paroxetine-induced erectile dysfunction[J]. Biomed Pharmacother, 2018, 107: 1473-1479. doi: 10.1016/j.biopha.2018.08.128



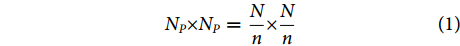
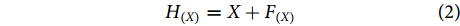
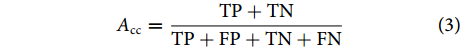
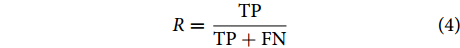
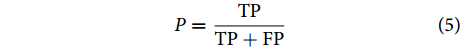

计量
- 文章访问数: 710
- HTML全文浏览量: 72
- PDF下载量: 28
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
