
Analysis of phytoplankton community in aquaculture crab pond using morphological and metagenomics methods
-
摘要:
为全面准确评估中华绒螯蟹 (Eriocheir sinensis) 养殖池塘中后期水体浮游植物群落结构特征及其与环境因子的关系,于2022年6—10月逐月监测水质指标,同时采用基于形态学和宏基因组学的人工镜检和高通量测序两种方法分析浮游植物的群落结构特征,探讨浮游植物优势种与环境因子的关系。结果显示,人工镜检鉴定的浮游植物共4门65种,而高通量测序数据分析所得浮游植物共4门1463种,其中人工镜检鉴定的优势种为蓝藻门的水华微囊藻 (Microcystis flos-aquae)、细小隐球藻 (Aphanocapsa elachista) 以及硅藻门的中肋骨条藻 (Skeletonema costatum),而高通量测序数据分析所得优势种为蓝藻门的水华束丝藻 (Aphanizomenon flos-aquae)、铜绿微囊藻 (M. aeruginosa)。人工镜检方法计算所得多样性指数普遍低于高通量测序结果。两种方法共同分析得出溶解氧(DO)、叶绿素 a (Chl-a)、总氮 (TN) 和氨氮 (NH3-N) 是影响浮游植物群落结构的关键环境因子。建议采用人工镜检与高通量测序法相结合的方式分析养殖期间的浮游植物群落结构,以便更全面、准确地评估养殖生态系统中的浮游植物群落结构特征及其与环境因子的关系。
Abstract:In order to fully and accurately evaluate the characteristics of phytoplankton community structure and its relationship with environmental factors in the middle and late periods of Chinese mitten crab (Eriocheir sinensis) pond culture, we had monitored the water quality indexes and phytoplankton community of aquaculture crab ponds from June to October 2022, and identified the phytoplankton community by artificial microscopy and high-throughput sequencing based on morphological and metagenomics methods. Then we discussed the relationship between dominant phytoplankton species and environmental factors. The results show that 65 species in four phyla were identified by artificial microscopy, and 1 463 species by metagenomic data analysis, in which the dominant species identified by artificial microscopy were Microcystis flos-aquae, Aphanocapsa elachista and Skeletonema costatum, and the dominant species identified by metagenomic data analysis were Aphanizomenon flos-aquae and M. aeruginosa. The diversity index calculated by manual microscopy was generally lower than that of metagenomic results. Dissolved oxygen, chlorophyll a, total nitrogen and ammonia nitrogen were the key environmental factors affecting phytoplankton community structure. It is suggested to simutaneously use artificial microscopy and high-throughput sequencing methods to analyze phytoplankton community structure in aquacultural water, so as to evaluate the characteristics of phytoplankton community structure and its relationship with environmental factors in aquaculture ecosystems more comprehensively and accurately.
-
为拓展海水养殖空间、应对近岸水质污染加剧问题及寻求更高的海域利用率,近年来我国大力支持依托桁架网箱发展深远海养殖并取得一定成效[1]。桁架网箱型式多样,有半潜式、全潜式、坐底式、自升式等。其中,自升式桁架网箱作为一种新型网箱,目前建成下水的仅有“湾区横洲号”和“联塑L001”两座,凭借养殖主箱体可沿桩腿下潜入水的结构特性,在极端海况条件下网箱可以有效减小波流载荷冲击,保护设施及养殖生物安全,为更好地适应深远海养殖极端环境、降低养殖风险、提升养殖收益提供了一种选择。
凭借着高性价比优势,高密度聚乙烯重力式 (HDPE) 深水网箱是目前发展深远海养殖的主要设施,相关技术较为成熟,关于其研究也开展得比较全面,涉及网箱及网衣水动力[2-5]、浮架结构强度[6-7]、锚泊技术[8-9]等。目前大型桁架网箱研究也主要集中在水动力分析、系泊分析、桁架主体结构安全等方面。水动力方面,不少学者采用物理模型实验的方式进行验证[10-11],此类验证方法虽有较好的模拟效果,但实验周期较长,模拟实验所需材料循环利用率较低。随着海工分析软件的不断发展,也有利用计算机技术开展数值模拟研究的,例如崔勇等[12]分析比较了双层网底网箱与单层网底网箱的受力运动模型,为多层网底鲆鲽网箱设计优化提供了理论依据;张婧等[13]、Lei等[14]利用海洋工程软件OrcaFlex对深海网箱模型进行水动力分析;陈炫光等[15]分析了串联浮筒的大小、位置对船型网箱水动力特性的影响。此类方法极大地促进了海工软件与深海网箱的结合,为深海网箱水动力评估提供了高效的参考方法。系泊方面,张新昊[16]、张松等[17]针对系泊方案进行了对比优选;黄小华等[18]比较分析了一种半潜式桁架结构渔场平台在3种系泊方式、3种压载状态和6种波况条件下的动力响应情况,该研究结果为今后我国半潜式渔场平台发展提供了一定的理论参考。此外,不少学者在新的领域不断尝试,开展网箱养殖与海上风电领域融合发展、平台系泊张力评估新方法等方面的探索研究[19-22]。而针对桁架主体结构安全方面,李贺[23]评估了某大型网架式深海网箱的结构安全性,对设计不合理之处提出了改进措施;孙树政等[24]在不考虑网衣的作用下,对单个网箱模块浮架结构屈服强度进行校核;此外,类似的研究还有柴佳瑜等[25]、李昊瑾[26]、汤建锋[27]对不同环境载荷作用下网箱的强度进行了校核分析。综上所述,大部分学者在对网箱整体结构强度进行探讨时针对网衣部分的处理均考虑未系缚或采用简化模拟的形式,极少学者将研究方向聚焦于桁架网箱结构响应评估时网衣及网衣不同参数对其影响的程度。
由于自升式桁架网箱是一种新型网箱,关于其研究还比较有限,庞国良等[28]针对一种8桩腿自升式桁架网箱,基于ANSYS/Mechanical通过求解不同工况下网箱各部件结构变形情况及应力分布评估网箱结构安全性,并验证了这种型式网箱对极端环境条件的适用性,同样的方法也被用于对一种单桩腿八边形可组合式单元升降网箱的安全性分析[29],但网箱模型也仅考虑了网箱主体结构,忽略了网衣系统对网箱结构响应评估结果的影响。因此,本研究以自升式桁架网箱为研究对象,通过构建包含网衣系统的自升式桁架网箱模型,旨在探明网衣系统对桁架网箱结构响应的影响。
本研究基于SESAM软件开展相关仿真计算。首先建立了有无网衣的自升式桁架网箱数值模型,开展了不同工况下网箱结构响应仿真计算,从桩腿支反力及网箱桁架结构强度校核结果方面,对比分析了有无网衣对网箱结构响应的影响;然后基于附带网衣的网箱数值模型,进一步探讨了网衣不同群化比及不同数量系缚纲绳对桁架网箱结构响应评估的影响,研究可为大型桁架网箱结构响应评估提供技术参考。
1. 网箱计算模型
1.1 网箱结构参数
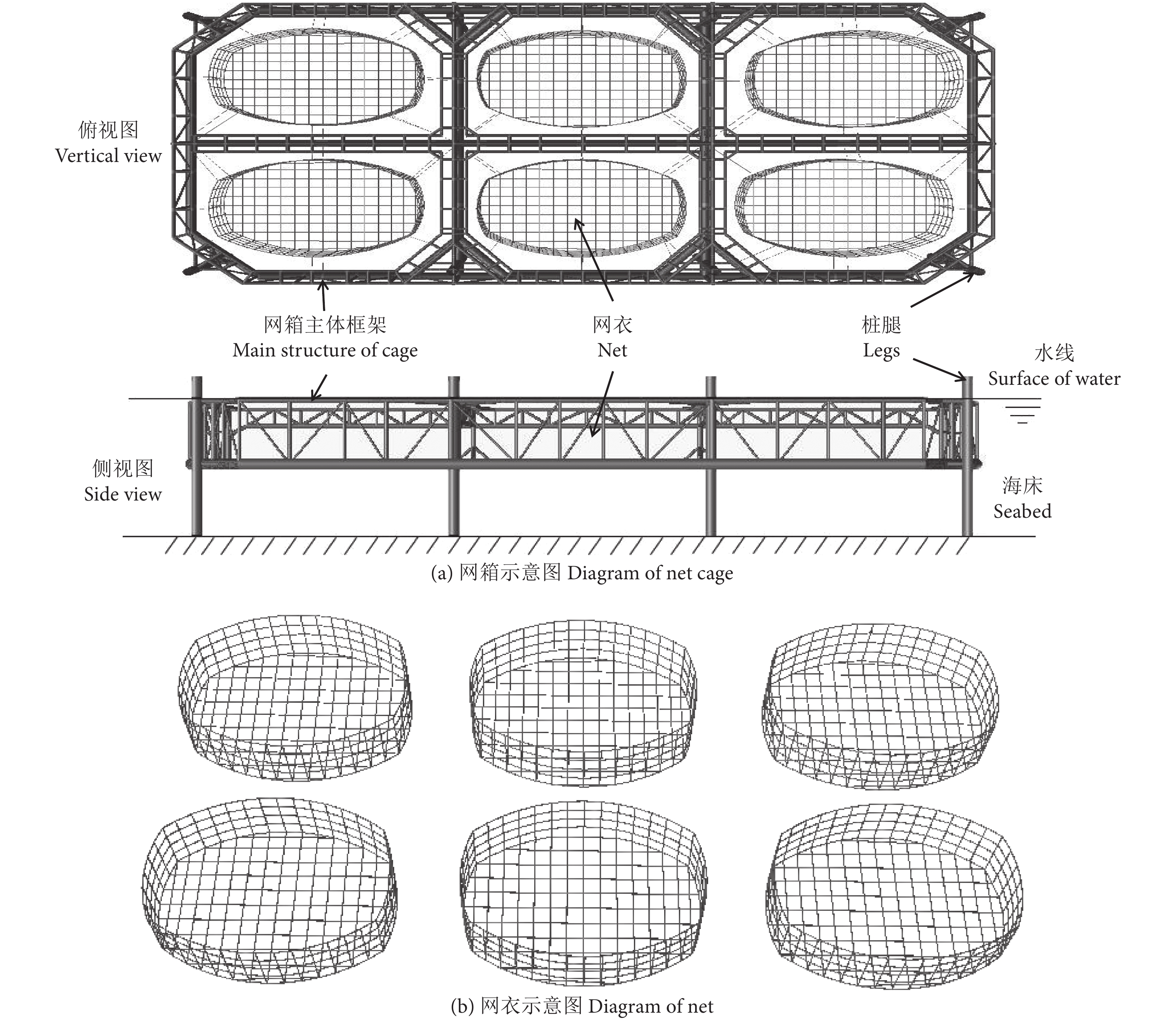
本研究对象为一种8桩腿自升式桁架网箱 (图1-a),网箱总长为133 m,总宽为70 m,主体高为10.3 m,分为6个养殖区,养殖水体总计约9×104 m3。网箱包括网箱主体框架、桩腿以及网衣系统3个部分,其中网衣通过纲绳系缚于网箱主体框架相邻桁架管上来构造养殖水体空间,网箱每个养殖区网衣形状类似于椭圆柱体 (图1-b),其中横轴长度为34 m,纵轴长度为24 m,高度为9.6 m,采用方形网目网衣作为网衣材料,网目目脚长度为60 mm,网线直径为3.5 mm,网箱主体可沿桩腿进行上下移动以满足不同工况需要,网箱桩腿及主体框架所使用的材料为Q345钢,网衣材料为尼龙,详细的网箱结构参数见表1。需要说明的是,本研究主要探讨网衣结构及部分网衣参数对网箱整体结构响应评估的影响,因此仅考虑了网箱主体框架上部处于水面以上的正常作业状态,吃水深度为9.3 m。
表 1 自升式桁架网箱尺寸及参数Table 1. Principal dimensions and material parameters of jack-up truss net cage参数 Parameter 数值 Value 网箱总长 Cage length/m 133 网箱总宽 Cage width/m 70 主体框架高度 Cage height/m 10.3 作业水深 Operation water depth/m 22 网箱吃水 Cage draught/m 9.3 桩腿高度 Leg height/m 26 网衣长度 Net length/m 34 网衣宽度 Net width/m 24 网衣高度 Net height/m 9.6 网衣网目尺寸 Mesh size of net/mm 60 1.2 网衣模拟方法
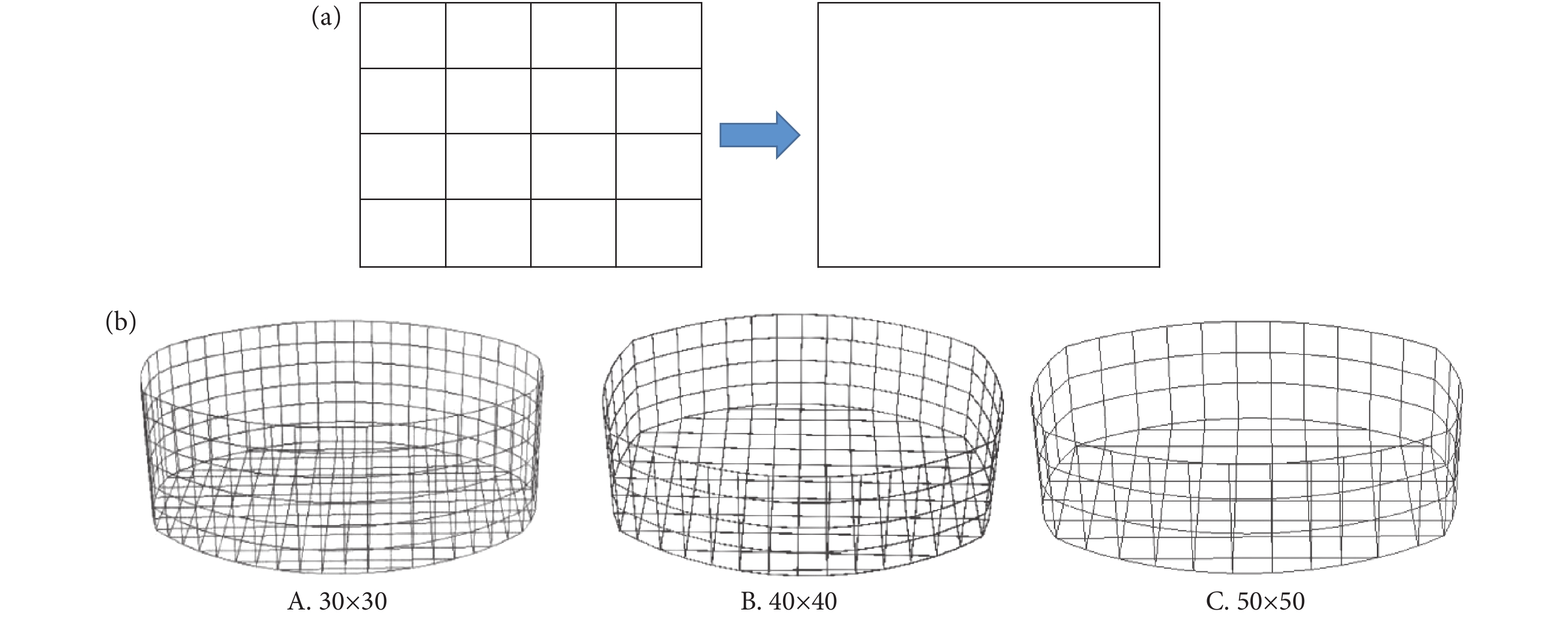
网衣在宏观尺度上是由数万计的细小网目组成,就本研究而言,网目目脚长度为60 mm,而网衣为长34 m×宽24 m×高9.6 m,如果对网衣采用考虑实际网目尺寸的精细建模方式,建模及计算成本将十分巨大。网目群化技术是将网衣中相邻多个实际网目合并成一个“大网目” (图2-a),进而在仿真中极大地减少了计算成本。本研究根据一种改进的网目群化方法对自升式网箱网衣进行等效简化[30],该方法重点考虑了网衣等效前后的水动力影响,根据Morison方程调整水动力系数,保持网衣等效前后所受水动力相同。为探讨网衣在不同群化比条件下对网箱结构响应的影响,本研究考虑了30×30 (即将30×30个网目等效为1个网目) 、40×40和50×50 3种网衣群化比。不同网衣群化比下的网衣群化后示意图见图2-b。
![]() 图 2 网目群化示意图 (a) 及不同群化比网衣群化后的网衣模型 (b)Figure 2. Diagram of mesh grouping (a) and net models with different mesh grouping ratios (b)
图 2 网目群化示意图 (a) 及不同群化比网衣群化后的网衣模型 (b)Figure 2. Diagram of mesh grouping (a) and net models with different mesh grouping ratios (b)1.3 网箱模型建立
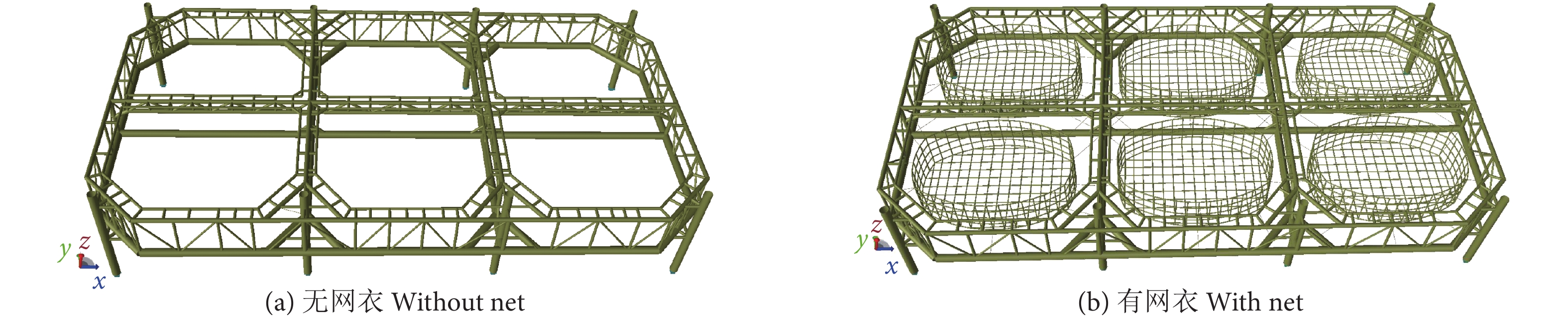
考虑到本研究中的自升式桁架网箱主体部分均为桁架管结构,桩腿和网衣均为细长杆件,因此在SESAM/GeniE建模中,将模型全部模拟成梁单元。此外,由于该网箱在长度和宽度方向均为轴对称结构,因此建模中仅需构建四分之一网箱模型,然后通过镜像操作便可实现网箱全部模型的建立。
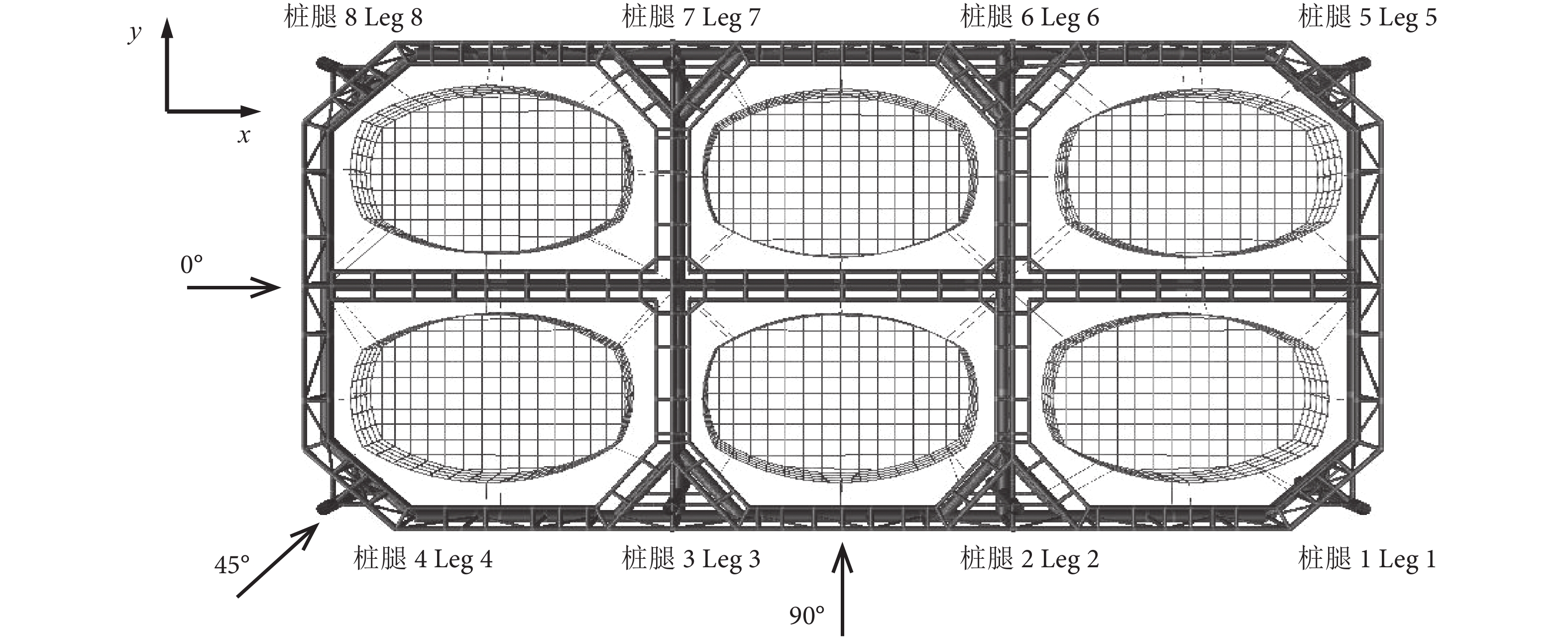
坐标系及网箱载荷方向见图3。其中,坐标原点位于网箱艉部中心处,x轴指向网箱艏部为正,y轴指向网箱左侧为正,z轴向上为正,将沿x轴正向定义为0° 环境载荷方向,沿y轴正向定义为90° 环境载荷方向。
由于本研究主要关注网箱主体结构响应,因此在建模中忽略了网箱桩靴和桩土的相互作用,在模型边界条件处理上,直接约束了8个桩腿底部节点所有方向自由度。网衣部分则是以网箱主体框架为基础,连接网衣至相邻桁架管上。最终建立的无网衣和有网衣的自升式桁架网箱数值模型见图4,随后通过上述模型研究网衣及不同网衣系缚方式对自升式桁架结构的影响,并以桩腿支反力和桁架利用系数 (Utilization factor, UF) 值的形式输出对比分析。
2. 计算理论与工况
2.1 环境载荷计算
网箱在实际作业中主要受到风、浪、流载荷作用。本研究的自升式网箱水线以上受风部件主要是8根桩腿,对网箱整体结构而言其受风载影响较小,因此分析中主要考虑波浪和海流两种载荷。另外,考虑到自升式网箱为轴对称结构 (图3),波流载荷方向上只考虑0°、45°、90° 3个方向。
自升式桁架网箱主要构件均为细长桁架管,相对于波浪波长而言为典型的小尺度细长构件,因此网箱结构所受波浪力可以利用Morison 公式进行计算,对于细长圆杆结构,所受到波浪力为:
$$ F_{ {{\rm{wave}} }}{\text{=}}\frac{1}{2} C_{\mathrm{d}} \rho D_{\rm{u}}|U|{\text{+}}C_{\rm{m}} \rho \frac{\pi D^2}{4} \frac{\partial u}{\partial {t}} $$ (1) 式中:
$ {C}_{\mathrm{d}} $ 为拖曳力系数;$ {C}_{\mathrm{m}} $ 为惯性力系数;$ \rho $ 为海水密度 (kg·m−3);D为桩柱直径 (m);$ {U} $ 为桩柱轴线处水质点的水平方向速度 (m·s−1);本研究取拖曳力系数$ {C}_{\mathrm{d}}{\text{=}}0.7 $ 和惯性力系数$ {C}_{\mathrm{m}}{\text{=}}1.2 $ 。对于海流载荷,根据稳定流动条件下的杆件阻力数学表达,可以得出桁架杆结构所受到海流力为:
$$ F_{\mathrm{c}}{\text{=}}\frac{1}{2} \rho C_{\rm{d}} A V_{\rm{c}}^2 $$ (2) 式中:
$ \rho $ 为海水密度 (kg·m−3);$ {C}_{\mathrm{d}} $ 为阻力系数,$ A $ 为迎流面积 (m2);$ {V}_{\mathrm{c}} $ 为海流流速 (m·s−1)。2.2 计算工况确定
波浪载荷方面,本研究考虑使用自升式网箱正常作业条件下的极端波浪载荷[21]来探讨网衣及不同网衣参数对自升式桁架网箱结构响应的影响,此时波高为6 m,波浪周期为10.8 s,波浪方向考虑0°、45°、90° 3个方向。海流载荷方面,以0.25 m·s−1为间隔,从0~1.5 m·s−1不等的流速条件,海流方向与波浪同向。波浪相位角选取对网箱结构安全校核十分重要,由于本研究重点探究网衣对网箱结构响应计算结果的影响,因此波浪相位角方面,直接采用庞国良等[28]中的结果,具体计算工况见表2。
表 2 网箱结构响应计算工况Table 2. Loading conditions of structural response of cage工况
Loading condition载荷方向
Loading direction/(°)流速
Current velocity/(m·s−1)相位角
Phase angle/(°)工况
Loading condition载荷方向
Loading direction/(°)流速
Current velocity/(m·s−1)相位角
Phase angle/(°)LC1 0 0 75 LC12 45 1.00 90 LC2 0 0.25 75 LC13 45 1.25 90 LC3 0 0.50 75 LC14 45 1.50 90 LC4 0 0.75 75 LC15 90 0 102 LC5 0 1.00 75 LC16 90 0.25 102 LC6 0 1.25 75 LC17 90 0.50 102 LC7 0 1.50 75 LC18 90 0.75 102 LC8 45 0 90 LC19 90 1.00 102 LC9 45 0.25 90 LC20 90 1.25 102 LC10 45 0.50 90 LC21 90 1.50 102 LC11 45 0.75 90 3. 网箱结构响应计算与分析
3.1 有无网衣对网箱结构响应评估的影响
根据表2所列计算工况,开展自升式桁架网箱有无网衣状态下的结构响应计算。对于自升式网箱来说,在外部载荷作用下桩腿是支撑网箱的最关键部件,因此以桩腿支反力作为网箱结构响应评估结果的一个指标。此外,网箱主体桁架管的强度校核作为评估网箱结构安全的关键,其作为网箱结构响应评估结果的另一个指标。在SESAM/GeniE中,梁单元的校核结果可以直接以UF值的形式自动输出,本研究中网箱桁架管被模拟为梁单元,选择通过UF值来反映桁架管的强度校核结果。
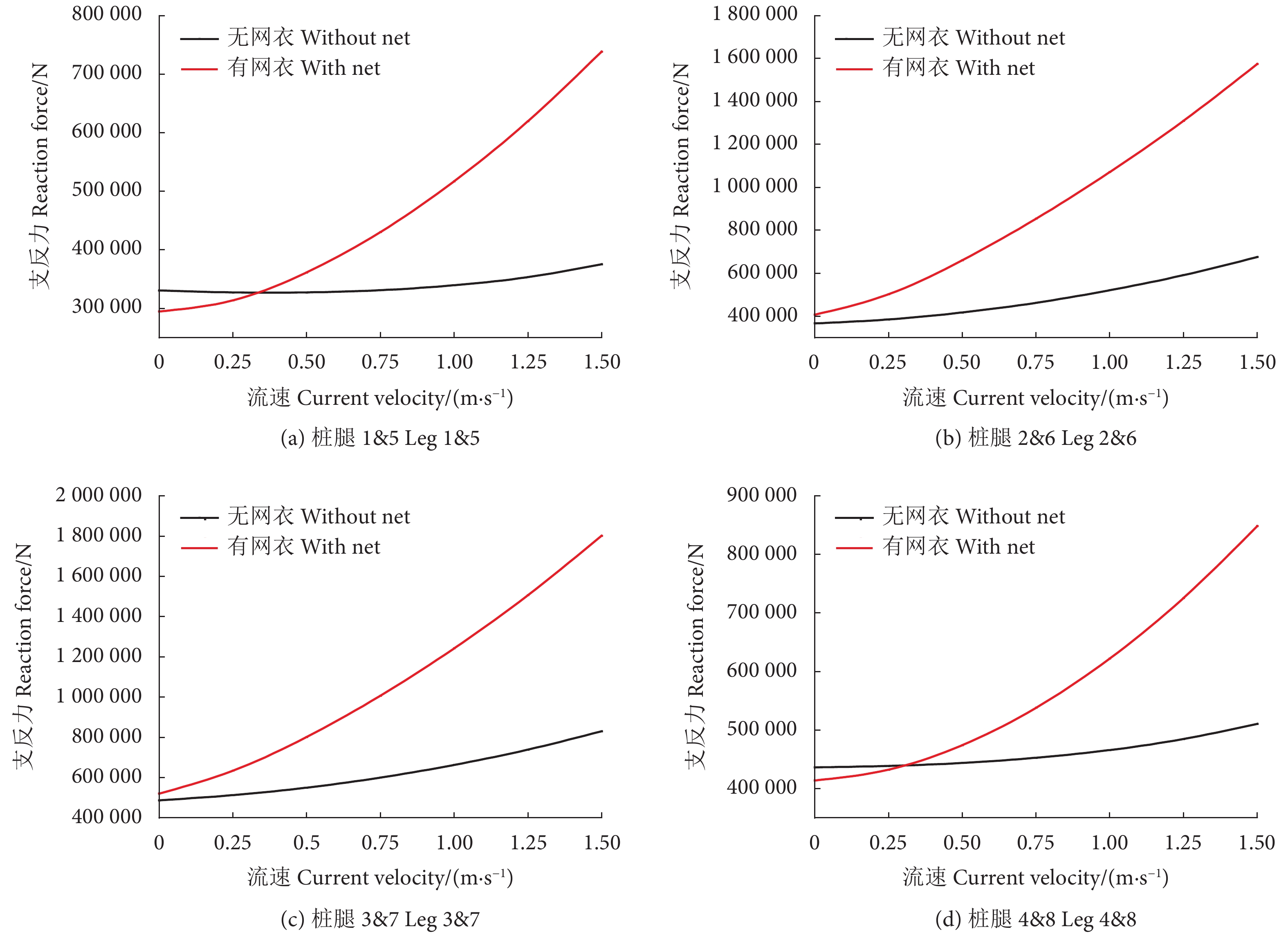
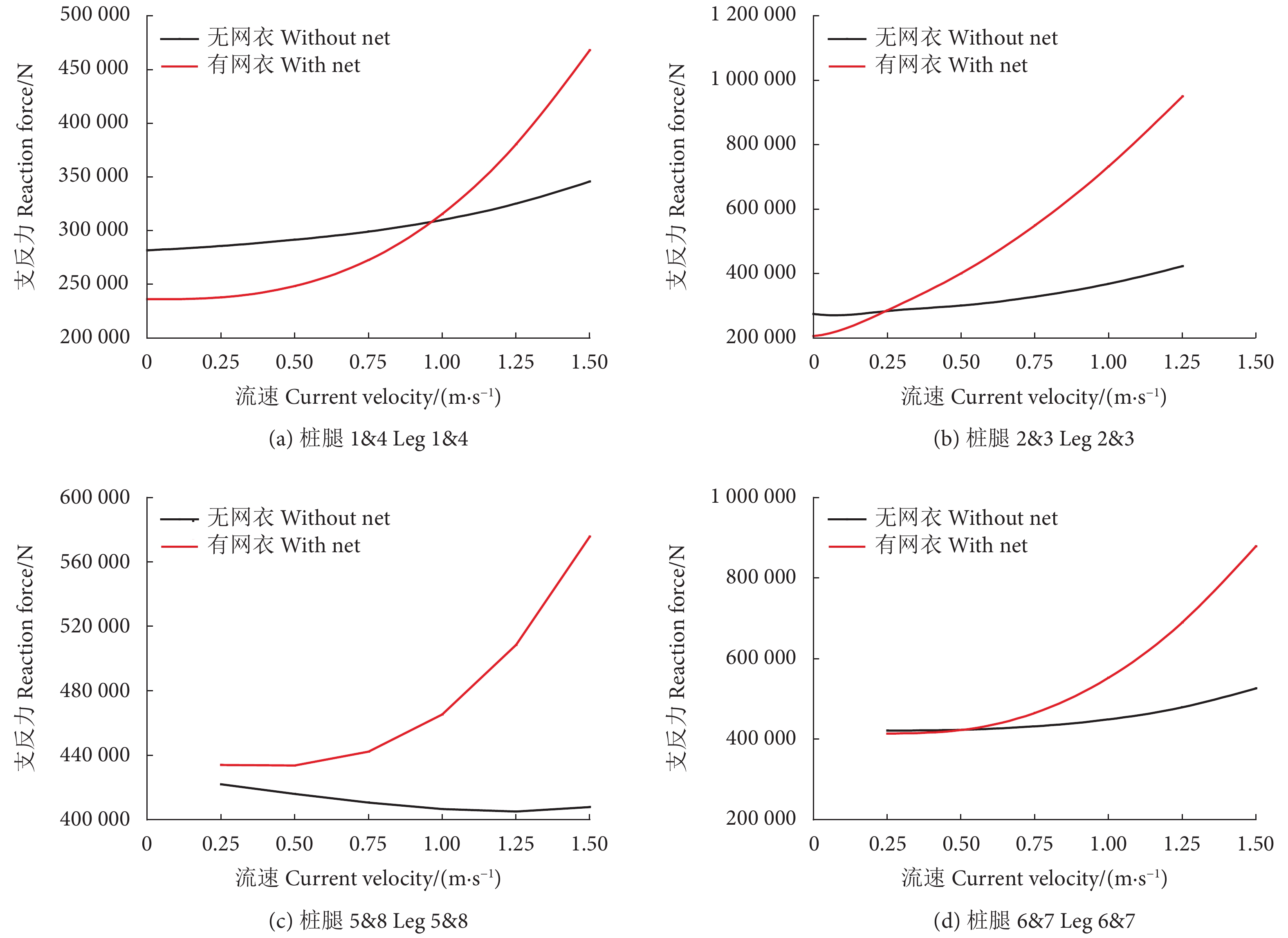
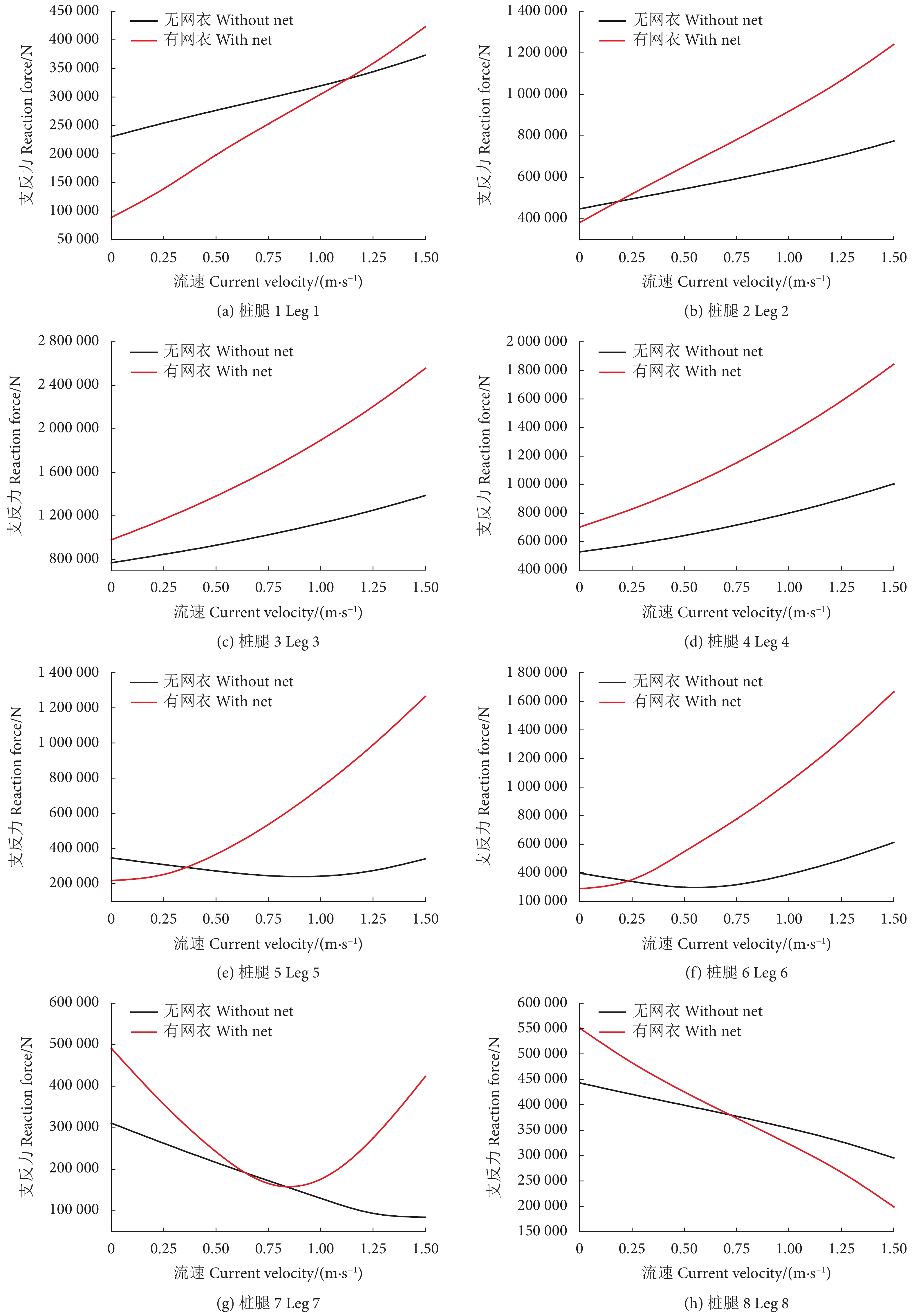
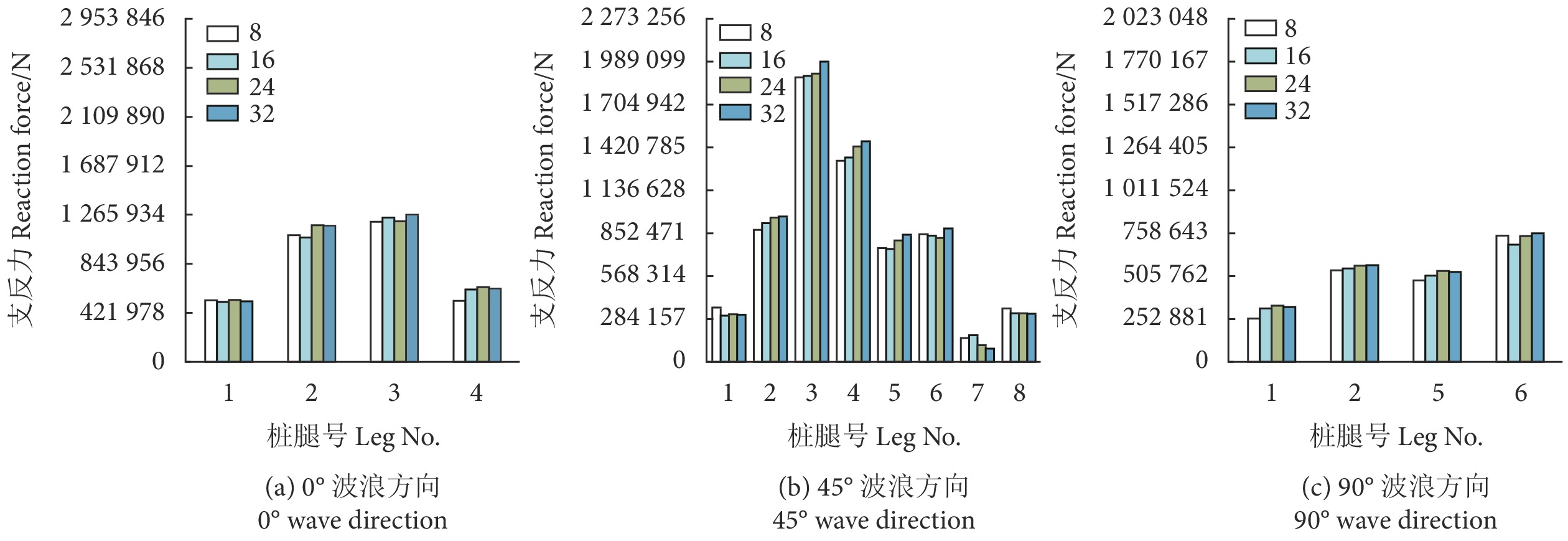
0°、45° 和90° 波流方向条件,网箱有无网衣状态下各个桩腿支反力计算结果见图5—图7。由于网箱结构对称性,0° 波浪方向下,桩腿2&6、1&5、3&7、4&8的支反力基本相同,90° 波浪方向下,桩腿1&4、2&3、5&8、6&7的支反力基本相同。从图中可以看出,除个别工况条件可能受到波浪相位选择的影响,不同波流方向下有网衣网箱的桩腿支反力基本均大于无网衣网箱的,并且随着流速的增加网箱在有无网衣状态下的桩腿支反力差值增大。
表3为不同工况条件网箱有无网衣状态下的主体桁架结构最大UF值。可以看出,考虑网衣后,网箱主体桁架UF值明显增大,比如在工况LC7中,两者相差近26%,并且随着流速的增加UF值不断增大。
表 3 有无网衣的利用系数值Table 3. Utilization factor value with net and without net工况
Loading condition有网衣
With net无网衣
Without net工况
Loading condition有网衣
With net无网衣
Without net工况
Loading condition有网衣
With net无网衣
Without netLC1 0.66 0.59 LC8 0.74 0.67 LC15 0.30 0.26 LC2 0.67 0.59 LC9 0.76 0.67 LC16 0.30 0.28 LC3 0.69 0.60 LC10 0.77 0.68 LC17 0.31 0.30 LC4 0.71 0.61 LC11 0.79 0.69 LC18 0.35 0.30 LC5 0.74 0.61 LC12 0.80 0.70 LC19 0.36 0.31 LC6 0.77 0.62 LC13 0.82 0.71 LC20 0.45 0.35 LC7 0.81 0.64 LC14 0.84 0.72 LC21 0.50 0.43 从网箱桩腿支反力和桁架结构强度的校核结果可以看出,网衣对网箱主体结构的响应评估有较大影响,尤其是在高流速条件下,网衣的存在会大幅增加网箱桩腿受力和桁架UF值,说明在进行桁架网箱结构响应评估时不可忽略网衣的作用。
3.2 不同网衣参数对网箱结构响应评估的影响
3.2.1 不同群化比
在自升式桁架网箱结构响应仿真评估中,上文论证了网衣部件作用不可忽略,但在实际建模过程中,由于网目尺寸较小,网衣建模均需要进行群化处理。为了评估不同网衣群化比对网箱结构响应评估的影响,根据2.2所述,在不改变网线直径的情况下,分别建立了30×30、40×40和50×50网衣群化比下自升式网箱模型。计算工况方面,根据3.1的结果,在0°、45° 和90° 方向分别选取工况LC5、LC12和LC19进行计算,同样通过桩腿支反力和桁架结构强度校核结果开展相关评估。
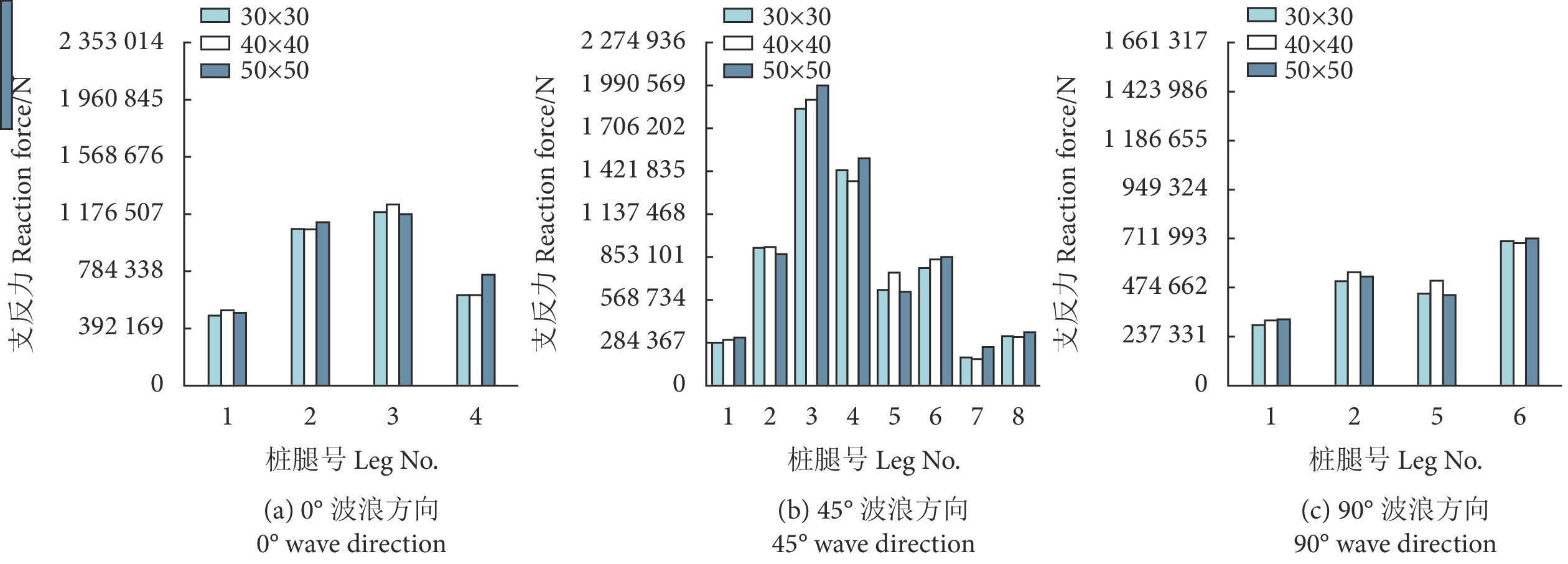
从图8中可以看出,在不同工况、不同网衣群化比条件下,桩腿支反力相差不大,最大差值出现在45° 波浪方向工况LC12,桩腿3支反力在网衣群化比为50×50时为1 990 566.486 N,网衣群化比为30×30时为1 836 066 N,两者相差8.4%。从表4中可以看出,在不同工况、不同网衣群化比条件下,网箱桁架UF值相差也不大,最大差值出现在工况LC5,50×50和40×40群化比间,相差0.07,可见不同群化比的网衣对自升式深海网箱整体结构强度计算的影响不大。此外,在数值仿真中,若重点关注的是桁架结构响应情况,且为获得更高的仿真计算效率,可以选择较大的网目群化比对网箱网衣部分进行群化处理。
![]() 图 8 不同群化比下不同波浪方向桩腿支反力Figure 8. Leg reaction force in different wave directions with different mesh grouping ratios表 4 不同群化比下的的利用系数值Table 4. Utilization factor values with different mesh grouping ratios
图 8 不同群化比下不同波浪方向桩腿支反力Figure 8. Leg reaction force in different wave directions with different mesh grouping ratios表 4 不同群化比下的的利用系数值Table 4. Utilization factor values with different mesh grouping ratios群化比
Grouping ratioLC5 LC12 LC19 30×30 0.75 0.77 0.30 40×40 0.74 0.80 0.36 50×50 0.81 0.83 0.35 3.2.2 网衣系缚方式
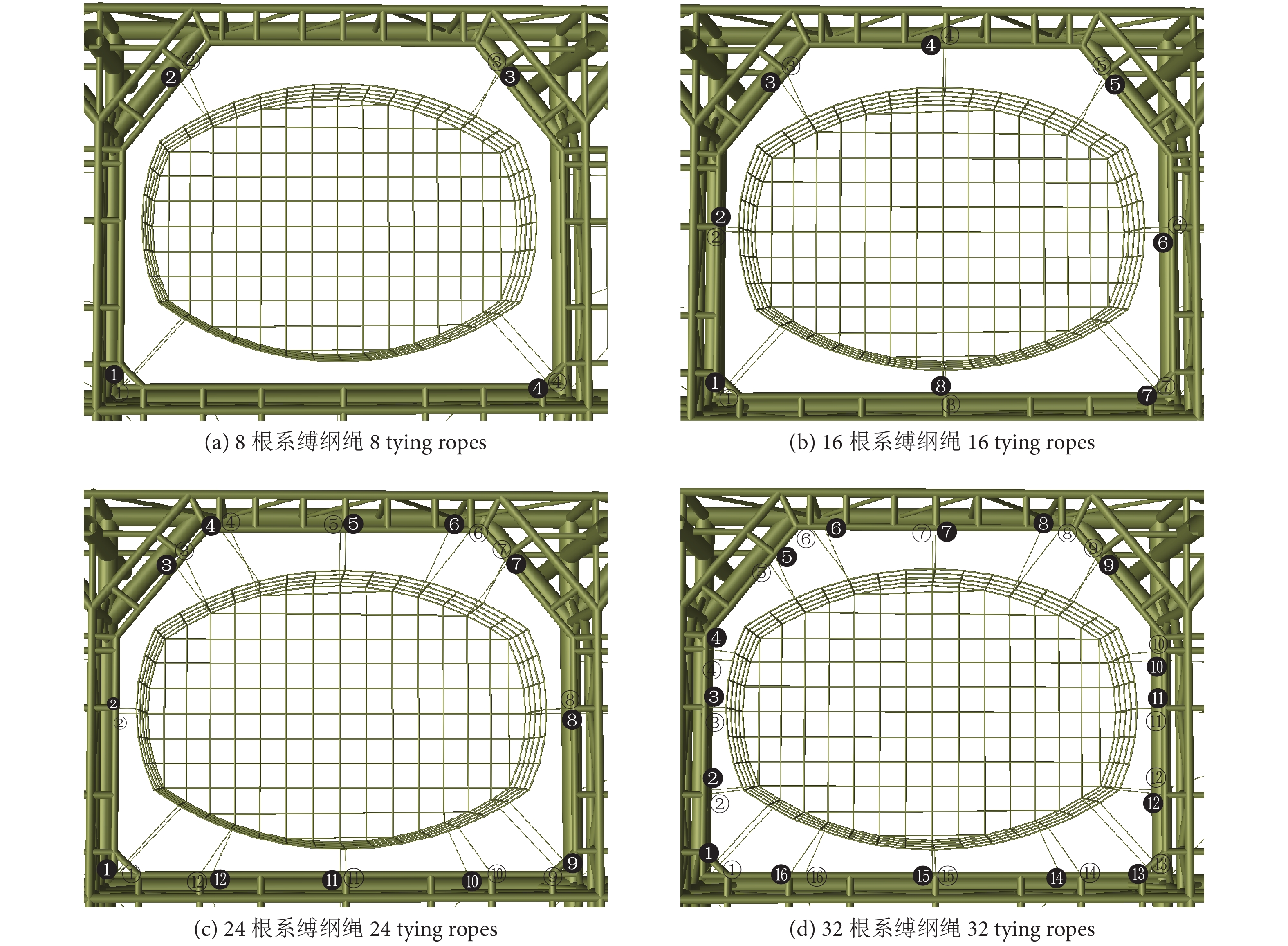
作为网衣与网箱主体框架的连接部分,网衣在布设时不同数量的系缚纲绳可能会对网箱整体结构响应有影响,通过设置8、16、24和32根系缚纲绳,同样在0°、45° 和90° 方向分别选取工况LC5、LC12和LC19开展相关计算,系缚纲绳及系缚点位置如图9所示。
![]() 图 9 系缚纲绳及系缚点位置注:空心圆代表上系缚点的编号,实心圆代表下系缚点的编号。Figure 9. Position of tying rope and tying pointNote: The hollow circles represent upper tie points No. and the solid circles represents lower tie points No..
图 9 系缚纲绳及系缚点位置注:空心圆代表上系缚点的编号,实心圆代表下系缚点的编号。Figure 9. Position of tying rope and tying pointNote: The hollow circles represent upper tie points No. and the solid circles represents lower tie points No..不同波浪方向下不同数量系缚纲绳的网箱对应的桩腿支反力如图10所示。可以看出,在45°波浪方向下,8根与32根系缚纲绳网箱的5号桩腿支反力差值最大 (9.8%),其余桩腿支反力的变化不大。对比表5中不同系缚纲绳数量下的UF值,不难看出最大差值出现在LC12的8根系缚纲绳的网箱与24、32根系缚纲绳的网箱上,但仅相差0.05,因此,网衣系缚纲绳数量对桩腿支反力和网箱桁架结构强度校核结果影响不大。
![]() 图 10 不同波浪方向下不同系缚纲绳数量的框架所对应的桩腿支反力Figure 10. Leg reaction force with different number of tying ropes in different wave directions表 5 不同系缚绳数量下的利用系数值Table 5. Utilization factor values with different number of tying ropes
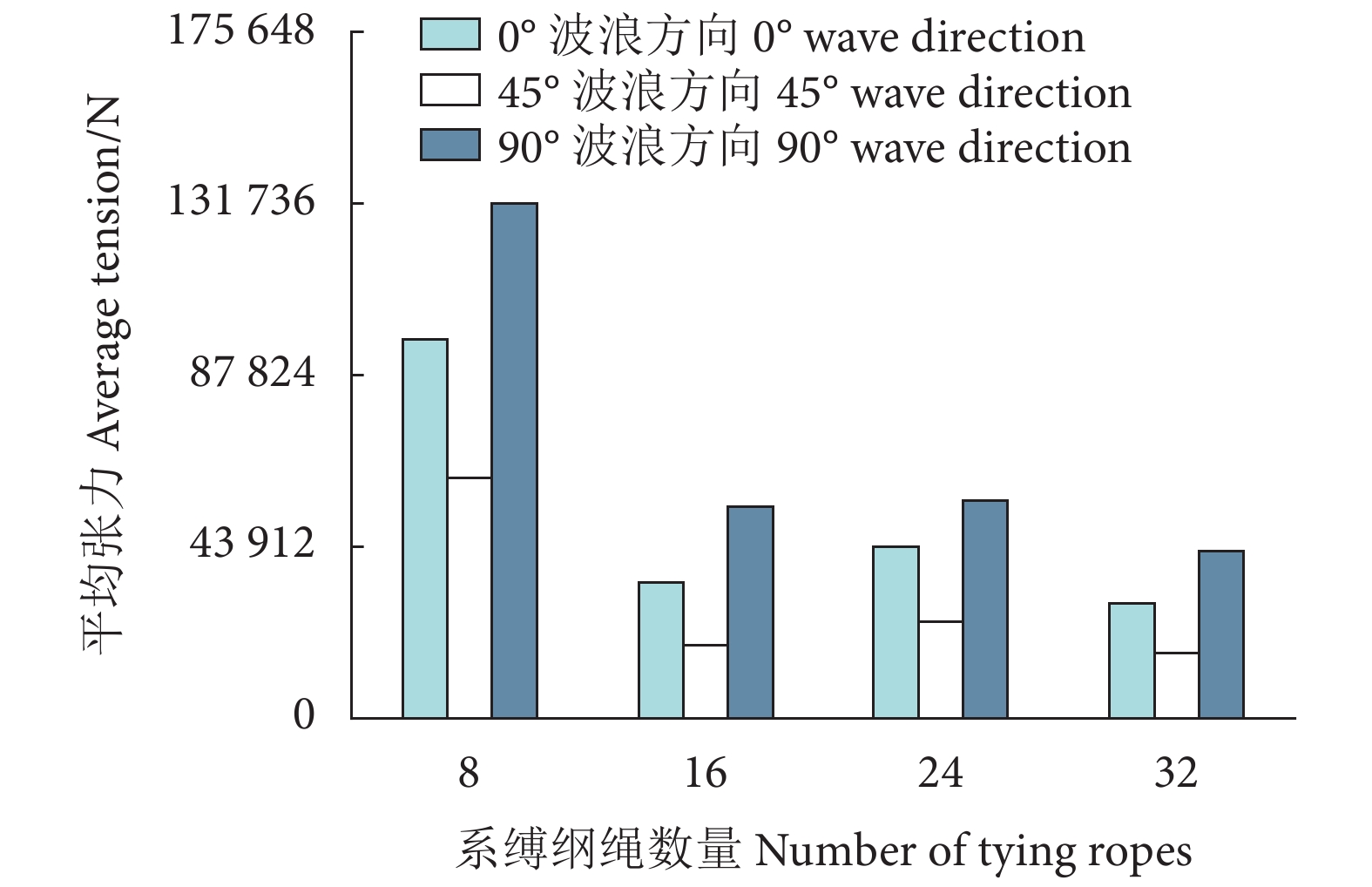
图 10 不同波浪方向下不同系缚纲绳数量的框架所对应的桩腿支反力Figure 10. Leg reaction force with different number of tying ropes in different wave directions表 5 不同系缚绳数量下的利用系数值Table 5. Utilization factor values with different number of tying ropes系缚纲绳数量 Number of tying rope LC5 LC12 LC19 8 0.75 0.77 0.32 16 0.74 0.80 0.36 24 0.75 0.82 0.34 32 0.75 0.82 0.33 图11为不同数量系缚纲绳下网箱系缚纲绳在3个波浪方向下的平均张力,可以看出3个波浪方向下,随着系缚纲绳的增加,系缚的平均张力明显减少。因此,为保证网衣系缚安全,在网衣制作中增加系缚纲绳是易于操作的有效手段。
4. 结论
本研究针对自升式桁架网箱,着重探究了网衣对网箱结构响应评估的影响,通过构建网箱仿真计算模型,分析了在有无网衣及不同网衣参数条件下的网箱主体结构响应。结果表明:1) 网衣对网箱主体结构强度存在较大影响,尤其在高流速条件下,网衣的存在会大幅增加网箱桩腿及主体结构的受力,证明在校核网箱结构强度时不可忽略网衣的作用;2) 通过对比30×30、40×40、50×50这3组不同网目群化比下的桩腿支反力和UF值,发现不同群化比下的网衣对自升式桁架网箱整体结构强度和网箱桩腿受力的影响变化不大,在数值仿真中可以考虑使用较大的群化比来提升计算效率;3) 通过对8、16、24和32根系缚纲绳下的网箱桩腿支反力、网箱主体UF值以及系缚纲绳的平均张力对比分析,可以看出网衣系缚纲绳数量对网箱整体结构强度评估和网箱桩腿受力基本无影响,但增加系缚绳可以有效减少系缚点受力。由于本研究是基于SESAM/Genie梁单元模拟网衣网线,未考虑网衣柔性变形特性以及网衣系缚后的缩结效应等,后续将进一步考虑实际情况,深化相关研究。
-
![]()
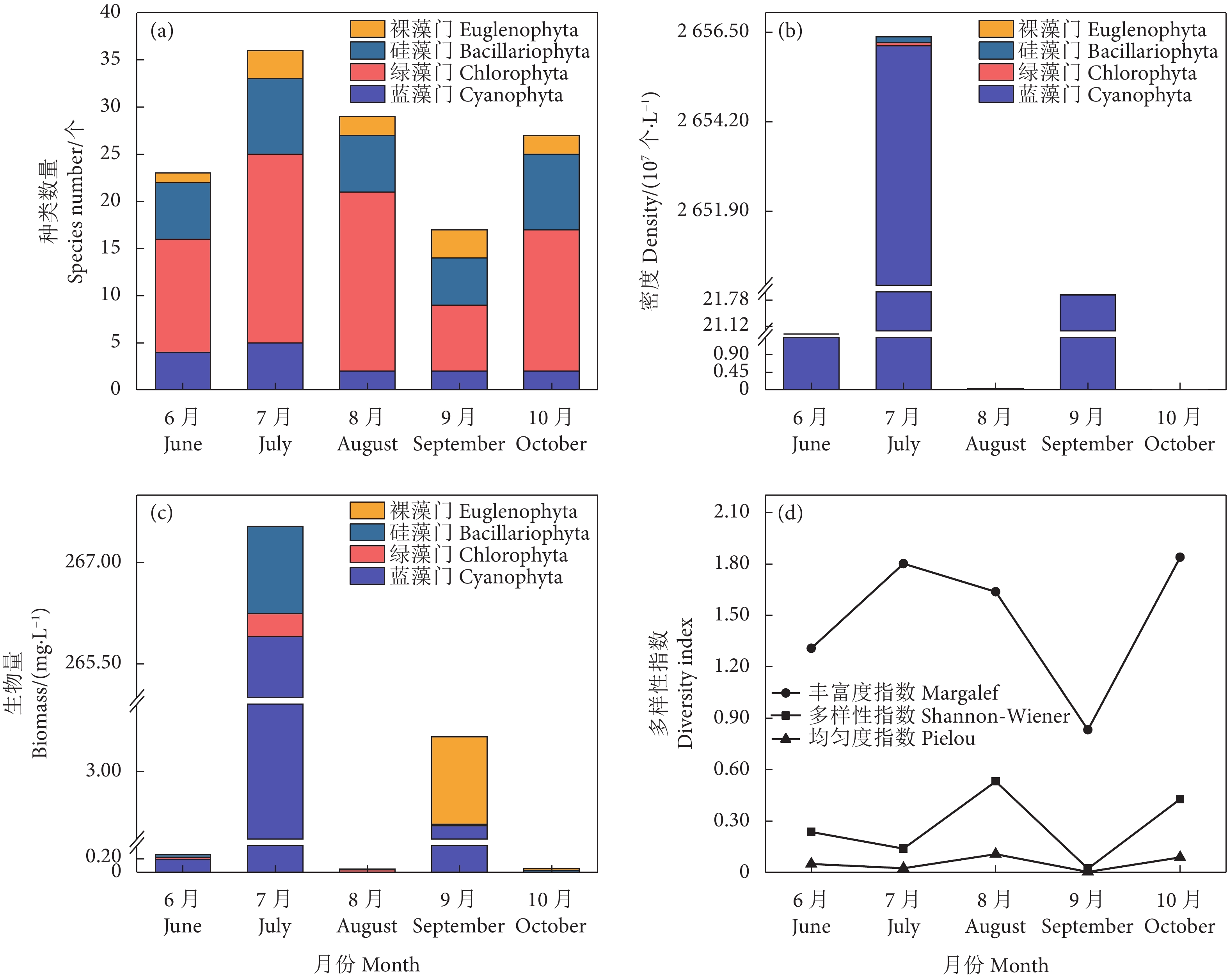
图 1 不同月份的浮游植物种类数量、密度、生物量和多样性指数
Figure 1. Species composition, density, biomass and diversity index of plankton in different months
![]()
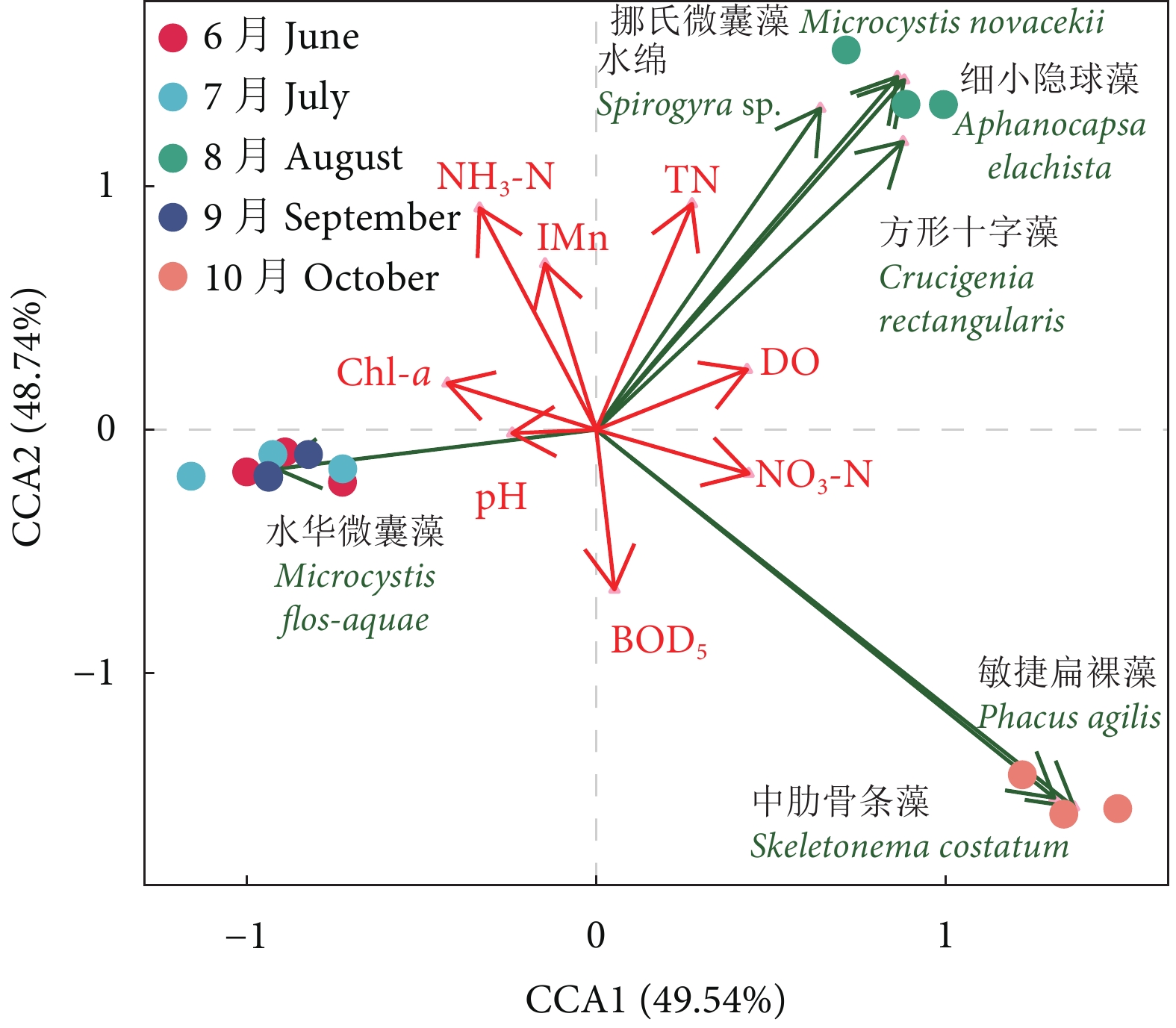
图 2 养殖蟹塘浮游植物优势种与环境因子的CCA排序分析
Figure 2. CCA ordination of phytoplankton dominant species and environmental factors in aquaculture pond of E. sinensis
![]()
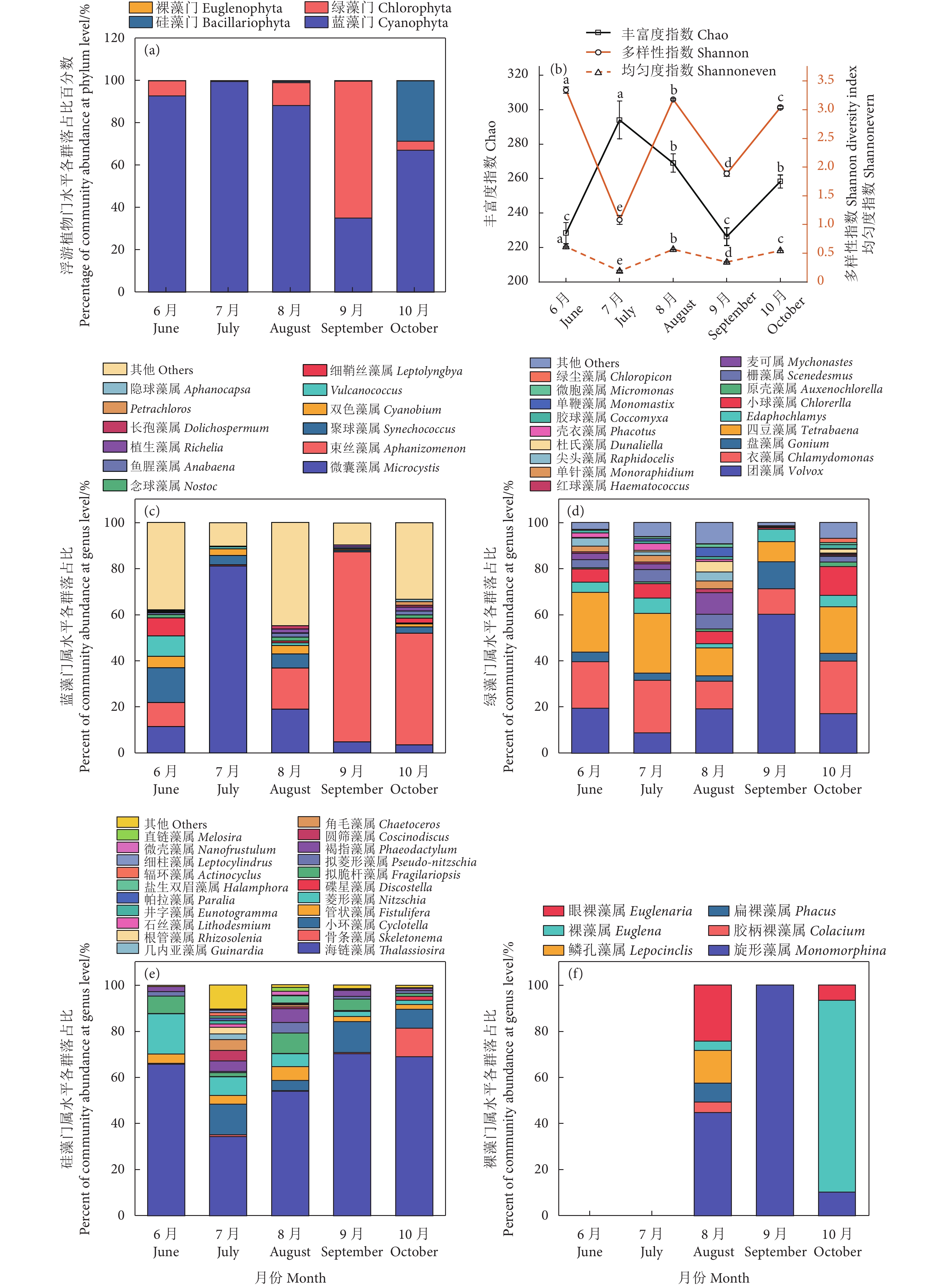
图 3 浮游植物门水平各群落组成(a)、多样性指数(b)、蓝藻门、绿藻门、硅藻门和裸藻门属水平各群落组成(c—f)
Figure 3. Percent ages of community abundance at phylum level (a) and diversity indices of plankton (b), percent of community abundance at genus level of Cyanophyta, Chlorophyta, Bacillariophyta, and Euglenophyta (c–f) in different months
![]()
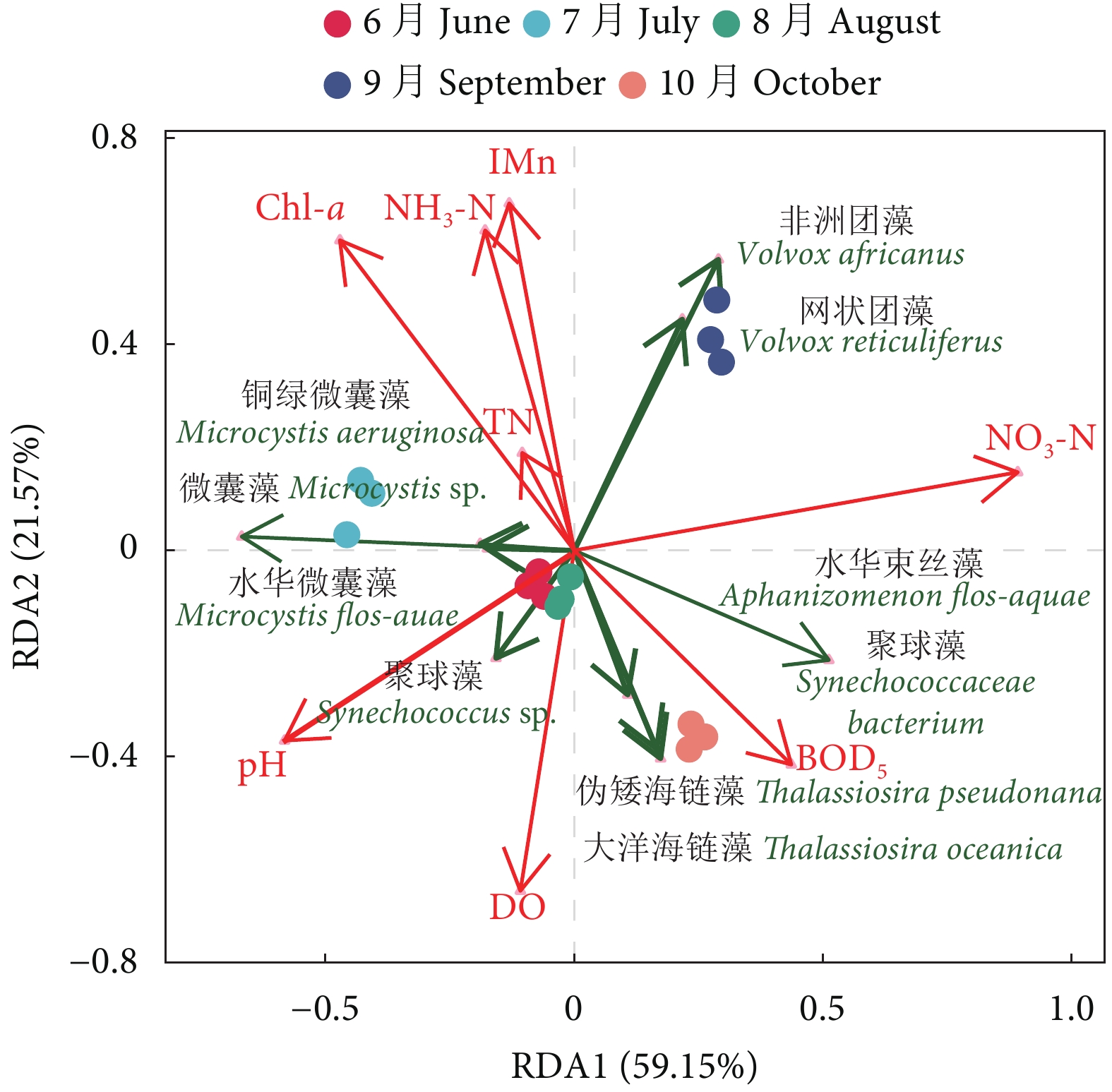
图 4 养殖蟹塘浮游植物优势种与环境因子的RDA排序分析
Figure 4. RDA ordination of phytoplankton dominant species and environmental factors in aquaculture pond of E. sinensis
表 1 中华绒螯蟹养殖池塘水体理化指标
Table 1 Physicochemical parameters in water bodies of E. sinensis aquaculture ponds
指标
Index6月
June7月
July8月
August9月
September10月
OctoberpH 9.97±0.06a 9.15±0.25b 8.65±0.44b 7.92±0.39c 8.62±0.13b 水温 Water temperature/℃ 33.75±0.43a 33.78±1.27a 29.63±0.04b 22.00±0.02c 20.04±0.54d 溶解氧 DO/(mg·L−1) 11.88±1.61a 6.61±1.00bc 11.07±1.45a 5.23±0.95c 9.37±2.62ab 叶绿素 a Chl-a/(μg·L−1) 1.33±0.58d 50.67±2.08a 20.67±8.08c 33.33±6.11b 7.00±1.00d 总磷 TP/(mg·L−1) 0.11±0.01d 0.23±0.02c 0.34±0.01b 0.43±0.04a 0.09±0.01d 磷酸盐 PO4/(mg·L−1) 0.08±0.01c 0.16±0.03b 0.30±0.02a 0.28±0.02a 0.03±0.00d 总氮 TN/(mg·L−1) 0.89±0.03c 1.14±0.09b 2.41±0.24a 1.25±0.10b 0.78±0.02c 氨氮 NH3-N/(mg·L−1) 0.65±0.01c 0.55±0.04d 0.88±0.07a 0.75±0.04b 0.21±0.01e 亚硝态氮 NO2-N/(mg·L−1) 0.01±0.00b 0.01±0.00b 0.05±0.00a 0.01±0.00b 0.01±0.00b 硝态氮 NO3-N/(mg·L−1) 0.02±0.01d 0.01±0.01d 0.23±0.02c 0.46±0.04a 0.37±0.04b 高锰酸钾指数 IMn/(mg·L−1) 5.59±0.07b 7.43±0.20a 8.04±0.39a 8.06±1.20a 5.53±0.15b 生物需氧量 BOD5/(mg·L−1) 5.20±0.00a 3.50±0.10c 3.57±0.06c 4.07±0.25b 5.00±0.53a 注:同行数据不同上标字母表示组间存在显著性差异 (P<0.05);n=3。 Note: Values with different superscripts within each row are significantly different (P<0.05); n=3.  下载: 导出CSV
下载: 导出CSV
表 2 中华绒螯蟹养殖池塘浮游植物优势种优势度
Table 2 Dominance index of phytoplankton dominant species in aquaculture pond of E. sinensis
浮游植物
Phytoplankton优势种
Dominant species6月
June7月
July8月
August9月
September10月
October蓝藻门 Cyanophyta 水华微囊藻 Microcystis flos-aquae 0.98 1.00 — 1.00 — 细小隐球藻 Aphanocapsa elachista — — 0.82 — — 挪氏微囊藻 Microcystis novacekii — — 0.04 — — 绿藻门 Chlorophyta 水绵 Spirogyra sp. — — 0.04 — — 方形十字藻 Crucigenia rectangularis — — 0.04 — — 硅藻门 Bacillariophyta 中肋骨条藻 Skeletonema costatum — — — — 0.88 裸藻门 Euglenophyta 敏捷扁裸藻 Phacus agilis — — — — 0.06 注:数字表示物种优势度指数;“—”表示物种未达到优势度0.02。 Note: The numbers represent dominance index of the species; "—" represents the species has not reached the dominance standard of 0.02.
下载: 导出CSV
表 3 中华绒螯蟹养殖池塘浮游植物优势种占比
Table 3 Composition of phytoplankton dominant species in aquaculture pond of E. sinensis
浮游植物
Phytoplankton优势种
Dominant species占比前三的种
Top three species/%6月
June7月
July8月
August9月
September10月
October蓝藻门 Cyanophyta 水华束丝藻 Aphanizomenon flos-aquae 9.63 15.73 28.86 32.52 聚球藻 Synechococcus sp. 8.06 铜绿微囊藻 Microcystis aeruginosa 6.79 52.26 10.52 微囊藻 Microcystis sp. 4.00 水华微囊藻 Microcystis flos-aquae 3.70 聚球藻 Synechococcaceae bacterium 7.63 绿藻门 Chlorophyta 非洲团藻 Volvox africanus 15.84 网状团藻 Volvox reticuliferus 12.79 硅藻门 Bacillariophyta 大洋海链藻 Thalassiosira oceanica 9.82 伪矮海链藻 Thalassiosira pseudonana 9.75
下载: 导出CSV
-
[1] 周永昌, 姜晓东, 龙晓文, 等. 中草药对中华绒螯蟹幼蟹生长及免疫力的影响[J]. 水产科学, 2019, 38(2): 188-193. [2] KATMOKO G M D, RISJANI Y, MASITHAH E D. Analysis of phytoplankton structure community, water quality and cultivation performance in Litopenaeus vannamei intensive pond located in Tembokrejo Village, Muncar, Banyuwangi[J]. J Exp Life Sci, 2021, 11(3): 68-76. doi: 10.21776/ub.jels.2021.011.03.01
[3] 孙军, 宋煜尧, 刘海娇. 海洋浮游植物快速鉴定与监测技术[J]. 海洋科学进展, 2022, 40(4): 701-716. [4] 侯德昌, 张莹莹, 魏文志. 不同中华鳖养殖模式浮游植物功能群特征及水环境评价[J]. 安徽农业科学, 2022, 50(9): 96-99, 170. doi: 10.3969/j.issn.0517-6611.2022.09.023 [5] MERCURIO A L, QUERIJERO B, CHING J. Phytoplankton community in aquaculture and non-aquaculture sites of Taal Lake, Batangas, Philippines[J]. J Exp Biol Agric Sci, 2016, 4(1): 66-73. doi: 10.18006/2016.4(1).66.73
[6] 蒲炜佳, 董世鹏, 张东旭, 等. 三疣梭子蟹池塘综合养殖系统浮游植物群落结构及其与环境因子的关系[J]. 中国水产科学, 2022, 29(4): 549-561. [7] MANOYLOV K M. Taxonomic identification of algae (morphological and molecular): species concepts, methodologies, and their implications for ecological bioassessment[J]. J Phycol, 2014, 50(3): 409-424. doi: 10.1111/jpy.12183
[8] 方圆, 黎慧, 王李宝, 等. 凡纳滨对虾小型温棚养殖模式水体微生物群落的研究[J]. 南方水产科学, 2023, 19(3): 29-41. [9] 陈琼, 李贵阳, 罗坤, 等. 凡纳滨对虾 (Litopenaeus vannamei) 亲虾繁殖期水体微生物多样性[J]. 海洋与湖沼, 2017, 48(1): 130-138. [10] 高权新, 李云莉, 齐占会, 等. 基于宏基因组学的中国沿海密集养殖水域秋季底质细菌多样性研究[J]. 海洋渔业, 2019, 41(3): 354-363. [11] 卫鹏, 毕相东, 戴伟, 等. 淡水养殖池塘微型和超微型浮游植物的群落结构组成[J]. 大连海洋大学学报, 2022, 37(1): 113-119. [12] 乔玲, 常志强, 李健, 等. 基于形态学和高通量测序的海水池塘生态养殖系统中浮游植物多样性比较[J]. 渔业科学进展, 2022, 43(2): 32-43. [13] 国家环境保护总局水和废水监测分析方法编委会. 水和废水监测分析方法[M]. 北京: 中国环境出版社集团, 2013: 88-284. [14] 赵文. 水生生物学[M]. 北京: 中国农业出版社, 2015: 16-126, 493-498. [15] FU R B, LI Z P, ZHOU R, et al. The mechanism of intestinal flora dysregulation mediated by intestinal bacterial biofilm to induce constipation [J]. Bioengineered, 2021, 12(1): 6484-6498.
[16] STALDER T, BARRAUD O, JOVÉ T, et al. Quantitative and qualitative impact of hospital effluent on dissemination of the integron pool[J]. ISME J, 2014, 8(4): 768-777. doi: 10.1038/ismej.2013.189
[17] LIU J F, LAI L Y, LIN J J, et al. Ranitidine and finasteride inhibit the synthesis and release of trimethylamine N-oxide and mitigates its cardiovascular and renal damage through modulating gut microbiota[J]. Int J Biol Sci, 2020, 16(5): 790-802. doi: 10.7150/ijbs.40934
[18] 刘淑娟. 中肋骨条藻 (Skeletonema costatum) 赤潮生消规律及其对浮游生物群落结构的影响[D]. 舟山: 浙江海洋大学, 2019: 3-6. [19] 赵艳民, 秦延文, 张雷, 等. 基于GIS的近30年长江口及其邻近海域赤潮时空分布特征研究[J]. 海洋科学, 2021, 45(12): 39-46. [20] 胡婧文, 弋钰昕, 尤庆敏, 等. 骨条藻属硅藻的分类、生态及生理研究进展[J]. 上海师范大学学报(自然科学版), 2020, 49(6): 698-708. [21] 韩志男. 崇明岛海水入侵特征及趋势分析[D]. 青岛: 国家海洋局第一海洋研究所, 2013: 1-6. [22] 董立新, 周绪申. 浮游植物多样性指数在内陆水体污染类型评价中的应用简述[J]. 海河水利, 2017(5): 57-60. [23] 龚进玲, 李学梅, 何勇凤, 等. 青鱼—鲢、鳙生态池塘浮游生物群落结构特征及其与环境因子的关系[J]. 水产科学, 2022, 41(4): 517-526. [24] 孟得微, 孙博, 张立民, 等. 东北盐碱池塘浮游植物群落结构特征与环境因子的关系[J]. 安徽农业科学, 2021, 49(21): 110-115. [25] 温旭, 马旭洲, 范伟, 等. 不同面积芦苇稻幼蟹塘浮游植物功能类群的结构特征[J]. 浙江大学学报(农业与生命科学版), 2019, 45(1): 85-94. [26] FAN L M, HU G D, QIU L P, et al. Variations in bacterioplankton communities in aquaculture ponds and the influencing factors during the peak period of culture[J]. Environ Pollut, 2020, 258: 113656. doi: 10.1016/j.envpol.2019.113656
[27] 夏德军, 隋燚, 王云平, 等. 稻田推水增氧养殖小龙虾新技术[J]. 中国水产, 2019(1): 88-90. [28] NONG X Z, SHAO D G, SHANG Y M, et al. Analysis of spatio-temporal variation in phytoplankton and its relationship with water quality parameters in the South-to-North Water Diversion Project of China[J]. Environ Monit Assess, 2021, 193(9): 593. doi: 10.1007/s10661-021-09391-6
[29] JIANG Y J, HE W, LIU W X, et al. The seasonal and spatial variations of phytoplankton community and their correlation with environmental factors in a large eutrophic Chinese lake (Lake Chaohu)[J]. Ecol Indic, 2014, 40: 58-67. doi: 10.1016/j.ecolind.2014.01.006
[30] 刘乾甫, 赖子尼, 高原, 等. 珠江三角洲地区精养淡水鱼塘浮游植物功能群特征[J]. 中国水产科学, 2018, 25(1): 124-136. [31] SIPAOBA-TAVARES L H, DIAS S G. Water quality and communities associated with macrophytes in a shallow water-supply reservoir on an aquaculture farm[J]. Braz J Biol, 2014, 74(2): 420-428. doi: 10.1590/1519-6984.27212
[32] 俞焰, 刘德富, 杨正健, 等. 千岛湖溶解氧与浮游植物垂向分层特征及其影响因素[J]. 环境科学, 2017, 38(4): 1393-1402. [33] 田盼, 宋林旭, 纪道斌, 等. 三峡库区神农溪不同时期溶解氧与叶绿素a垂向分布特征[J]. 水生态学杂志, 2022, 43(2): 1-8. [34] TAN X, GU H H, RUAN Y N, et al. Effects of nitrogen on interspecific competition between two cell-size cyanobacteria: Microcystis aeruginosa and Synechococcus sp.[J]. Harmful Algae, 2019, 89: 101661. doi: 10.1016/j.hal.2019.101661
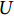
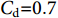
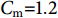
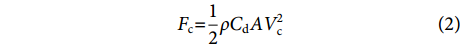
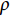
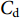
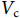
计量
- 文章访问数: 123
- HTML全文浏览量: 70
- PDF下载量: 34
 粤公网安备 44010502001741号
粤公网安备 44010502001741号
